VII. SISTEMAS MIXTOS: MOVIMIENTO GENERADO POR MICROTÚBULOS Y MICROFILAMENTOS
LA DIVISIÓN celular se inicia con el proceso llamado mitosis,
en el cual los cromosomas, duplicados, se separan y reparten en dos nuevos núcleos.
La separación de los cromosomas es seguida por la división del resto del citoplasma
o citocinesis. Cada célula hija tendrá entonces la información suficiente para
crecer y dividirse, a fin de continuar el proceso de proliferación. Por muchos
años los biólogos han estudiado la mitosis, describiendo desde hace más de 100
años las diferentes figuras producidas al interaccionar los cromosomas con estructuras
fibrosas y formar lo que se llama el huso acromático o aparato mitótico (Figura
VII. 1). Este huso, llamado así por su parecido a un huso de hilandera, cambia
de longitud, grosor, etc., durante la mitosis, y es sobre sus fibras en donde
los cromosomas parecen moverse para iniciar su segregación a lo que más tarde
serán las células hijas. Aunque la descripción detallada de la mitosis se hizo
hace muchos años, estas observaciones se limitaron a células fijadas, de modo
que reflejaban momentos de lo que realmente es un proceso dinámico y continuo.
Por otro lado, la caracterización bioquímica de las diversas estructuras participantes
en el proceso se logró apenas recientemente, de modo que hasta la fecha todavía
hay muchas incógnitas en relación con la mitosis.
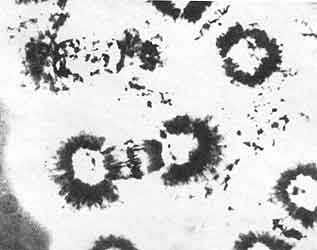
Figura VII. 1. Fotografía de los primeros aparatos mitóticos aislados por
D. Mazia y K. Dan. Los ásteres, el huso y los cromosomas en la placa ecuatorial
son claramente distinguibles. (Fotografía cortesía de D. Mazia).
Si consideramos el huso acromático como una máquina que mueve a los cromosomas debemos estudiar cómo está la máquina ensamblada para entender su funcionamiento. Empezaremos por analizar esta estructura. El huso no es algo permanente en las células. Se observa solamente durante la mitosis y se ensambla a partir de microtúbulos que a su vez, como vimos en capítulos anteriores, son polimerizados de las subunidades de tubulina. Una vez realizada la mitosis, los microtúbulos desaparecen para volverse a estructurar cuando la célula está lista para dividirse de nuevo. En algunas células, como las de un embrión, puede haber una división celular cada 10 ó 15 minutos. En otras, como las neuronas y los reticulocitos, que han llegado a un estado completamente diferenciado, no hay divisiones celulares (por lo menos en situaciones regulares, aunque experimentalmente éstas se logran inducir). En células en crecimiento activo la división celular se efectúa, aproximadamente, cada 24-48 horas y el proceso dura como una hora y media.
Como se vio en capítulos previos, los microtúbulos están formados por las tubulinas
(a y b) que al polimerizarse
forman una estructura con polaridad, esto es, con extremos asimétricos en donde
el ensamble y desensamble de subunidades procede con diferente cinética. Podríamos
imaginar que un microtúbulo es una flecha formada a su vez por flechas pequeñas,
todas alineadas con la punta orientada en la misma dirección y que por estas
características muestra propiedades diferentes en las puntas de la flecha o
en las colas. Se sabe actualmente que los microtúbulos del huso están orientados
con el extremo que crece más aprisa, llamado positivo, hacia el lado contrario
de donde está la estructura del huso llamada centrosoma (Figura VII. 2). Estas
propiedades facilitan la interacción diferencial con otras moléculas que se
asocian a los microtúbulos durante la mitosis, y que son capaces de transformar
energía química en movimiento, así como facilitar tanto la polimerización-despolimerización
de los microtúbulos como la unión de éstos a los cromosomas.
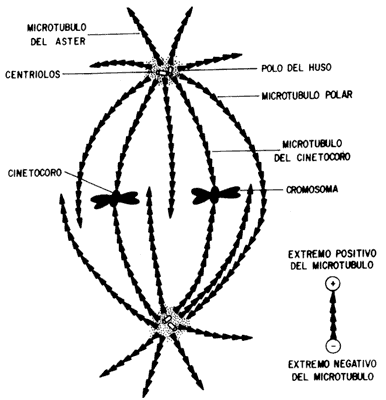
Figura VII. 2. Esquema del aparato mitótico. Las cabezas de flecha indican
la dirección en la que crecen los microtúbulos, que tienen como centros organizadores
a los ásteres.
Los centrosomas son otro elemento importante del huso. Estos se forman al inicio
de la mitosis a partir de un organelo formado por microtúbulos que existe permanentemente
en la célula, que es el centriolo. Cuando una célula va a dividirse este centriolo
se duplica, acción que se da, generalmente, cuando los cromosomas se están replicando
(Figura. VII. 3), y cada uno de ellos se mueve en el citoplasma para colocarse
en extremos opuestos, entre los cuales se formará el eje del huso acromático.
Alrededor de los centriolos, que realmente son centros organizadores de microtúbulos,
radian los microtúbulos del huso y aquellos que forman el centrosoma (Figura
I.1).
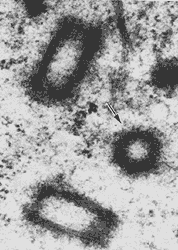
Figura VII. 3. Micrografía que muestra la organización de los microtúbulos
en los centriolos en una célula del oviducto de rata. En el corte transversal
(flecha) se puede apreciar el arreglo de éstos como nueve pares de tripletes.
(Fotografía cortesía de E. Dirksen).
Aunque los cromosomas no son propiamente parte del huso, cada cromátida (una
de las dos partes que forma a un cromosoma mitótico) tiene una región enriquecida
en proteínas que se conoce como el cinetocoro (Figura VII. 4). Esta región es
el punto de contacto entre los microtúbulos del huso y los cromosomas, y lo
que permite que cada cromátida se mueva por separado y se distribuya adecuadamente
a los nuevos núcleos.
Figura VII.4. Micrografía que muestra los microtúbulos del huso cuando hacen
contacto con la región del cinetocoro (cabezas de flecha) en los cromosomas.(Fotografía
cortesía de J. Pickett-Heaps).
Típicamente hay entre 15 y 35 microtúbulos uniendo a un cinetocoro con el centrosoma, aunque no todos los microtúbulos del huso llegan a los cinetocoros. Hay microtúbulos que se extienden de un centrosoma al otro y de los cuales depende la longitud del huso y otros más cortos que están interdigitados entre los microtúbulos más largos. Hace algunos años B. Nicklas, en la Universidad de Carolina del Norte, demostró que si se rompe la unión entre los microtúbulos y los cinetocoros, las cromátidas no pueden moverse, produciéndose una distribución anormal de los cromosomas. Estos experimentos hechos con un rayo láser, que rompe la unión microtúbulo- cinetocoro, demostraron también que el cinetocoro tiene polaridad y que su interacción con los microtúbulos de uno u otro centrosoma dependerá de su orientación. Esta especificidad garantiza que solamente una de las cromátidas se mueva hacia cada lado del huso, asegurando la distribución adecuada del material genético.
¿Cómo es entonces que estas estructuras logran el movimiento de los cromosomas?
Tratemos de integrar datos nuevos en la descripción clásica de la mitosis (figuras
VII. 5 y VII.6). Una vez que los cromosomas se han duplicado, cada uno de ellos
se mantiene unido, formado por dos cromátidas. El centriolo se ha duplicado
también al organizarse como centrosoma y dirige la polimerización de microtúbulos
a su alrededor. Algunos de estos microtúbulos interaccionan con las cromátidas
a través del cintocoro. A esta etapa se le llama prometafase. El cromosoma que
aun consiste de dos cromátidas, unido a uno o varios micrótúbulos, empieza a
moverse hacia el centrosoma del cual provienen los microtúbulos con los que
ha interaccionado. Esto se puede deber al acortamiento de los microtúbulos en
la base o a la interacción de una proteína específica en el cinetocoro con el
microtúbulo, que estaría inerte. El desplazamiento de un cromosoma duplicado
hacia un extremo se interrumpe cuando microtúbulos provenientes del centrosoma
situado en el extremo opuesto interaccionan con la otra cromátida y hay entonces
una fuerza que tira en el otro sentido. Este tirar en direcciones opuestas mueve
a los cromosomas a la zona ecuatorial del huso, dando por resultado la figura
llamada metafase. Es determinante la colocación de los cromosomas en el ecuador,
mediante su conexión con microtúbulos provenientes de los dos centrosomas, y
la mitosis prosigue sólo hasta que esto se ha logrado. En este momento las cromátidas
todavía están unidas entre sí y se desplazan hacia el centrosoma de donde provienen
los microtúbulos, con los cuales interaccionan. Se ha propuesto que el desplazamiento
de una cromátida hacia el centrosoma se hace por despolimerización de los microtúbulos
en sitio de unión con el cinetocoro, el cual practicamente se iría deteniendo
del extremo del túbulo que es cada vez más corto por la despolimerización progresiva
en ese polo. La cromátida se acercaría al centrosoma dando como resultado el
movimiento descrito en la anafase A.
Figura VII. 5. Esquema de la mitosis.
Por otro lado podría haber un componente con características elásticas, no
descrito todavía, que se uniera al cinetocoro y moviera a la cromátida,
utilizando a los microtúbulos como las vías de un tren. Este movimiento se haría
junto con la despolimerización los microtúbulos al nivel del cinetocoro. De
hecho se ha demostrado la presencia de actina y miosina en el huso acromático,
mediante el empleo de anticuerpos fluorescentes para visualizar a estas proteínas.
Sin embargo, aún no es posible concluir si estas proteínas podrían ser los elementos
elásticos en el movimiento de cromosomas.
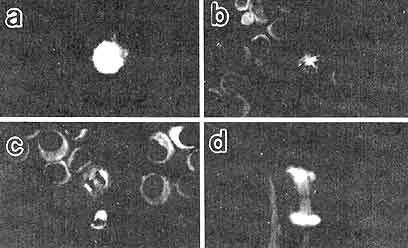
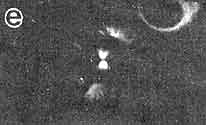
Figura VII. 6. Fotografías por microscopia de inmunofluorescencia en donde
se aprecian las fases de la mitosis esquematizadas en la figura VII. 5. Los
microtúbulos se ven decorados por un anticuerpo específico para tubulina, y
los cromosomas se muestran marcados con un indicador también fluorescente que
se une especificamente al ADN. a) Profase; b) placa
ecuatorial en la metafase: c) anafase; d) telofase, en donde pueden
verse los restos del cuerpo medio. (Fotografía cortesía de B. Brinkley).
En la anafase B los microtúbulos del huso que no están unidos a cinetocoros
crecen en su extremo positivo e interdigitan entre sí y se establecen puentes
laterales que forman otras proteínas asociadas a ellos. A través de estos puentes
los microtúbulos se repelen y se deslizan en direcciones contrarias, haciendo
que el huso en su conjunto se alargue, coadyuvando a la separación de las cromátidas.
Una de las proteínas identificadas como formadoras de puentes entre microtúbulos
es la cinesina. Este movimiento de microtúbulos que resulta del alargamiento
del huso puede hacerse en el laboratorio con husos aislados de varios tipos
de células. A mediados de los ochenta W. Zacheus Cande y sus colegas de la Universidad
de California, en Berkeley, aislaron husos acromáticos de células de algas diatomeas,
en presencia de ATP (el generador de energía), e investigaron el
movimiento de cromosomas en anafase. Encontraron que el movimiento dependía
de la fosforilación de proteínas unidas al huso y de la polimerización de los
micrcotúbulos en su extremo positivo. Esta elongación de microtúbulos ocurre
al mismo tiempo que el acortamiento de aquellos que directamente interaccionan
con los cinetocoros. ¿Cómo es posible entonces tener procesos diferentes y antagónicos
al mismo tiempo? Una posible explicación es que los dos tipos de microtúbulos
están inmersos en diferentes microambientes. Los que interdigitan están realmente
inmersos en la matriz citoplásmica, en donde pueden interaccionar con otras
moléculas y estabilizarse, de modo que pueden crecer cuando esto sea necesario.
Aquellos que interaccionan con los cinetocoros están, a su vez, interaccionando
con otro ambiente y dependen de otros mecanismos de control. Se sabe que el
calcio y las proteínas reguladoras de los niveles de Ca2+, como la
calmodulina, tienen un papel importante en el proceso de mitosis. Se ha encontrado
que las vesículas que almacenan calcio se concentran en las inmediaciones del
huso acromático y que hay, asimismo, por lo menos dos enzimas con actividad
hidrolítica sobre el ATP que son estimuladas por calcio.
Finalmente, una vez que las cromátidas llegan al extremo en que está el centrosoma,
durante la telofase, los microtúbulos que formaban el huso empiezan a despolimerizarse,
quedando un remanente de túbulos en la parte central de la célula que conecta
a los dos núcleos que ya han empezado a reestructurarse. La división del citoplasma
o citocinesis, que dará como resultado dos células hijas, empieza en la telofase
y se lleva a cabo por la estructuración de un anillo de constricción formado
por filamentos de actina y miosina en la parte media de la célula en mitosis.
Este anillo se forma a 90° del eje del huso acromático por polimerización
de monómeros de actina. La acción contráctil de los filamentos de actina y miosina
que se asocia a ellos constriñe al citoplasma y acaba por dividirlo (Figura
VII. 7). El anillo contráctil es una pequeña máquina que requiere de ATP
para hacer trabajo y puede estabilizarse y aislarse de las células. La formación
y contracción del anillo puede impedirse con drogas, como las citocalasinas,
que impiden la polimerización de la actina.
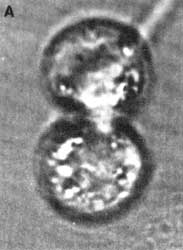
Figura VII.7. Células en citocinesis. A) Microscopia de contraste de fases;
B) diagrama del anillo de constricción. M = microtúbulos, c = cromosomas,
m= miosina, a= actina.
La membrana nuclear, que en la mayoría de las células se desestructura en la prometafase, se reorganiza en la etapa final de la telofase, alrededor de los cromosomas, para formar el nuevo núcleo. En los eucariones inferiores es común que la mitosis se lleve a cabo sin que la membrana nuclear se altere y que el huso acromático se organice con los ásteres en el citoplasma y las fibras del huso atravesando la membrana nuclear. En estos casos la mitosis no es muy típica y las fases no están bien definidas, pero básicamente el proceso debe depender de factores similares a aquellos que se presentan en una mitosis típica.
Se han podido contestar varias preguntas acerca de la mitosis, estudiando qué sucede cuando se altera el movimiento o la posición o formación de los centrosomas.
En relación al primer caso se han utilizado drogas, como la colchicina, vinblastina y la griseofulvina. Estos alcaloides, al unirse al dímero de tubulina, impiden que se añada al extremo de los microtúbulos que están creciendo, de modo que se interrumpe el equilibrio dinámico de polimerización de los microtúbulos. Al añadir este tipo de drogas a células en proliferación se detiene el proceso de la mitosis en metafase, de modo que los cromosomas quedan colocados en la placa ecuatorial, de donde no pueden proseguir a la separación de las cromátidas. Otras drogas, como el taxol, estabilizan a los microtúbulos ya formados y éstos no pueden despolimerizarse de nuevo, rompiendo el equilibrio dinámico entre polímero- dímero y teniendo como resultado la detención del proceso de mitosis.
La inestabilidad de los microtúbulos del aparato mitótico, requisito indispensable
para el movimiento por otro lado, impidió por mucho tiempo que esta maquinita
y sus componentes pudieran ser aislados y caracterizados. Fue en 1953 que D.
Mazia y K. Dan aislaron los primeros aparatos mitóticos de huevos de erizos
de mar que habían sido fertilizados in vitro. En estas células las mitosis
son sincrónicas una vez que empieza la división celular que da origen al embrión.
Estos investigadores pudieron aislar, en condiciones de estabilización de los
microtúbulos, un gran número de aparatos mitóticos y hacer su caracterización
bioquímica. Más tarde se estableció que los microtúbulos del aparato mitótico,
como otros microtúbulos citoplásmicos, están formados por las tubulinas a
y b. En el grupo de investigación del doctor Mazia
se caracterizaron inicialmente a las tubulinas que forman a éstos y otros tipos
de microtúbulos y se hicieron los experimentos que mostraron que el centriolo
es el organizador de los centrosomas y de los microtúbulos que radian a partir
de estas estructuras. Experimentalmente se pudo inducir la formación de múltiples
ásteres, fertilizando a los huevos de erizo de mar con varios espermatozoides,
cada uno de los cuales penetra al huevo con un centriolo, capaz de organizar
un áster y producir así un gran número de aparatos mitóticos (Figura VII.8).
Estas células no pueden continuar la mitosis por el caos que se produce al haber
tantos centros directrices del mismo proceso.
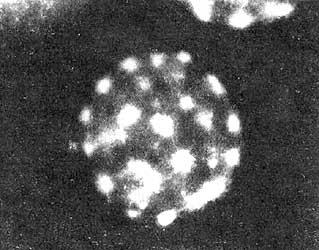
Figura VII.8. Múltiples ásteres en un huevo de erizo de mar producidos
por la fertilización con varios espermatozoides (polisperma). Los microtúbulos
que se originan de cada áster se identificaron al ser decorados por un anticuerpo
específico para tubulina. (Fotografía cortesía de G. y H. Schatten).
Recientemente se han identificado varias proteínas que podrían actuar como
motores moviendo a los cromosomas sobre los microtúbulos y que parecerían tener
un papel similar a la proteína cinesina mencionada en otro capítulo como un
motor celular. Esta proteína permite que los microtúbulos se desplacen unos
en relación con otros y muevan gránulos de pigmento o vesículas a lo largo de
una célula. El desplazamiento de los microtúbulos en el huso acromático se puede
llevar a cabo en formá similar (Figura VII. 9). El arreglo de los microtúbulos
en forma de huso, su regreso a microtúbulos dispersos en la interfase, la reorganización
de la membrana nuclear durante la mitosis, la condensación y descondensación
de los cromosomas y la formación del anillo de constricción, son todos procesos
controlados por acciones de fosforilación de varias de las proteínas
que forman estas estructuras o que ayudan al movimiento.
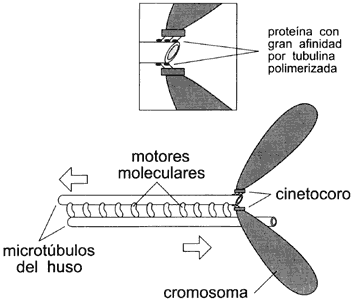
Figura VII.9. Esquema de la interacción de los microtúbulos con motores
moleculares (cinesina) para mover a los cromosomas.
Estas fosforilaciones se hacen por enzimas llamadas cinasas, la mayoría de las cuales son reguladas por los niveles de Ca2+ en el citoplasma y los cuales veremos que también regulan el grado de organización del citoesqueleto al asociarse a sus componentes e inducir su polimerización o despolimerización. Sin duda, aunque hay muchos elementos por identificar para entender completamente el proceso de la mitosis, los nuevos avances que pueden hacerse con la técnica de videomicroscopía, que permite observar el proceso dinámico, nos darán pronto una respuesta completa.