EL ORIGEN DE LAS ESPECIES
La especie es la unidad en que categorizamos a todos los seres vivientes, incluido el hombre (Homo sapiens). A pesar de que Darwin tituló su libro más importante con el encabezado de esta sección, y de que propuso a la selección natural como el mecanismo causante de los cambios que una especie puede experimentar, prácticamente no discutió el origen de las especies en sí y no dio ejemplos de cómo una especie puede dar origen a dos o más. Su esfuerzo más grande se centró en demostrar que las especies eran entidades muy variables, sin límites bien definidos, a las cuales la selección natural modifica constantemente, es decir, que las entidades reconocidas por el hombre como especies constituían unidades artificiales.
El concepto más adecuado para describir a una especie es el conocido como concepto biológico de especie, que la define como una población o serie de poblaciones de organismos entre los cuales ocurre un flujo genético libre en condiciones naturales, es decir, que los individuos de esa (s) población (es) pueden cruzarse libremente y tener progenie fértil en las condiciones del ambiente en el que viven. Por contraposición, los individuos de una especie no pueden cruzarse libremente con los de otra.
El hecho de que el cruzamiento entre los individuos debe darse en condiciones naturales es de especial importancia, ya que lograr que dos individuos pertenecientes a especies diferentes produzcan progenie en condiciones controladas por el hombre, no demuestra que se trata de la misma especie. El ejemplo mejor conocido al respecto es el de la cruza en zoológicos de dos especies que consideramos diferentes, como leones y tigres, los cuales son dos especies cercanamente relacionadas del género Panthera (P. leo y P. tigris). Se han obtenido, por medio de inseminación artificial, "tigrones" (cruza de tigre y leona) y "leogres" (cruza de león y tigresa). Lo anterior no quiere decir que leones y tigres pertenezcan a la misma especie, ya que hay una serie de elementos que impiden que ellos se crucen en condiciones naturales. Sus áreas de distribución son muy diferentes: el león es fundamentalmente africano y el tigre, asiático. Aunque dichas áreas se sobreponen en el noroeste de la India, nunca se ha encontrado un tigrón ni un leogre en la naturaleza porque los hábitos de ambas especies son también muy diferentes. Los leones tienen hábitos sociales, forman familias, cazan en grupo y habitan terrenos abiertos, como las sabanas. Los tigres, por el contrario, son de hábitos solitarios y prefieren ambientes selváticos o boscosos.
El concepto de especie no es un invento de los intelectuales europeos del siglo XVIII Responde a una realidad biológica, generalmente expresada por la forma, los hábitos y las características morfológicas de los organismos. Ya nos referimos en el capítulo VIII al profundo conocimiento que los pueblos nativos de Mesoamérica tenían, mucho antes del contacto cultural con los europeos, de los animales y las plantas que los rodeaban. Esto ocurre igualmente con otros pueblos de América, Asia y África. No se trata de un conocimiento que consista en "saberse de memoria" varios cientos o miles de nombres de plantas y animales, sino que es un conocimiento estructurado en sistemas lógicos y coherentes, basados en las afinidades o diferencias de las características de los organismos, no muy diferentes del sistema linneano y muy de acuerdo con el concepto biológico de especie.
La distinción de un grupo de organismos en una especie implica que para que haya alcanzado un grado de aislamiento reproductivo de las especies afines ha requerido un largo periodo de tiempo en el que pudo diferenciarse y adaptarse al ambiente en que vive. También implica que no hay una sola característica morfológica que la distinga de las especies cercanas, sino un conjunto de ellas, además de aspectos fisiológicos, de comportamiento, de distribución, etcétera.
El concepto de especie se enfrenta a pocos problemas cuando consideramos una población muy pequeña, en una reducida área de distribución y en un tiempo muy breve. Apenas empezamos a ampliar nuestra visión para incluir numerosas poblaciones, en una área muy amplia de distribución y en una escala temporal que incluya por lo menos varias generaciones de individuos, el concepto de especie empieza a perder fortaleza y a enfrentar serios problemas. El primero es que cuando consideramos poblaciones separadas geográfica o temporalmente, no sabemos con certeza si podrían cruzarse en condiciones naturales, lo cual es una premisa del concepto biológico de especie. En ambos casos de separación, pero particularmente en la temporal, se establecen grados de variación tan cercanos que resulta imposible definir dónde podría marcarse el límite entre dos entidades. Éste es el caso de muchas especies de fósiles que tienen representantes vivos morfológicamente muy semejantes y la delimitación entre especies en estos casos tiene un propósito más bien práctico. Si la base del concepto biológico de especie es el libre intercambio de genes entre individuos, entonces las especies que se reproducen por medios asexuales (como las bacterias) o cuyos individuos se reproducen sexualmente pero son estrictamente autofecundados, deben ser delimitadas con otros criterios, que generalmente son pragmáticos. Los casos de intergradación continua de poblaciones e individuos son muy frecuentes, particularmente entre plantas, y representan también un serio problema para definir fronteras intraespecíficas. Nuestro país es un cofre de tesoros en lo que a estos problemas se refiere, por su ubicación geográfica, su topografía, su geología y sus climas. Ejemplos notorios de grupos "problema" de organismos son varios complejos de especies de pinos, así como la mayoría de los encinos o robles mexicanos que pertenecen al género Quercus. De ambos géneros México posee el mayor número de especies en el mundo.
Las especies están subdivididas en la naturaleza en poblaciones más o menos bien definidas en las que los individuos se cruzan. A medida que la distribución de las especies es más amplia, las poblaciones de una especie se encuentran con una mayor variedad de ambientes. Esta diversidad propicia que las mutaciones que se produzcan tengan mayor probabilidad de ser seleccionadas favorablemente y fijadas en cada condición, de manera que la diversidad genética de las poblaciones aumenta con su área de distribución. Si el flujo genético entre las diferentes poblaciones es pobre, la divergencia entre los contingentes genéticos de las poblaciones puede aumentar lo suficiente como para crear barreras al cruzamiento o a la fertilidad entre ellas, incluso si llegasen a ponerse en contacto nuevamente por algún cambio ambiental. La reducción de la fertilidad se origina por varias causas, entre ellas la incompatibilidad genética, las diferencias en comportamiento, el aislamiento temporal por diferencias en épocas reproductivas, etc. Las barreras al cruzamiento y el aislamiento reproductivo no ocurren súbitamente ni en un solo individuo; son el producto de cambios acumulativos que toman mucho tiempo y que ocurren en toda una población.
Las barreras reproductivas pueden ocurrir sencillamente por un aislamiento
geográfico entre poblaciones de subespecies o de especies muy cercanas producido
por barreras físicas tales como cadenas montañosas o cuerpos de agua; aunque
también se presentan en especies que coexisten en el mismo sitio. En este caso
las barreras reproductivas son el resultado de diferencias genéticas entre los
miembros de las dos especies cercanas y que impiden la fertilización o el desarrollo
normal del embrión, o que de cualquier otra manera impiden la producción de
progenie viable y fértil entre las especies. Algunos ejemplos de estas barreras
genéticas son las diferencias de comportamiento ya mencionadas para los leones
y los tigres; el hecho de que el vuelo nupcial de dos especies de abejas ocurra
a horas diferentes del día; la imposibilidad de que los espermatozoides o el
polen alcancen a fertilizar el óvulo debido a barreras mecánicas o químicas
en el aparato reproductivo, etcétera. Las barreras reproductivas están presentes
en todas las especies, incluida la humana. A pesar de su gran movilidad, de
los grandes movimientos migratorios y del hecho de que en teoría una persona
de cualquier sexo pueda escoger como pareja a cualquiera otra del sexo opuesto
en el mundo, la realidad es que en su mayoría la selección de parejas en la
especie humana ocurre en áreas infinitamente pequeñas, tales como el barrio
en el que una persona vive. Diversos estudios de larga duración prueban lo anterior,
y es fácil convencerse de que factores de tipo social, racial, religioso o cultural
influyen mucho en la selección de la pareja en contingentes bien definidos y
discretos.
 15
15
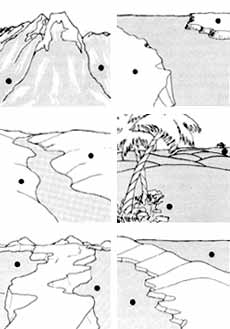
Distintos tipos de barreras geográficas que pueden aislar poblaciones de
una misma especie que, al diferenciarse, pueden llegar a constituir dos especies
distintas.
Cuando los estudios de Mendel fueron redescubiertos a principios de nuestro siglo, se produjo una corriente de pensamiento entre los genetistas y los biólogos que no aceptaban la idea de la evolución de los organismos como Darwin la había propuesto, es decir, basada en una lenta acumulación de cambios imperceptibles. Los resultados del monje agustino, obtenidos del estudio de caracteres sencillos y contrastantes, sugerían que los cambios en las especies no ocurrían en forma gradual, sino en saltos o pasos bien marcados y definidos, ejemplificados por los contrastantes cambios en talla o color de las flores de los chícharos de Mendel. Uno de los redescubridores de tal trabajo, Hugo de Vries, acuñó el término de mutaciones para los cambios súbitos que observó en sus experimentos de hibridación con las prímulas. Aunque los cambios observados por De Vries eran más bien la expresión de caracteres recesivos de las plantas, el término de mutación se conservó para describir los cambios súbitos y heredables en la estructura genética de los organismos. Para los genetistas mendelianos de principios de siglo, estas modificaciones súbitas del genotipo de los individuos eran las causas determinantes de la evolución de las especies, la cual ocurría en un constante proceso de saltos, como si la evolución fuese un chapulín. Como veremos un poco más adelante, ocurrió una conciliación entre la corriente evolucionista "saltatoria" y las ideas darwinianas de cambio paulatino basadas en la variabilidad genética de los organismos.
A pesar de la interpretación equivocada de De Vries, antes mencionada, es claro que sí ocurrían cambios abruptos en la estructura genética de las especies (que incluso el hombre ha aprovechado en sus prácticas de domesticación de animales y plantas), por lo que la transformación de las especies no ocurre exclusivamente por la lenta acumulación de cambios imperceptibles. Darwin mismo acabó por aceptar que tales cambios podían también contribuir a la evolución de las especies, aunque afirmando que lo hacían de forma secundaria.
Las mutaciones son de dos tipos: cambios pequeños en porciones bien definidas de la estructura del ADN, generalmente en un par de bases, a los que se llama mutaciones puntuales, y cambios en grandes secciones del ADN, o incluso en el cromosoma entero, que reciben el nombre de mutaciones cromosómicas. No entraré a describir con mayor detalle las diferentes formas por las que dichas mutaciones se producen durante la replicación del ADN o de la meiosis, y sólo mencionaré que ambos tipos pueden ser provocados por radiaciones o por ciertas sustancias químicas mutagénicas, aunque también pueden ocurrir espontáneamente. Muchas de esas mutaciones, particularmente las de tipo cromosómico, son letales y no producen progenie viable o si lo hacen no se pueden seleccionar favorablemente en una población. Un ejemplo de esto último es el caso del mongolismo o síndrome de Down en la especie humana, resultante de la existencia de un cromosoma extra en el par 21, lo cual aumenta el número cromosómico a 47, en lugar de 46. Otras mutaciones sí pueden representar un cambio favorable y por lo tanto ser seleccionadas positivamente. Si el cambio es suficientemente importante para darle una gran ventaja al mutante, éste pronto dominará en una población, siempre y cuando las características ambientales para las cuales es particularmente apto permanezcan sin modificación. Otras mutaciones son neutrales y no tienen valor adaptativo alguno, pero pueden llegar a fijarse y a formar parte del contingente genético de la población, particularmente si se trata de poblaciones pequeñas.
Otro mecanismo de producción de especies es la llamada poliploidia, un accidente en el proceso de la meiosis que produce un tipo de mutación cromosómica consistente en la multiplicación exacta del número de cromosomas de una especie; este mecanismo es muy frecuente en plantas y muy raro entre los animales. Los gametos, células especializadas para la reproducción de las especies (espermatozoides, óvulos o polen), poseen un número básico (haploide) de cromosomas. El resto de las células del organismo de la mayoría de las especies tienen un número diploide de cromosomas; la poliploidia duplica (produciendo tetraploides, es decir, cuatro veces el número haploide de cromosomas), triplica (produciendo hexaploides) o aun multiplica más el número cromosómico diploide. Consideremos por ejemplo una planta de una especie con un número cromosómico de 14; cada célula tendrá dos copias de siete cromosomas (que es el número haploide). Durante la meiosis los dos cromosomas de cada tipo deberán aparearse para completar el proceso de formación del gameto. Cada gameto debe tener un cromosoma de cada tipo, es decir, un total de siete cromosomas. Supongamos que, debido a un accidente durante el proceso de la meiosis, algunos nuevos individuos portan el doble de cromosomas, es decir cuatro de cada tipo, en lugar de dos, lo que daría un total de 28 cromosomas. Estas plantas tetraploides pueden producir gametos que contendrán 14 cromosomas en lugar de los siete del número original haploide de la especie, y podrán cruzarse para producir progenie fértil con 28 cromosomas. Pueden también cruzarse con gametos haploides para formar híbridos con un número total de 21 cromosomas (7 + 14). Sin embargo, estos híbridos triploides no pueden producir gametos normales, debido a que al inicio de la meiosis en el momento del apareo de los cromosomas, existirán tres en lugar de dos cromosomas de cada tipo para aparearse, por lo que se produce una distribución anómala de los cromosomas en las células hijas. Por lo general, esto es suficiente para interrumpir la meiosis, por lo que el híbrido se vuelve total o parcialmente infértil. Si esto ocurre, las formas diploides y las tetraploides se habrán aislado y constituido dos nuevas especies en un solo paso generacional.
Otra forma importante en que la poliploidia puede ocurrir exitosamente es cuando dos especies diferentes se cruzan y producen un híbrido que resulta estéril debido a diferencias marcadas entre los cromosomas de las dos especies, lo cual no permite un apareamiento adecuado en el momento de la meiosis. Sin embargo, si en un accidente meiótico el híbrido logra duplicar exactamente el número de sus cromosomas, no tendrá problema para realizar de ahí en adelante exitosamente la meiosis, ya que contará con dos cromosomas de cada tipo proveniente de cada uno de los padres, lo que le permitirá llevar a cabo una meiosis normal, aunque ahora con el doble de cromosomas. El nuevo híbrido poliploide resultará ser una nueva especie, diferente y aislada de sus progenitores.
Una forma más en la que se pueden producir cambios en la estructura genética de una población susceptibles en ciertas circunstancias de llevar a la formación de razas, subespecies e incluso especies nuevas, es el fenómeno conocido como deriva genética. Ésta ocurre cuando una característica o una mutación neutral se produce en una población pequeña, que se aísla de un contingente más grande por alguna catástrofe ambiental o por el efecto de colonización de una nueva área por uno o muy pocos individuos. Por ejemplo, la fijación de una mutación neutral o un gene en una población requiere un número de generaciones que es una proporción de cuatro veces la población reproductiva efectiva. Un gene dado o una mutación neutral se fijará en una población de dos aves que colonizan una nueva isla en sólo ocho generaciones, siempre y cuando todos los individuos de la progenie sobrevivan. En una población de 15 000 aves, tomará 90 000 generaciones fijar el mismo gene o mutación neutral. En otras palabras, el hecho de que el contingente genético de una población surja de un número reducido de individuos (que solamente representan una pequeñísima muestra del contingente genético de la población de la que se separaron, ya no digamos de su especie), puede producir cambios muy considerables en la constitución genética de la nueva población comparada con la original. Si las diferencias son suficientemente fuertes y se mantiene un aislamiento geográfico prolongado, puede darse origen a una nueva especie.
15 Charles Darwin, El origen de las especies, México, Consejo Nacional de Ciencia y Tecnología, 1982.