II. METABOLISMO BACTERIANO
TODOS los seres vivos llevan a cabo el procesamiento de los nutrientes que los mantienen vivos. A este conjunto de procesos, se le conoce como metabolismo y consiste de un gran número de reacciones químicas destinadas a transformar las moléculas nutritivas en elementos que posteriormente serán utilizados para la síntesis de los componentes estructurales; como pueden ser las proteínas. Otra parte importante del metabolismo es la de transformar y conservar la energía que está contenida en una reacción química en algún proceso que requiera de energía, como puede ser el trabajo o el movimiento.
Es evidente que los nutrientes son transformados cuando entran en un organismo, ya que en ningún caso el alimento contiene todas las moléculas que una célula requiere. Esto se vio con claridad al observar el crecimiento normal de levaduras en un medio de cultivo que sólo contenía glucosa como única fuente de energía. Así pues, se pensó que la síntesis de todos los componentes celulares se llevaba a cabo en el interior de las levaduras. Hoy sabemos que las transformaciones que sufre la glucosa no ocurren en un solo paso, sino que, por el contrario, se forman varios productos intermedios que en muchas ocasiones no tienen una función específica a no ser la de formar parte de lo que se conoce como vía metabólica.
La transformación de los nutrientes en compuestos útiles para la subsistencia de un organismo se lleva a cabo por medio de las reacciones químicas que realizan unas proteínas conocidas como enzimas. De tal forma que no podemos hablar del metabolismo si no describimos brevemente qué son y cómo funcionan las enzimas.
II.1 LAS ENZIMAS SON EFICIENCIA Y RAPIDEZ
La enzimología es una parte fundamental de la bioquímica que se ha desarrollado rápidamente en los últimos cinco decenios. Ciertas manifestaciones de la acción enzimática eran conocidas desde mucho tiempo atrás y si bien sus causas eran ignoradas, ya se conocían varios procesos como la fermentación, la digestión y la respiración o consumo de oxígeno.
Las enzimas son proteínas especializadas capaces de transformar químicamente una molécula; son sin duda las moléculas biológicas más notables ya que solamente transforman a una molécula y a ninguna otra y esto lo hacen varios cientos de veces por segundo. Estas características las hacen mucho mejores que cualquier catalizador químico hecho por el hombre.
Figura 8. (a) Cadena lineal de aminoáidos. (b) Cadena tridimensional de aminoácidos.
Pasteur pensaba que la fermentación era llevada a cabo por enzimas y que su acción estaba íntimamente ligada a la estructura y a la vida de las levaduras. Un hecho de gran importancia que marcó el inicio de la enzimología y de la bioquímica ocurrió cuando, en 1877, Edward Buchner logró que un extracto libre de levaduras enteras llevara a cabo la fermentación alcohólica. Esto implicaba que tales enzimas, de gran importancia para la célula viva, eran capaces de llevar a cabo su función con independencia de la integridad de la estructura celular. En 1926, el notable bioquímico J. B. Sumner aisló y cristalizó por primera vez una enzima, la ureasa de frijol, y comprobó que era una proteína, lo que era contrario a la opinión general en aquel tiempo. Tuvieron que pasar 10 años, hasta 1926 que fue cuando Northrop cristalizó otras tres enzimas, para establecer finalmente que éstas son proteínas especializadas en una reacción. Hoy en día se conocen quizá más de 2 000 enzimas diferentes y, aunque se sabe mucho acerca de ellas, quedan aún grandes dudas sobre su control genético y su modo de acción.
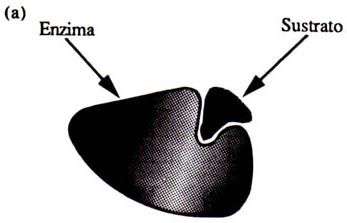 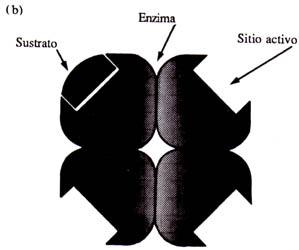 |
Figura 9. (a) Enzima con uno o dos sitios activos y una cadena de aminoácidos. (b) Enzima con varias cadenas de aminoácidos y varios sitios activos.
Estas proteínas actúan sobre una sola molécula, es decir, son como una llave y una cerradura: la llave es la molécula y la cerradura es la enzima. La posibilidad de que una llave abra otra cerradura que no sea la que le corresponde es muy remota. Pues bien, lo mismo ocurre con las enzimas: la posibilidad de que una enzima actué sobre otra molécula que no sea la que le corresponde es muy baja.
Las enzimas, en su calidad de proteínas, se hallan formadas por cadenas de aminoácidos, las cuales se arreglan espacialmente en formas variadas. En otras palabras, las cadenas de aminoácidos constituyen estructuras tridimensionales (Figura 8).
Así, estas complicadas estructuras forman un espacio o hueco en donde se une la molécula que va a ser transformada; a este hueco se le denomina sitio activo. Las enzimas tienen entonces un sitio activo y una estructura tridimensional que las hace ser únicas. Sin embargo, pueden existir enzimas con dos o más sitios activos y con una estructura integrada por una o varias cadenas de aminoácidos (Figura 9).
La velocidad a la que dichas enzimas trabajan es muy alta y esto las hace muy eficientes. Su actividad está en función directa de la cantidad de moléculas que va a ser transformada y su acción es regulada en parte por la aparición en el medio del producto final de la reacción, que de hecho inhibe su actividad. Su acción puede ser concertada es decir, que una enzima se encuentra dentro de un proceso más o menos largo de degradación o de síntesis y de esta forma el producto de dicha enzima será el sustrato de la siguiente enzima y así sucesivamente (Figura 13).
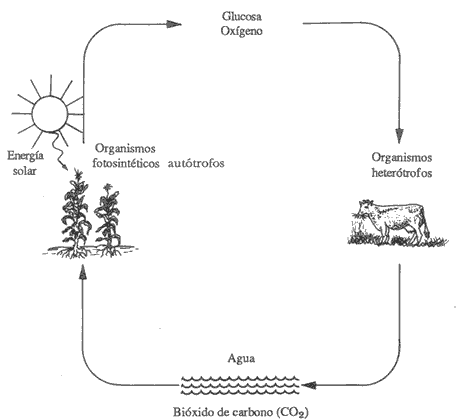
Figura 10. Simplificación de la relación que guardan los organismos autótrofos y heterótrofos.
El metabolismo se podría definir como el conjunto de reacciones químicas que ocurren en la célula y que tienen por objeto generar la energía y los componentes necesarios para que ésta lleve a cabo sus funciones. Esta definición estaría incompleta si no aclaramos antes que todas estas reacciones están altamente coordinadas y dependen del perfecto funcionamiento de los sistemas multienzimáticos.
II.2 UNOS SERES VIVOS DEPENDEN DE OTROS
Organismos autosuficientes y organismos dependientes. Para poder llevar a cabo todas sus reacciones metabólicas, los seres vivos necesitan una fuente de alimento y energía.
De acuerdo con su fuente de obtención de nutrientes, los organismos vivos se pueden dividir en dos grandes grupos. Por una parte, los autosuficientes (autótrofos), que utilizan al bióxido de carbono (CO2) y al agua (H20) como única fuente de alimento y a partir de estos compuestos producen todas las moléculas necesarias para su subsistencia, crecimiento y proliferación. Por otra parte, los que dependen de otros compuestos o de sus productos, se llaman heterótrofos. Estos no pueden utilizar el bióxido de carbono como tal y por lo tanto deben obtener el carbono que necesitan a partir de otros compuestos que se encuentran en el medio que los rodea, como por ejemplo la glucosa, que es una molécula mucho más compleja que el C02. Lo cual quiere decir que unas formas de vida requieren de otras y así se establece una cadena alimenticia. Ya que unos organismos son relativamente autosuficientes y otros requieren de fuentes de carbono más complejas, su interrelación es necesaria y vital. Algunos ejemplos de células autosuficientes son las de las plantas, las bacterias fotosintéticas y algunas bacterias no fotosintéticas. Sin embargo, la mayoría de los organismos no son autosuficientes y por lo tanto requieren siempre de una asociación benéfica con un organismo que sí lo sea.
El ciclo del carbono y del oxígeno. En la figura 10 podemos ver cómo se relacionan los organismos fotosintéticos y los que dependen de ellos. Los primeros, por medio de la energía luminosa del Sol, más el agua y el bióxido de carbono (C02) del suelo, llevan a cabo la fotosíntesis, produciendo así glucosa y oxígeno. A partir de estos compuestos básicos que se convierten en materia orgánica, los organismos dependientes obtienen a su vez los nutrientes que, al ser utilizados, son transformados y liberados al medio en forma de agua y C02. Estas moléculas a su vez serán nuevamente utilizadas por los organismos fotosínteticos.
Así pues, la interrelación entre ambos tipos de organismos crea a su vez un ciclo importante para la conservación de la vida en la Tierra. A éste se le conoce como ciclo del carbono y del oxígeno.
El ciclo del nitrógeno. El nitrógeno es sin duda un elemento muy importante para el metabolismo, ya que forma parte de las proteínas, los ácidos nucleicos y de muchas moléculas de gran importancia biológica. De manera similar a lo que ocurre con el carbono, el nitrógeno forma parte de un ciclo que hace a los diferentes tipos de organismos vivos depender entre sí. Esto se debe básicamente a que, a pesar de que el nitrógeno se encuentra distribuido en cantidades enormes en la atmósfera, es un elemento poco reactivo y debido a eso no puede ser aprovechado por la mayoría de las formas de vida. El nitrógeno debe ser asimilado por las plantas bajo la forma de nitrato o amoniaco, o bien por los animales bajo la forma de animoácidos, los cuales forman parte estructural de las proteínas.
La asimilación del nitrógeno en los ciclos de vida depende de varios factores. Las plantas toman el nitrógeno de la tierra en la forma de nitrato y lo utilizan para llevar a cabo las funciones metabólicas que dependen de este elemento. Por otra parte, los organismos dependientes utilizan las proteínas vegetales como nutrientes para aprovechar el nitrógeno contenido en éstas y lo devuelven a la tierra en forma de amoniaco como producto de desecho, o bien como producto de descomposición después de su muerte como se muestra en la figura 11. En la tierra, una especie de bacterias transforma al amoniaco en nitrato, el cual puede nuevamente ser utilizado por las plantas.
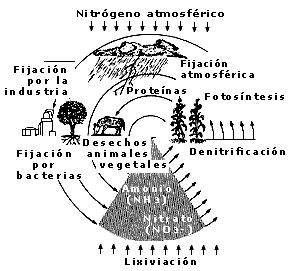
Figura 11. El ciclo del nitrógeno. Este ciclo mantiene el balance entre las dos vastas reservas de compuestos nitrogenados: la atmósfera y la corteza terrestres.
Existen, sin embargo, algunas especies de bacterias muy peculiares, conocidas como fijadoras de nitrógeno, que son capaces de procesar el nitrógeno atmosférico que por lo general no se asimila. Estas bacterias lo transforman en un producto útil para los organismos que dependen de él, como las plantas. Las bacterias fijadoras de nitrógeno atmosférico se asocian íntimamente con las raíces de algunas plantas. De esta forma, algunas, como el frijol, no requieren de ningún tipo de abono o fertilizante, ya que su asociación con la bacteria les permite utilizar en forma directa el nitrógeno atmosférico. Este tipo de simbiosis se antoja atractiva para que otras plantas, que requieren de fertilizantes, sean capaces también de asociarse con estas bacterias. Un gran número de investigadores en México y otras partes del mundo se encuentra interesado en este problema tan importante para la agronomía. Así, se están estudiando las posibilidades de establecer este tipo de interacción bacteria-planta en plantas que normalmente no lo hacen.
Hasta la fecha sólo se conoce un tipo de microorganismo que sea totalmente autosuficiente y que no dependa de ningún otro ser vivo, ya que utiliza la luz del Sol como fuente de energía, obtiene el carbono a partir del bióxido de carbono y el nitrógeno de la atmósfera. Estos organismos son las algas verdeazules, las cuales, se piensa, fueron los primeros pobladores de la Tierra cuando se establecieron las condiciones climáticas adecuadas.
II.3 LA ACTIVIDAD METABÓLICA AUMENTA O DISMINUYE
El metabolismo en los seres vivos es muy flexible y se puede ajustar a la cantidad y tipo de nutrientes disponibles en el medio que rodea a un microorganismo. Un ejemplo lo podemos obtener de las levaduras, las cuales se adaptan a las condiciones en que se les mantenga, pues poseen todo el conjunto de enzimas necesario para sintetizar sus elementos básicos como los aminoácidos y todos los demás componentes celulares, de tal manera que si crecen en un medio con glucosa como única fuente de energía, pueden sintetizar todas las moléculas que necesitan, aunque esto implica un gran gasto en energía. La levadura puede evitarlo si le proporcionamos los aminoácidos y nutrientes requeridos, ya que economiza el aparato enzimático responsable de la síntesis de todos los compuestos que necesita para vivir.
Actualmente es posible tener en el laboratorio cultivos controlados de bacterias en los cuales el investigador modifica a voluntad la composición de los nutrientes. Esto ha permitido conocer y entender cómo los microorganismos modifican su metabolismo en función de la calidad nutritiva del medio en el cual se les hace crecer. Si, por ejemplo, la concentración de nutrientes ricos en nitrógeno baja, el microorganismo iniciará inmediatamente la síntesis de las enzimas necesarias para elaborar sus propios ácidos nucleicos y aminoácidos; todo está finalmente regulado y enfocado hacia la máxima economía metabólica y, en última instancia, al ahorro de energía.
II.4 PARTES DEL METABOLISMO
El metabolismo se puede dividir en dos grandes partes: el catabolismo y el anabolismo.
La fase del metabolismo que descompone las moléculas grandes en pequeñas es el catabolismo. Las moléculas grandes como las proteínas, las grasas (lípidos) o los azúcares (carbohidratos), que provienen de nutrientes del medio ambiente, se descomponen enzimáticamente. El producto de esta descomposición o degradación lo forman las moléculas más simples y pequeñas como, por ejemplo, el ácido láctico, el ácido acético, el bióxido de carbono, el amoniaco y la urea. De estas reacciones químicas de degradación se obtiene la energía química contenida en las estructuras de las grandes moléculas. Esta energía se conserva en forma de molécula conocida como adenosíntrifosfato (ATP), la cual es de vital importancia en el metabolismo de cualquier organismo vivo.
Por otra parte, el anabolismo es la fase del metabolismo durante la cual se
sintetizan de nuevo las moléculas que la bacteria o célula utiliza para regenerarse,
mantenerse o dividirse, como son: las grasas, las proteínas, los azúcares o
carbohidratos y los ácidos nucleicos (ADN y ARN),
que forman parte funcional o estructural de los organismos. Esto lo lleva a
cabo el microorganismo o la célula a partir de los constituyentes primarios
que se obtienen de los nutrientes. Sin embargo, tales procesos de síntesis requieren
de energía y ésta la proporciona el ATP que fue generado durante
el catabolismo. Así, el anabolismo y el catabolismo se llevan a cabo simultáneamente
y cada uno está regulado en forma muy precisa, ya que ambos procesos son interdependientes
(Figura 12). Las enzimas que llevan a cabo las diferentes reacciones en el metabolismo
funcionan en forma secuencial, lo que quiere decir que el producto de una reacción
enzimática será a su vez degradado por la siguiente enzima de la cadena de reacciones
de alguna vía metabólica, proceso al que se conoce como de sistemas multienzimáticos.
La razón de este tipo secuencial de reacciones se debe, aparentemente, a los
múltiples puntos de control que ofrece, ya que un solo paso para la conversión
de una sustancia presentaría serios riesgos para un organismo; probablemente
por esta razón tal estrategia no ha sido adoptada por la naturaleza.
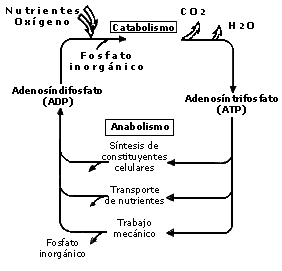
Figura 12. El anabolismo y el catabolismo. Todos los procesos que
ocurren en la célula o bacteria requieren de energía. Esta energía está
almacenada como moléculas de ATP, que se forma a partir
de ADP y fosfato inorgánico.
La degradación de los nutrientes: Las proteínas, los lípidos y los carbohidratos son degradados mediante reacciones enzimáticas que ocurren una después de la otra. Este proceso de degradación está dividido en tres etapas que a grandes rasgos son las siguientes:
La primera consiste en la conversión de moléculas complejas a moléculas más simples; esto quiere decir que una proteína, que no es más que una cadena de aminoácidos, tiene que ser convertida de nuevo en sus constituyentes, que son los aminoácidos, para que éstos puedan ser debidamente aprovechados en la segunda etapa de degradación. Estos procesos degradativos no requieren de energía y ocurren en el interior del microorganismo.
La segunda etapa consiste en la conversión de estas moléculas, ya bastante simples, en una aún más simple y común, independientemente del origen de las moléculas. Es decir, que tanto los carbohidratos como las proteínas o los lípidos son convertidos en una molécula mucho más simple llamada "acetil coenzima A".
Es a partir de esta molécula que la tercera etapa del metabolismo se lleva a cabo y consiste en generar la energía que necesita la célula para realizar procesos vitales, como desplazarse o dividirse, entre otros. En esta etapa ocurre uno de los ciclos metabólicos más importantes de la biología: el ciclo de los ácidos tricarboxílicos o ciclo de Krebs. Así pues, todas las moléculas que sirven a un organismo como fuente de subsistencia son llevadas, por medio de diversos caminos de degradación, a un camino metabólico común. Esta vía metabólica común degrada una sola molécula en una serie de pasos y como resultado se obtiene energía principalmente en forma de ATP. Los caminos metabólicos están finamente controlados. Estos mecanismos de control consisten en que los niveles de algunos productos regulan la actividad de algunas enzimas, de tal forma que su actividad se incrementa o disminuye dependiendo de los niveles del producto. Por otra parte, las concentraciones de ATP, que como ya vimos es una molécula muy importante en el metabolismo de la célula, regulan también la actividad de ciertas enzimas y esto lo hacen por medio de la unión del ATP a las enzimas susceptibles de ser reguladas por esta molécula. La regulación del metabolismo es vital, y de ésta depende que un un organismo produzca solamente la cantidad necesaria de cada una de las moléculas que requiere para subsistir y por lo tanto que el desperdicio de energía sea mínimo.
Tanto las bacterias como las células de los seres superiores generan la energía necesaria durante el catabolismo y la almacenan en forma de ATP. Así, éstas pueden realizar funciones vitales como el movimiento, el transporte de nutrientes a su interior y la síntesis de las moléculas que forman parte de su estructura o que tienen funciones específicas y deben ser sintetizadas en el interior. La síntesis de las moléculas es continua e implica un recambio constante entre las moléculas que se degradan y las que se sintetizan. De hecho, los procesos de degradación se conocen, aunque las señales que los gobiernan son aún tema de intensas investigaciones.
El estudio del metabolismo se ha apoyado en el uso de microorganismos a los cuales se les induce un cambio genético o mutación. Este tipo de enfoque ha permitido entender la mayoría de las vías metabólicas. Así, por ejemplo, el hongo Neurospora crassa puede crecer en un medio simple que contenga glucosa como única fuente de carbono y amoniaco como fuente de nitrógeno. Sin embargo, si se expone este hongo a rayos X, se obtiene una neurospora que ya no crece en el medio simple. Esta mutante solamente puede crecer en un medio de cultivo al cual se le ha añadido el compuesto que ya no sintetiza dicho microorganismo. Un ejemplo de esto son las mutantes del hongo que ya no crecen a menos que se añada al medio un aminoácido conocido como arginina (como todos los aminoácidos contiene nitrógeno). Lo cual quiere decir que la síntesis de este aminoácido está alterada y el hongo, por lo tanto, no puede utilizar el nitrógeno del medio para sintetizarlo. Esta mutante no crecerá a menos que se le adicione dicho aminoácido en el medio de cultivo. Existen toda otra serie de mutantes similares que difieren en los pasos en que el metabolismo se encuentra alterado; así se han podido conocer las diferentes etapas que forman parte de una vía metabólica.
II.5 LA MAQUINARIA DEL METABOLISMO
El metabolismo es, entonces, un sistema complejo de reacciones químicas llevadas a cabo por las enzimas que son las responsables de la transformación de los nutrientes en moléculas útiles para la bacteria.
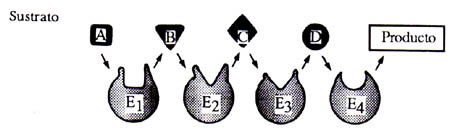 |
 |
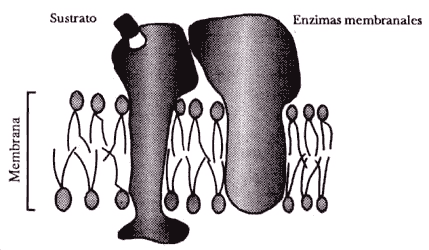 |
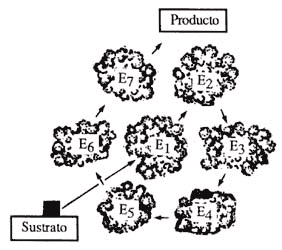
Figura 13. Sistemas multienzimáticos. Estos pueden ser tanto solubles como membranales, de su disposición depende su función.
La complejidad de los sistemas multienzimáticos que lleva a cabo el metabolismo es variable. En algunos casos las enzimas se encuentran en el interior de la célula y las moléculas que van a ser degradadas interaccionan libremente. Estos sistemas funcionan con base en la difusión libre de los productos del metabolismo (metabolitos) en el interior de la bacteria o célula. En otras palabras, la probabilidad de que una molécula interaccione con la enzima adecuada es alta y la reacción ocurre por lo tanto a una gran velocidad. Por otra parte, existen sistemas multienzimáticos organizados espacialmente en la membrana celular; esto permite que las reacciones ocurran de una manera organizada, funcionando juntos como una perfecta maquinaria engranada. Este tipo de disposición de las enzimas en una membrana puede llegar a ser muy complicada. De hecho, su funcionamiento es aún un problema biológico que atrae la atención de varios grupos de científicos en el mundo. Estos complejos enzimáticos altamente organizados forman parte indispensable de los organismos vivos desde las bacterias hasta los mamíferos. La interacción íntima entre sus componentes facilita la difusión de los metabolitos, y por lo tanto la velocidad a la que ocurren las reacciones es también muy alta. La figura 13 ilustra los tipos de sistemas enzimáticos y su disposición en el interior de la bacteria o de su membrana.
El metabolismo puede tener dirección y tamaño. Como ya vimos, existen sistemas multienzimáticos asociados a una membrana celular. Estos sistema generalmente aprovechan la división entre uno y otro lado de la membrana, es decir entre el interior y el exterior de la bacteria. Esta disposición permite dividir un compartimiento de otro, ya que las membranas celulares son impermeables.
La idea de que el metabolismo en compartimientos puede tener dirección y tamaño
parece complicado o sin sentido, y así fue como lo tomaron los bioquímicos en
los años sesenta. En 1961, Peter Mitchell, un microbiólogo inglés, propuso por
primera vez que la síntesis de la molécula conocida como ATP, se
debía a que ciertas reacciones metabólicas tenían como resultado el movimiento
de una especie química cargada o ión, en este caso un protón (H+).
Este protón, entonces, era el responsable de que el ATP fuera sintetizado.
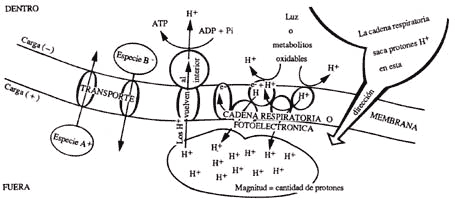
Figura 14. Algunas reacciones metabólicas tienen dirección y magnitud; estas reacciones se llevan a cabo en una membrana que funciona como barrera y que permite acumular una especie química, que al equilibrarse genera trabajo o energía.
La forma como funciona el complejo enzimático que sintetiza el ATP puede expresarse así: logra que un protón que se encuentra en el interior sea sacado al exterior. La figura 14 muestra en forma esquemática este proceso. Sumergidas en la membrana de la bacteria se encuentran las enzimas que componen la cadena respiratoria (así se les conoce, ya que al funcionar consumen oxígeno). La cadena respiratoria utiliza los compuestos que se producen durante el catabolismo en el ciclo de Krebs; éstos tienen la propiedad de ceder un electrón que con un protón formará un hidrógeno que ya no tiene carga. Este hidrógeno es transportado al lado opuesto de la membrana en donde pierde nuevamente el electrón y se libera el protón resultante al medio. Así, al cabo de varios ciclos se logra acumular una gran cantidad de protones en el exterior de la bacteria y esto tiene como consecuencia que se establezca una diferencia en la concentración de protones, que tiene la tendencia natural a equilibrarse. Un ejemplo podría ser la situación imaginaria de que un grupo grande de personas se encuentre en un pequeño cuarto comunicado con otro por una puerta que sólo se mueve en el sentido del cuarto vacío. Al estar tan apretadas empujarán dicha puerta y se empezará a llenar el cuarto vacío, y pasado un tiempo el número de personas será igual en ambos cuartos. Volviendo a la membrana de una bacteria, lo que ocurre es que la energía que se libera cuando se reequilibria la concentración de los protones en ambos lados de la membrana, permite que se sintetice el ATP.
Lo anterior, que hoy en día se ve claro, fue muy difícil de aceptar por la comunidad científica de la época y pasaron 16 años para que esta teoría fuera completamente aceptada. Su aceptación culminó con la entrega del premio Nobel a Peter Mitchell, ya que su teoría permitió explicar una gran cantidad de fenómenos hasta entonces oscuros. Por ejemplo, si estas reacciones con dirección producen una diferencia en la concentración de protones se tendrá también una diferencia de cargas (+) (afuera una gran carga positiva). Esto puede ser usado por algunas especies químicas para entrar, por ejemplo, si su carga es positiva (A+), o para salir si es negativa (B-). Como estos procesos metabólicos tienen dirección, magnitud y sentido, se dice que son "vectoriales"; además, son muy complicados y todavía no los comprendemos en su totalidad, pero baste decir que constituyen un pilar fundamental de los procesos energéticos celulares, ya que explican reacciones vitales como la síntesis del ATP o el transporte de nutrientes. Hasta ahora hemos visto a las bacterias que necesitan obtener su energía a partir de los nutrientes que se encuentran en el medio. Sin embargo, otro grupo de bacterias muy importantes son aquéllas cuya fuente principal de energía es la luz que proviene del Sol y de ellas nos ocuparemos a continuación.
II.6 BACTERIAS FOTOSINTÉTICAS
Menos familiares para nosotros, pero igualmente importantes, son las bacterias que tienen la facultad de utilizar la luz como fuente de energía. Éstas desempeñan un papel fundamental en la ecología y han servido para el estudio del proceso molecular de la fotosíntesis en numerosos laboratorios. Cuando se toman muestras de las profundidades de un lago, cualquiera se sorprendería al notar que el agua huele muy mal; además, hay que agregar el color verdoso o rojizo del agua. Si la analizamos con más detalle, veremos que hay producción de gas y que no existe oxígeno. Este ambiente tan hostil alberga a las poco conocidas bacterias fotosintéticas, responsables de una gran parte de la producción de materia orgánica en los lagos y parte importante de la cadena biológica. Estos organismos son muy antiguos y se piensa que son los descendientes directos de las primeras células fotosintéticas que existieron en la Tierra, entre las que se pueden contar las algas verdeazules y las bacterias fotosintéticas verdes y púrpuras. Las algas verdeazules se pueden encontrar como células únicas o en colonias en la tierra, los ríos, los lagos y los océanos. Estos organismos pueden vivir utilizando CO2- como única fuente de carbono y algunos son capaces de fijar el nitrógeno atmosférico.
Las bacterias fotosintéticas verdes que producen azufre (sulfurosas), como Chlorobium, viven en ambientes carentes de oxígeno, como en el fondo de algunos lagos ricos en materia orgánica. Estos organismos le dan el color verde característico a estas aguas. Así encontramos también bacterias púrpuras sulfurosas, como Chromatium, que viven en ambientes también carentes de oxígeno y que utilizan compuestos con azufre en su metabolismo. Éstas se encuentran también en algunos lagos y en manantiales de aguas termales sulfurosas. Finalmente tenemos a las bacterias púrpuras no sulfurosas, que tienen la peculiaridad de utilizar la luz en ausencia de oxígeno, como las bacterias verdes y las púrpuras sulfurosas, pero además pueden utilizar al oxígeno durante los periodos de oscuridad. Estas bacterias poseen en su interior una complicada organización de moléculas encargadas de captar la luz solar y conservar la energía proporcionada por esta forma de energía química. La luz que es aprovechada es la luz visible y gran parte de la cual es empleada también por las plantas verdes, de tal forma que los pigmentos encargados de captar la luz lo hacen en las regiones del espectro de la luz visible que no son utilizadas por las plantas. Estas moléculas se conocen como pigmentos antena y, dado que la intensidad de la luz que llega a las profundidades donde se encuentran estas bacterias es muy baja, su captación debe ser muy eficiente para así aprovechar la energía al máximo.
Estos microorganismos cuentan con estructuras internas que contienen las moléculas que captan la luz de tan baja intensidad. Estas estructuras se conocen como cromatóforos y contienen moléculas de clorofila típica de estas bacterias, que son las que captan la luz y retienen la energía que ésta proporciona (Figura 15).
Las bacterias fotosintéticas realizan una parte muy importante de la transformación y generación de la materia orgánica de lagos y ríos. Su existencia es determinante en el frágil equilibrio energético de este planeta. Estudiarlas es de gran importancia. En México existen ya algunos grupos interesados en estos microorganismos y en sus mecanismos de transformación y conservación de la energía solar.
La fotosíntesis. Éste es sin duda uno de los procesos más importantes de captura de energía en nuestro planeta y lo podemos definir como la forma de utilización de energía solar que han desarrollado algunos microorganismos fotosintéticos y las plantas, mediante la cual llevan a cabo la síntesis de todos sus componentes celulares. Este proceso es fundamental para todos los organismos vivos, ya que es la piedra angular del ciclo de utilización de la materia en la Tierra.
La luz solar es la fuente primordial de energía en la Tierra, tanto para los organismos fotosintéticos como para casi todos los organismos heterótrofos. Se utiliza a través de una complicada serie de procesos que van desde las cadenas alimenticias hasta complejos procesos industriales. Por ejemplo, el hombre utiliza la energía generada por la fotosíntesis de organismos que existieron hace millones de años y cuyos productos de descomposición se emplean en forma de gasolinas y derivados del petróleo, que proporcionan los combustibles para casi todas las máquinas construidas por el hombre.
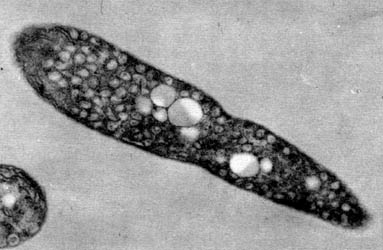
Figura 15. Micrografía electrónica de una bacteria fotosintética no sulfurosa, Rhodospirillum rubrum. En su interior se pueden observar pequeñas estructuras llamadas cromatóforos.
El descubrimiento de la fotosíntesis. El desarrollo histórico de la fotosíntesis se inicia entre 1770 y 1777 cuando Joseph Priestley, uno de los descubridores del oxígeno, llevó a cabo importantes experimentos sobre el intercambio de materia que ocurre durante la fotosíntesis. El experimento de Priestley consistió básicamente en consumir el oxígeno de un recipiente cerrado, lo que logró con una vela que ardía en el interior. Al consumirse el oxígeno, observó que la vela se apagaba. Por otra parte, si en dicho recipiente con aire se introducía además de la vela un ratón, al poco tiempo de consumirse el oxígeno de la cámara el animal moría. Ahora bien, si dentro del recipiente se incluía una planta, tanto la vela como el ratón se mantenían sin problema alguno. Priestley concluyó que las plantas verdes llevan a cabo la producción del oxígeno. Al parecer, este proceso era la inversa de la respiración en los animales, en la cual se consume oxígeno. A pesar de tan acertados experimentos, Priestley no logró determinar que este proceso estaba ligado a la luz. Fue Ingenhousz, un médico holandés, quien lo descubrió algunos años después, identificando que la porción verde de las plantas era la responsable de la producción del oxígeno.
Posteriormente, en el siglo XIX se concretó otra serie de importantes descubrimientos que condujeron finalmente a dilucidar que las responsables de la fotosíntesis en las plantas son las hojas y que hay producción de materia orgánica durante este proceso que se puede esquematizar como sigue:
La capacidad de llevar a cabo el proceso de fotosíntesis se encuentra en una gran variedad de organismos, tanto en bacterias como en plantas y algas.