IV. LAS REVELACIONES DE LA MOLÉCULA MAESTRA
Consecuencias de la revolución metodológica en la adquisición de conocimiento.
EN LAS secciones anteriores he intentado proveer al lector de
un marco conceptual adecuado para comprender de una manera más precisa las múltiples
aplicaciones del ADN recombinante. Quizá con el material proporcionado
hasta el momento se pueda tener una noción de la potencialidad extraordinaria
de esta metodología. Este capítulo y el siguiente estarán orientados a describir
algunas de las áreas en las que las aplicaciones del ADN recombinante
han tenido influencia. Trataré de concentrarme en aquellas que resultan más
evidentes, interesantes o potencialmente importantes por su magnitud en nuestra
concepción del mundo o en nuestra forma de vida. Quisiera hacer incapíe en que
esta relación de aplicaciones es, por necesidad, extremadamente limitada: la
ingeniería genética ha influido virtualmente en todo el espectro de la investigación
biológica experimental.
Recuerdo haber leido una aseveración hecha por un científico unos pocos años después de que se iniciara la epidemia mundial del SIDA, quien afirmaba que si esta enfermedad hubiera hecho su aparición antes del surgimiento del ADN recombinante, seguiríamos buscando su causa debajo de las piedras.
A riesgo de ser repetitivo, quisiera también hacer comentarios acerca de mi perspectiva personal sobre la importancia del ADN recombinante. Mi experiencia en el campo de la investigación científica se inició a finales de la década de los setenta. En esos años la ingeniería genética estaba en su infancia, y se empezaba a establecer en el Instituto de Investigaciones Biomédícas de la UNAM. Cuando reparo en una serie de aspectos relativos al quehacer científico tal como eran hace 15 años, y los comparo con la situación actual, no ceso de sorprenderme. Cuando reviso (como una de mis actividades semanales) el cúmulo de conocimientos que aparece en la literatura científica, ésta es comparable a un verdadero alud. Los resultados que se reportan en un solo artículo habrían requerido de varios años de trabajo hace una década. Me parece que un cálculo conservador podría indicar que la velocidad a la que se pueden desarrollar nuevos conocimientos en biología experimental es unas 10 veces mayor de la requerida en el momento que inicié mi trabajo como científico. Estamos en un punto en el que quizá la limitante empieza a ser la capacidad de la comunidad científica para asimilar e integrar los conocimientos generados. Es posible, también, que estemos cercanos al punto en el que la velocidad de crecimiento en la acumulación de conocimientos empiece a disminuir por razones económicas: ¡se requerirá destinar una proporción excesiva de recursos humanos y materiales para sostener el paso que llevamos actualmente!
Tomemos algunas muestras de los avances que hago mención.
El inicio del estudio sobre la naturaleza de los genes se ocurre, naturalmente, en algunos de los organismos más simples como las bacterias. En términos generales, el estudio de todo tipo de microorganismos se ha convertido en una actividad con múltiples facetas. La experimentación tiene hoy, gracias al ADN recombinante, dimensiones completamente nuevas.
Una vez aislado e identificado un gene, podemos alterarlo y reintroducirlo a la célula bacteriana para observar las consecuencias de la alteración. Desde los albores de esa era de la ingeniería genética, al final de los años setenta, se empezaron a acumular datos acerca de genes microbianos, y la estructura y función del gene se fue clarificando y cobrando detalles (recuadro lV.I).
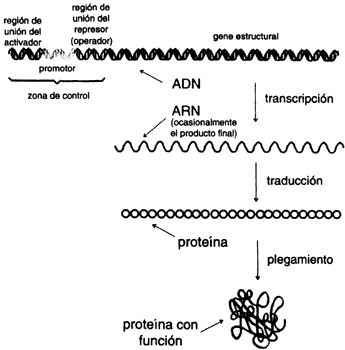
Por ejemplo, para ilustrar el análisis de la regulación de un gene, tomemos un experimento simple. Supongamos que queremos saber en qué condiciones se activa la expresión de un conjunto de genes que participan en la respuesta al calor de una bacteria. Estos genes pueden formar parte de un complejo circuito, y sería difícil seguir la actividad de cada uno por medio de su función natural. En cambio, lo que podemos hacer gracias al ADN recombinante es construir un gene mixto por cada gene que deseamos estudiar asociando a la región regulatoria otro gene estructural que sí sea fácil de seguir. Esto es lo que llamamos un gene reportero. Por ejemplo, podemos usar un gene que codifique para una enzima que actúe en un sustrato sin color y que al combinarse lo convierta en un producto colorido. De hecho actualmente se dispone de varios genes reporteros con las características antes descritas. El gene de la ß-galactosidasa, uno de los genes caracterizados en las épocas pioneras de la biología molecular; ha sido utilizado con este propósito. (Curiosamente esta misma enzima, la ß-galactosidasa, es responsable de que los seres humanos digiramos la lactosa de la leche, y al desactivarse su expresión, en muchos adultos se desarrolle la intolerancia a la leche.) Pues bien, si introducimos a la célula bacteriana un gene quimérico o mixto, cuya región regulatoria está relacionada con la respuesta al calor, y la región estructural codifica para la ß-galactosidasa, observaremos la respuesta que normalmente tendría el gene de interés, simplemente advirtiendo el cambio de color de las colonias transformadas.
Por medio de estudios en los que se alteran varias partes de los genes y se prueban en diversas condiciones, existe hoy día una idea extremadamente elaborada acerca de las formas como se regulan los genes que intervienen en todo tipo de procesos. Sin embargo, distamos mucho de haber comprendido satisfactoriamente, incluso los procesos de regulación más simples.
INMUNOLOGÍA Y ENFERMEDADES INFECCIOSAS
Algunos de los fenómenos biológicos que resultan más familiares son los que se manifiestan en el curso de las enfermedades infecciosas. En este tipo de situaciones se observan interacciones entre el agente infeccioso y el enfermo que se han podido comprender bien en el nivel molecular. La descripción de algunos aspectos básicos sobre el funcionamiento del sistema inmunológico, así como de lo que entendemos acerca de ciertos mecanismos de patogenicidad puede ilustrar muy bien el papel que desempeñan las moléculas biológicas en ellos y cómo se ha podido enfocar su estudio mediante el uso del ADN recombinante.
Los anticuerpos y cómo se producen
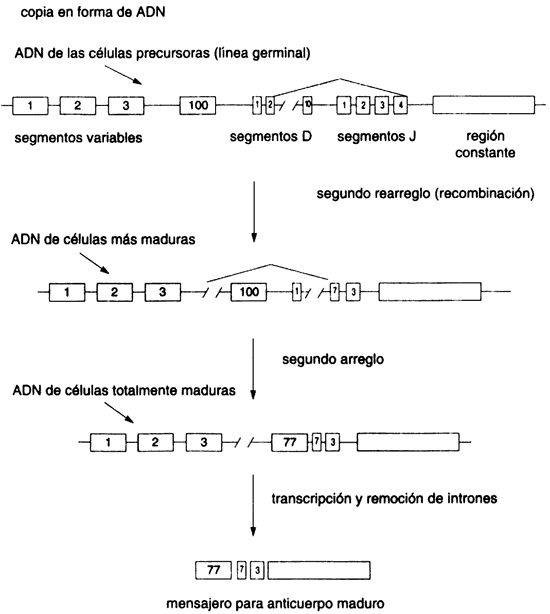
Cuando una sustancia extraña invade el organismo se desencadena una serie de reacciones de defensa, fundamentalmente para establecer que esa sustancia no es propia. Uno de los principales componentes de este sistema lo constituyen los anticuerpos, que se unen y "marcan" la sustancia extraña para su posterior destrucción. Estas sustancias son (quizá el lector lo recuerde) proteínas. Los anticuerpos llamaron la atención de los científicos desde hace muchas décadas. El acceso a ellas había sido relativamente fácil, dado que son abundantes en la sangre. Los primeros experimentos de separación de los componentes de la sangre revelaban varias manchas numerosas en el patrón electroforético (véase en el capítulo 1, "Separación y análisis de las proteínas", ), las cuales se denominaron globulinas. Una de ellas, la que correspondía a lo que se llamó a-globulinas, agrupaba los anticuerpos hoy nombrados inmuno-globulinas G. Muy pronto, después de que se identificó que estas sustancias eran los anticuerpos, los inmunólogos se preguntaban cómo era posible que el organismo fuera capaz de generar tal multitud de proteínas, como para ser capaces de reconocer cualquier sustancia extraña que nos invadiera. Más aún, ciertas células cancerosas, llamadas mielomas, se pueden obtener en gran cantidad, y su análisis reveló que cada una de ellas producía uno, y sólo un tipo de anticuerpo. Por el dogma central de la biología molecular se infería que cada anticuerpo era el producto de un gene y esto hacía surgir una paradoja: dados los millones de anticuerpos diferentes que se sabía que existían en un organismo dado, ¡los genes que codifican para ellos deberían ocupar todo el genoma del organismo! Diversas hipótesis fueron avanzadas para explicar lo que sucedía. Como es frecuente, hubo alguien que intuyó la respuesta correcta: los inmunólogos Dreyer y Benett propusieron una osada hipótesis en 1965. Dado que se sabía que los diversos anticuerpos eran muy parecidos entre sí y sólo diferían en una pequeña parte de su secuencia, ellos pensaron que lo que podría estar ocurriendo es que existiera sólo un gene para la parte constante y muchos genes para las pequeñas partes variables. Esta hipótesis significa que el genoma tendría que rearreglarse para poder generar los genes maduros, que codificarían para el anticuerpo particular que una célula dada expresaba. Esta hipótesis fue muy mal recibida porque se pensaba que el arreglo del genoma era muy estable.
La respuesta a esta interrogante sólo surgió una vez que se dispuso de las
técnicas de ADN recombinante. Entonces fue posible realizar una
serie de experimentos, que le hicieron ganar a Susumu Tonegawa el Premio Nobel
de fisiología o medicina otorgado en 1987, en los que se demostró el funcionamiento
de uno de los procesos más complejos de la evolución biológica. En su demostración,
Tonegawa y sus colaboradores utilizaron la propiedad de hibridización de ácidos
nucleicos para mostrar que el ARN mensajero purificado de un mieloma
es capaz de asociarse a un solo pedazo de ADN obtenido del mieloma,
pero se une a dos pedazos de ADN proveniente de cualquier otra
célula. En efecto, el ADN estaba realizando un reacomodo para los
genes de las inmunoglobulinas. El trabajo sucesivo de Tonegawa, y muchos otros
investigadores, estableció lo que hoy conocemos con bastante precisión respecto
a la biosíntesis de los anticuerpos (véase el recuadro IV.2). En este proceso
el organismo genera un variadísimo repertorio de anticuerpos, a ciegas, de una
manera muy eficiente, para después hacer proliferar sólo a las células que son
capaces de manufacturar el anticuerpo útil en un momento dado, es decir, cuando
se presenta el antígeno, o sustancia extraña.
En los últimos 15 años se ha acumulado una enorme cantidad de información acerca del funcionamiento del sistema inmunológico. No sólo conocemos la secuencia de aminoácidos de miles de diferentes anticuerpos y de sus genes, sino que se han identificado un sinnúmero de factores adicionales que modulan la compleja serie de interacciones que ocurre en una respuesta inmune. Este conocimiento ha servido como base para el uso de anticuerpos en la terapéutica del futuro inmediato, así como para crear una base firme en el diseño de nuevas vacunas (véase el capítulo siguiente).
Estudio de agentes infecciosos
Quién no recuerda haber oído hablar de las épocas pioneras en que Luis Pasteur y los otros cazadores de microbios rompieron moldes demostrando la existencia de microorganismos, que eran los responsables de la putrefacción y de muchas enfermedades. Efectivamente, la microbiología es un gran pilar de la investigación biológica experimental, y como ya mencionamos, sentó incluso las bases de la genética molecular. El avance de estas investigaciones, sin embargo, no escapó a los límites de la metodología disponible, encontrándose con obstáculos difíciles de sortear hasta el surgimiento del ADN recombinante.
Pensemos, por ejemplo, en lo complicado que resulta trabajar con una bacteria o un virus patógeno. Se requiere cultivarlo en grandes cantidades, lo que entraña un gran riesgo para el experimentador. Además, muchos organismos patógenos sólo se reproducen en condiciones muy especiales, como en el interior de su organismo hospedero. La posibilidad de donar sus genes abre, sin embargo, innumerables de posibilidades.
Los agentes infecciosos están siendo estudiados activamente desde diversos ángulos. Los genes responsables de la patogenicidad de muchas bacterias han sido aislados y secuenciados; se han podido comparar los genes de cepas patógenas con los de cepas no patógenas; se han identificado, a nivel fino, los componentes de la superficie de agentes infecciosos que les permiten evadir la respuesta inmune... En fin, la lista podría seguir por varias páginas.
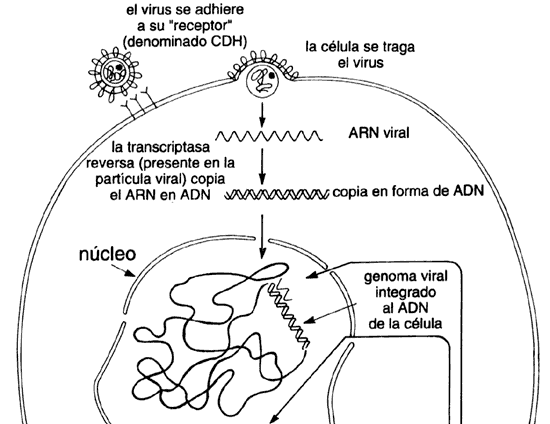
Tomemos, por ejemplo, el virus de la inmunodeficiencia humana (VIH) que produce el SIDA. El cual sólo se encuentra en ciertas células de la sangre: una clase particular de linfocitos (glóbulos blancos) del tipo T. De un enfermo sólo se pueden aislar cantidades minúsculas del virus. Sin embargo, gracias a la ingeniería genética, a sólo unos pocos años de haber sido identificada la enfermedad, se han hecho grandes avances. La inmensa mayoría de los estudios que se han realizado alrededor de este virus emplean ADN recombinante, y han generado un vasto conjunto de conocimientos, que han permitido su diagnóstico y sentado las bases para lograr su prevención y tratamiento. Aun cuando dicho tratamiento no es todavía una realidad, todo parece indicar que el control de esta enfermedad se logrará en un tiempo récord, particularmente si se toma en cuenta lo increíblemente difícil de conocer y atacar, y lo elusivo que resulta ser el virus que la causa.
Específicamente, a pesar de que el virus del SIDA sólo se encuentra
en pequeñas cantidades dentro de las células de enfermos humanos (es decir que
no se puede cultivar), desde hace varios años se conoce la secuencia completa
de su genoma de 9200 bases. Asimismo, se han identificado todos y cada uno de
lo genes que lo constituyen y les han asignado funciones a la mayoría de ellos.
Su ciclo de vida ha sido disecado en sus aspectos fundamentales, haciendo referencia
a otros virus similares (de la familia de los retrovirus, que pueden causar
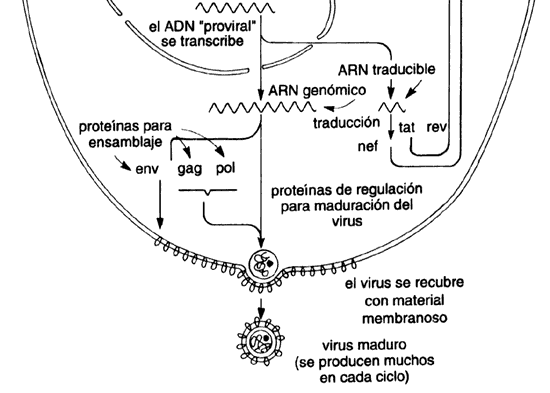
cáncer). Se conoce cómo el virus, después de introducirse a las células, copia
su genoma de ARN lo convierte en ADN, y lo incorpora
o integra al ADN de la célula humana. Desde ahí expresa sus genes,
de lo que resulta la producción de más virus y su salida de la célula, listos
para atacar otras células (véase el recuadro IV.3).
La utilidad de este tipo de conocimientos para lograr el control del SIDA
y de otras enfermedades se ilustrará al abordar la aplicación de la biotecnología
en el campo de la salud, en el capítulo siguiente y, nuevamente, en el VI.
ESTUDIOS SOBRE LAS CAUSAS DEL CÁNCER
Todos sabemos sobre la gran cantidad de recursos materiales y humanos que se han invertido en la incesante búsqueda de la humanidad para conocer las causas, la naturaleza y la forma de curar el cáncer. Aunque estos estudios han ido produciendo conocimientos útiles, la mayoría de lo que sabemos actualmente sobre cáncer proviene de estudios relativamente recientes, que emplean ADN recombinante.
Hay que destacar que cuando nos referimos al cáncer, hablamos de fenómenos patológicos que tienen algo en común: la proliferación descontrolada de cierto grupo de células del organismo, pero quizá no es tan claro para nosotros que hay muchas causas y condicionamientos que pueden desembocar en la aparición del cáncer. Existen factores del ambiente, factores genéticos y hasta infecciosos relacionados con muchos tipos de cáncer. En el fondo, el cáncer es una enfermedad genética. Algo ocurre al programa genético de una célula y ésta no responde más al control normal y se divide continuamente, generando su progenie, la cual hereda la cualidad de dividirse descontroladamente. La lesión debe haber ocurrido en el genoma, y por ello se transmite a las células descendientes. Durante décadas se ha estudiado incesantemente esta enfermedad. Ya en 1911 Peyton Rous describía que el cáncer se podía trasplantar de un pollo enfermo a otro, lo que eventualmente se pudo atribuir a un virus causal.
Como se dijo ya, en el estudio del cáncer, el ADN recombinante ha tenido un impacto generalizado y definitivo. Hemos aprendido más al respecto del cáncer en los últimos 15 años que en todas las décadas anteriores, en las que también hubo intensa investigación. En este proceso de conocimiento ha sido crucial la identificación de genes específicos causantes del cáncer, denominados oncogenes, inicialmente identificados dentro de los virus cancerígenos. Si reflexionamos un momento, lo realmente asombroso es que nuestro organismo tal vez sea un sistema capaz de controlar de manera exquisita la proliferación de gran diversidad de tipos celulares. Evidentemente debe haber genes cuya actividad promueva la proliferación de las células, y que cesan de actuar en un momento determinado, de acuerdo con un programa preestablecido. Quizá uno de los aspectos más interesantes de la investigación del cáncer es que nos ha acercado al conocimiento íntimo de los mecanismos de regulación de la proliferación celular. Resulta ser que muchos oncogenes no son más que versiones alteradas de genes esenciales, naturalmente presentes en todas nuestras células. Estos genes son ahora identificados como protooncogenes.
Hasta el momento existen por lo menos cuatro tipos muy bien definidos de oncogenes:
unos participan en la recepción de señales externas de proliferación, cuyos
productos se localizan en la membrana de las células; otros codifican, en sí
mismos, para señales de proliferación, que son secretadas al medio extracelular;
otros más intervienen en la interpretación o transducción de la señal de proliferación;
finalmente, encontramos genes cuyos productos interaccionan específicamente
con el ADN, causando la expresión de genes que, a su vez, inducen
la proliferación celular.
¿Cómo es que estos genes se transforman de protooncogenes a oncogenes? Es fácil entender los mecanismos, después de que la clonación de los genes y su secuenciación ha revelado las lesiones responsables. (Para explicar algunos de los fenómenos bien caracterizados sobre el cáncer, es importante que el lector tenga presente la descripción un poco más precisa de lo que es un gene en el recuadro IV.1).
Un protooncogene puede ser activado al insertarse un virus cerca de su región
regulatoria.2![]() El gene se expresa ahora de acuerdo con el programa del virus, y no con el programa
natural de la célula. Otra manera de activarse es cuando la lesión resulta de
anormalidades cromosómicas. Nuevamente, los rearreglos tienen como consecuencia
en la asociación del oncogene a nuevas regiones regulatorias.
El gene se expresa ahora de acuerdo con el programa del virus, y no con el programa
natural de la célula. Otra manera de activarse es cuando la lesión resulta de
anormalidades cromosómicas. Nuevamente, los rearreglos tienen como consecuencia
en la asociación del oncogene a nuevas regiones regulatorias.
Por ejemplo, se ha observado la transformación de oncogenes por una sola mutación, o mutación puntual. Puede ocurrir que la alteración de un solo nucleótido y la consecuente alteración de un solo aminoácido, resulte en que la proteína correspondiente muestre una actividad alterada que la saca de control. Ciertos oncogenes resultan de un fenómeno de este tipo. !Ocurre entonces, que en ciertas circunstancias, una alteración en uno de los 3 mil millones de pares de bases del genoma humano resulta en la aparición de cáncer!
Es importante destacar, sin embargo, que aunque se conozcan muchas de las causas moleculares que pueden desembocar en cáncer, y se hayan identificado los genes respectivos, el panorama es, en realidad, mucho más complejo. La interacción de otros genes, los antioncogenes, regula y modera la actividad de los oncogenes. El sistema inmunológico es capaz de destruir específicamente células cancerosas, eliminando quizá gran cantidad de brotes de cáncer que se producen espontáneamente en todos los individuos. Es, pues, por la interacción de todos estos factores, causativos, antagónicos y moduladores, que ocurre el desenlace final de la aparición o no de la enfermedad. Claramente, cualquier factor ambiental o infeccioso que cause mutaciones, puede, en principio, propiciar la aparición de cáncer. La susceptibilidad del individuo, medida por la eficacia de su sistema inmune y la particular actividad de mecanismos de control, determinará la frecuencia con la que las lesiones genéticas producen la enfermedad, o son contrarrestadas.
Entre los descubrimientos más recientes se encuentran ante oncogenes que están muy frecuentemente asociados a ciertos tipos de cáncer. Uno de ellos codifica para la proteína denominada p53, la cual fue seleccionada molécula del año en 1993 por la revista Science. Como su nombre lo indica (una denominación como p53 responde típicamente a una proteína de actividad desconocida, pero de peso molecular de alrededor de 53 mil), esta proteína no se consideró muy importante durante una década después de su descubrimiento. Hoy día sabemos que el gene que codifica para esta proteína se encuentra mutado en hasta el 50% o más de los pacientes con ciertos tipos de cáncer.
Evidentemente, el acervo de conocimientos que se ha ido acumulando permite la apertura de más y más avenidas nuevas para el tratamiento del cáncer (véase el capítulo siguiente).
El reto de entender los fenómenos biológicos a nivel molecular de ninguna manera se circunscribe a las funciones celulares. El reto más formidable es desenterrar los fenómenos integradores que nos permiten interaccionar con nuestro ambiente, percibirlo y transformarlo. La evolución biológica produjo, en un periodo de 3 ó 4 mil millones de años, seres vivos de enorme complejidad, capaces de reaccionar ante el ambiente de maneras verdaderamente complejas. Muchos de los fenómenos que hasta ahora hemos descrito son comunes a todos los seres vivos, pero otros atañen sólo a las formas de vida más complejas. En el ser humano se conjugaron factores biológicos que han dado paso a una nueva y reciente etapa de la evolución: la cultural. En los últimos 50 o 100 mil años, las comunidades humanas se han embarcado en la preservación y transmisión de información, que ha desembocado en una total transformación de sus actividades y de su capacidad para impresionar al resto de la naturaleza. Los sistemas biológicos que subyacen esta portentosa capacidad son objetos de intensa curiosidad para los científicos. No es de extrañarse entonces que, en la actualidad, una proporción significativa de los biólogos experimentales estén dedicados al estudio de los sistemas nervioso y endocrino.
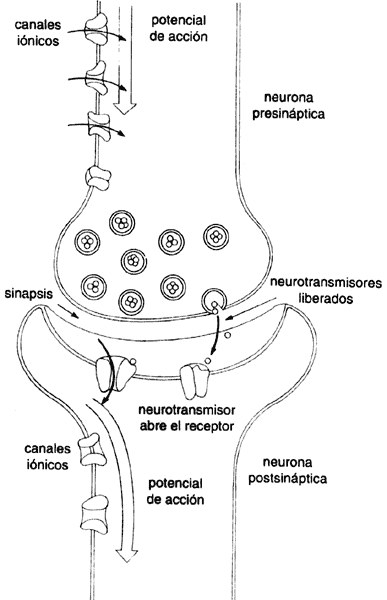
El estudio de la función neuronal a nivel molecular ha revelado la importancia de varios tipos de proteínas. Es importante destacar el papel primordial de los canales iónicos, que participan en gran número de los procesos de todas las células y su comunicación con el exterior. La comunicación del impulso nervioso depende, precisamente, de la existencia de canales que permiten el flujo de iones, es decir, átomos o moléculas con carga electroquímica, en ciertas condiciones. Los canales iónicos se encuentran en las membranas de las células, al igual que otras proteínas que reciben el nombre de receptores. De la interacción de receptores con moléculas mensajeras dependen numerosas transacciones biológicas. En otras palabras, en la superficie de las células se desarrollan una infinidad de interacciones y transacciones moleculares, mediadas por receptores y canales, que dan como resultado la respuesta de las células a los estímulos exteriores. En el estudio del sistema nervioso, los canales y los receptores tienen un papel protagónico (véase la figura IV.1)
En este campo, también adquiere especial relevancia la utilización de sistemas modelo. Sabemos que muchos de los experimentos clásicos de neurobiología se hicieron con gatos, como animales de experimentación. Sin embargo, a nivel molecular; el organismo modelo que ha probado ser más valioso es la mosca de la fruta, Drosophila sp.
Debido a la gran cantidad de estudios genéticos previamente realizados en esta especie, ha sido factible aislar moscas mutantes con deficiencia en alguno de los procesos relacionados con la función nerviosa, y después localizar e identificar los genes responsables de dichas funciones. Quizá nos intrigue cómo es posible obtener moscas mutantes en genes que intervienen en el sistema nervioso. Además, podríamos preguntarnos: ¿que tienen que ver los genes del sistema nervioso de la mosca con los de un ser humano?
La capacidad de aislar mutantes de genes que participan en procesos específicos es una de las herramientas más poderosas de la biología experimental. En el caso de Drosophila podemos ver la aplicación de los métodos más simples e ingeniosos, producto de muchas décadas de tradición en su estudio. Se han aislado genes del sistema nervioso utilizando desde las técnicas más clásicas hasta las más complejas y modernas, producto del conocimiento de biología molecular.
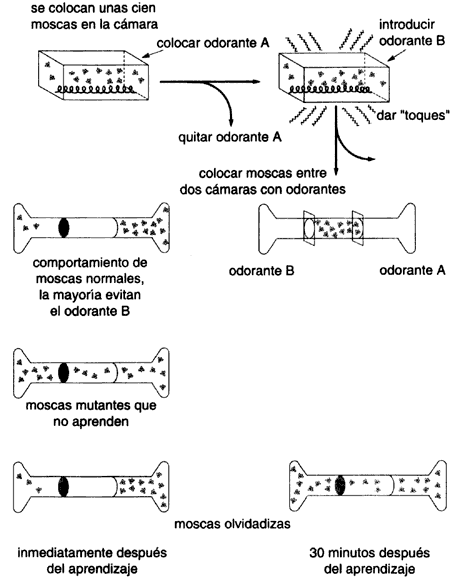
Un ejemplo interesante es el aislamiento de genes relacionados con los procesos
de la memoria. Para este propósito se diseña un experimento muy simple (véase
la figura IV.2). Un conjunto de moscas se introduce en un pequeño recipiente
que contiene una sustancia aromática. Después se retira el aromatizante y se
aplica otra sustancia de aroma diferente, sólo que esta vez acompañada de un
choque eléctrico. Si las moscas han aprendido a asociar el desagradable choque
eléctrico con un aromatizante y no con el otro, entonces deberán alejarse del
primero. Esto es precisamente lo que se observa al colocar a las moscas en una
cámara central conectada a dos cámaras con aromatizante. Las moscas se dirigen
preferentemente a la cámara del olor que no asocian con los choques eléctricos.
Este experimento se puede hacer ahora con moscas previamente sujetas a un tratamiento
mutágeno (por ejemplo, con un tratamiento químico suave de los huevecillos).
Entre las moscas así tratadas puede haber mutantes de muchos tipos, pero algunas
podrían tener alterados los procesos de memoria. Y, en efecto, en el experimento
de las tres cámaras conectadas, se pueden identificar moscas que ya no son capaces
de recordar que un olor se asocia a un choque eléctrico, y se distribuyen por
igual a lo largo de las cámaras. Otras, por ejemplo, se pueden acordar de la
asociación olor-choque eléctrico por un corto periodo, pero luego lo olvidan,
más rápido que las moscas normales. Experimentos parecidos a éstos se han podido
hacer desde hace muchas décadas y, de hecho, una gran colección de miles de
mutantes de todo tipo, está disponible en Drosophila. Lo interesante
es que a partir del ADN recombinante, ahora ya podemos saber qué
genes están alterados y obtener su secuencia.
|
Lo anterior nos conduce a la segunda pregunta: ¿qué relación tienen estos genes mutantes con el sistema nervioso humano? El análisis de los genes mutantes mencionados reveló que las alteraciones se encuentran en proteínas que participan en la señalización celular, muchas de ellas se conservan en todo el mundo eucarionte, desde las levaduras hasta los humanos, como por ejemplo la enzima adenilato ciclasa, cuya alteración es responsable de uno de los fenotipos de pérdida de memoria en Drosophila. En las células nerviosas de mamíferos (por supuesto incluyendo a los seres humanos), también hay adenilato ciclasa. Es altamente probable que la función de la adenilato ciclasa interviene asimismo en los procesos moleculares que subyacen en la memoria de los seres humanos. ¿Quizá a la memoria de corto plazo?
Al observar hallazgos tan interesantes como éstos, suena lógico pensar en cómo seguir avanzando para averiguar más acerca del fenómeno y contrastar hipótesis. Una de las formas más poderosas para experimentar con organismos superiores es reintroducir en ellos genes alterados, de manera muy similar a los genes bacterianos (véase en este capítulo, "Estructura y función del gene").
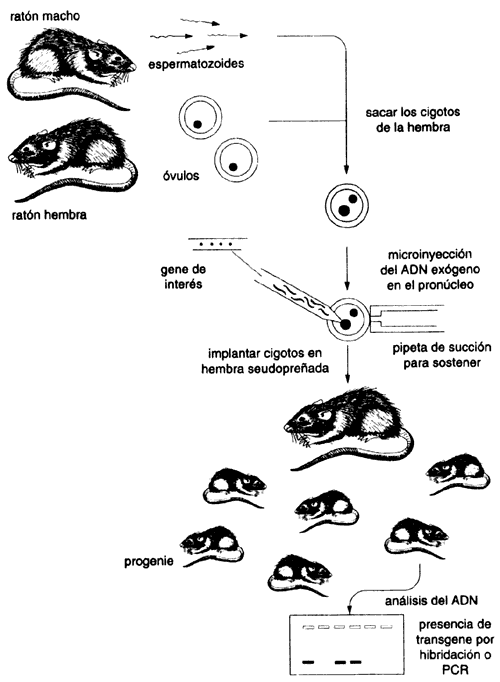
Un organismo al que artificialmente se le han introducido genes exógenos se
conoce como organismo transgénico. La técnica para producir organismos superiores
transgénicos usando ADN recombinante se introdujo a principios
de los años ochenta, inicialmente utilizando ratones en los que la embriología
estaba muy desarrollada (recuadro IV.4). Hoy en día se pueden producir diversos
animales y plantas transgénicas, entre los que se encuentra Drosophila.
Con todos los genes disponibles de esta mosca, y la posibilidad de manipularlos
e introducirlos nuevamente al organismo completo, el panorama para el experimentador
es verdaderamente atractivo. Incluso se dispone de métodos tan ingenuos como
el de la coloración, mencionado anteriormente (véase en este capítulo, "Estructura
y función del gene", p 69). En la mosca de la fruta se dispone del gene rosy,
que es básicamente inocuo, pero produce moscas con ojos rosa, en vez de rojos,
y que permite distinguir una mosca transgénica entre muchas otras que no lo
son, por simple inspección. ¡Podemos pensar ahora en introducir cambios
de diversa naturaleza al gene de la adenilato ciclasa neuronal y observar qué
efectos tienen sobre el aprendizaje de las moscas! Después podemos tratar de
hacer un experimento similar; pero ahora con un ratón.
Otro notable descubrimiento reciente se refiere a los mecanismos moleculares para la detección de los olores. Éste es un fenómeno que ha fascinado a los científicos por mucho tiempo. ¿Cómo podemos distinguir entre los miles o millones de diferentes aromas? En la actualidad los más complejos métodos instrumentales de detección de sustancias palidecen ante la sensibilidad y la capacidad de discernimiento de un sistema biológico.
¡Todavía utilizamos perros para detectar pistas o identificar drogas en las
maletas en los aeropuertos! Un experimento que inició la resolución del dilema
sobre el mecanismo de discriminación de olores fue concebido por Leslie Buck,
una estudiante del postdoctorado que trabajaba en el laboratorio del doctor
Richard Axel, respetado biólogo molecular estadunidense, quienes concibieron
que probablemente el problema de distinguir sustancias químicas muy diversas
podría resolverse con un mecanismo similar al de los anticuerpos (véase en este
capítulo, "Los anticuerpos y cómo se producen"), es decir; a través de un sistema
combinatorio. Pensaron que quizá existiría un conjunto de receptores (proteínas
membranales) parecidos entre sí, pero cada uno capaz de interaccionar con alguna
parte diferente de un odorante. Con algunos cientos de receptores las células
de la nariz podrían distinguir millones de diferentes olores. Esto ocurriría
si la respuesta de la célula depende de la combinación de señales que se genera
de la estimulación de conjuntos diferentes de receptores, diferentes para cada
odorante. A partir de este concepto se dieron a la tarea de identificar en los
ARN mensajeros producidos por las células correspondientes, conjuntos
de moléculas que fueran similares, y pudieran ser la base de este mecanismo
que supusieron podría existir. Con gran satisfacción descubrieron que, en efecto,
las células sensibles al olor expresan un conjunto de receptores muy parecidos
entre sí, y relacionados con la percepción del olor. Aunque todavía hay mucho
trabajo por hacer antes de desentrañar claramente el mecanismo responsable de
este proceso, los descubrimientos descritos muestran cómo el conocimiento previo
(sistema de anticuerpos), la imaginación, y una poderosa metodología nueva,
permiten los avances de la ciencia.
Evidentemente, las descripciones anteriores no abarcan los cientos de genes relacionados con la función neuronal que se han estudiado utilizando ADN recombinante, sólo intentan ilustrar algunas de las maneras como se han llevado a cabo estos estudios.
Conforme nos adentramos en identificar transacciones moleculares que afectan niveles superiores de la experiencia humana, las cosas se tornan muy interesantes. Y muy controvertidas.
Los avances del ADN recombinante están permitiendo proponer hipótesis concretas, verificables, respecto de los condicionantes genéticos de formas complejas de comportamiento. Algunos reportes recientes ilustran estos conceptos.
A partir del análisis de una familia holandesa en la que los varones presentaban
con frecuencia trastornos de conducta (tales como comportamiento violento, incendiarismo,
exhibicionismo y otros), se identificó como probable al gene condicionante:
el que dirige la elaboración de la enzima monoaminooxidasa, de cuya actividad
depende la formación de ciertos neurotransmisores. Un par de años después, se
reportó el resultado de experimentos con ratones transgénicos a los que inactivó
específicamente el gene de monoaminooxidasa.3 ![]() Se observó que los ratones mostraban, entre otras deficiencias, un comportamiento
marcadamente agresivo.
Se observó que los ratones mostraban, entre otras deficiencias, un comportamiento
marcadamente agresivo.
Con una concepción similar, se han hecho recientemente propuestas sobre la posible identificación de genes correlacionados con la homosexualidad. Por el momento, sólo se ha establecido, por mapeo genético, la posible localización gruesa de dicho gene, pero existe la posibilidad de identificarlo plenamente, tal y como se hizo para la fibrosis quística (véase en el capítulo anterior, "Aislamiento de genes responsables de enfermedades hereditarias").
Es importante advertir, dicho lo anterior; que los nuevos conocimientos sólo abren avenidas para enriquecer nuestra concepción de nosotros mismos, inmersos en la evidente complejidad de la naturaleza humana y las relaciones sociales. Es irresponsable hablar del descubrimiento del "gene del alcoholismo", como si estuviera seguro de una relación causa-efecto exclusiva. También sería irresponsable y empobrecedor ignorar la posible influencia de nuestra constitución genética sobre nuestro comportamiento.
El desarrollo embriológico por el cual se genera un organismo completo a partir de un óvulo fecundado es uno de los procesos más fascinantes de la naturaleza. Los estudiosos de la embriología han contemplado extasiados el proceso en el que a partir de células aparentemente idénticas, se van creando estructuras cada vez más intrincadas; cómo se va adquiriendo forma y función.
Nuevamente es sólo hasta la era del ADN recombinante que se ha podido empezar a entender los circuitos moleculares que subyacen en este proceso fundamental. Y nuevamente, el estudio de organismos modelo ha probado ser extraordinariamente importante.
El espacio nos constriñe a mencionar solamente una pequeña muestra de este efervescente campo de la biología experimental moderna, tomando como ejemplo los llamados genes homeóticos, descubiertos en mutantes de Drosophila, en los que, en el desarrollo embrionario se obervaban fenotipos muy llamativos. En uno de ellos, denominado Antennapedia, las estructuras de las antenas son sustituidas por patas. En el bicoid, el embrión se desarrolla con dos telsones o colas, sin tórax ni cabeza (desde luego estas moscas mutantes en genes homeóticos no sobreviven después de ciertos estadios del desarrollo).
Estas mutantes fueron aisladas hace varias décadas, y los genetistas moleculares modernos intuyeron que las lesiones deberían encontrarse en genes de crucial importancia para el desarrollo, en alguno de los interruptores primordiales para la diferenciación. En efecto, estos genes resultaron ser genes maestros, y sus equivalentes o similares luego fueron encontrados en toda la escala filogenética de los eucariontes: levaduras, plantas, animales, etcétera.
Después de una década de experimentación sabemos muchas cosas sobre muchos genes homeóticos. Estos son genes maestros que codifican para proteínas que regulan la actividad del ADN. Las proteínas que codifican se unen específicamente a ciertas secuencias dentro de los cromosomas, activando o inhibiendo la expresión de genes particulares. También se han aislado, en gran parte por los hallazgos de Drosophila, genes homeóticos de organismos superiores como el ratón y el hombre. Continuamente aparecen nuevos genes y nuevos fenotipos causados por éstos.
En uno de los experimentos más llamativos realizados recientemente, se obtuvieron moscas transgenéticas en las que un gene homeótico, que determina etapas fundamentales de la formación de los ojos, se puso bajo el control de señales que activan genes en las patas. El resultado: ¡moscas con ojos perfectamente formados, pero creciendo precisamente en las patas!
Los nombres de algunos de los genes identificados en la mosca de la fruta nos indican el tipo de procesos que se están desentrañando: daughterless (sin hijas), wingless (sin alas), hunchback(jorobada), hairy (peluda). Otros muestran la naturaleza jugetona de los investigadores: sonic que es parte del complejo de genes hedgehog (quizá el lector recuerde al personaje del juego electrónico Sega Genesis).
El estudio de las plantas, desde el punto de vista molecular; ha estado rezagado respecto al de otros organismos. Es quizá una paradoja que las leyes fundamentales de la herencia fueron desarrolladas por Mendel utilizando plantas, y que la genética molecular no se haya inclinado por éstas como uno de sus primeros objetos de estudio. Hay razones que explican esta situación, que veremos más adelante.
El cultivo y manejo de las plantas constituye uno de los factores cruciales del desarrollo de la humanidad. La transición de las culturas de cazadores y recolectores a la vida sedentaria, basada en la agricultura, representa un parteaguas en la evolución cultural. En este sentido, los pueblos han dedicado gran cantidad de su energía e imaginación al mejoramiento y manejo de sus cultivos. La selección de mejores variedades de plantas es una actividad que se ha llevado a cabo por milenios. En efecto, se han empleado las más refinadas técnicas empíricas y la genética clásica para constituir una profesión que revestía gran importancia hacia la mitad de este siglo. La llamada revolución verde, que ha permitido incrementar de manera importante la productividad agrícola, es fruto de la aplicación de estas disciplinas.
Sin embargo, el arribo del ADN recombinante no tuvo en el área de plantas un impacto tan inmediato como el que vimos en el campo de la salud. Quizá la razón principal es que los tiempos de generación de las plantas las han hecho objetos de estudio bastante dificiles. La planta no ha sido un sistema modelo muy útil para resolver preguntas básicas de genética molecular. Por supuesto que esta situación no impidió que se desarrollaran con ellas importantes estudios moleculares, pero no con la intensidad y abundancia que en microorganismos y animales. Esta situación ha cambiado en la última década. Muchos grupos de investigación han detectado la importancia de aplicar las técnicas de ADN recombinante a la biología vegetal, y el gran potencial que tiene la manipulación de las plantas para el bienestar de la humanidad. Una de las áreas más activas y promisorias de la biología moderna es, hoy día, la biología vegetal.
Ingeniería genética de plantas y la tecnología
Describamos primero la explotación de un fenómeno natural que ocurre entre
la bacteria Agrobacterium tumefaciens y ciertas especies vegetales. Agrobacterium
es capaz de invadir la planta e introducirle un segmento de ADN,
llamado plásmido Ti, desarrollando eventualmente un tumor. Esto quiere
decir que una bacteria ha podido hacer plantas transgénicas desde hace millones
de años. En efecto, parte del ADN del plásmido Ti se integra
al genoma de la célula vegetal infectada y lo hereda establemente a las células
hijas. Otro fenómeno importante que facilita la ingeniería genética de plantas
es que éstas tienen gran capacidad de regeneración: muchas plantas pueden regenerar
un organismo completo a partir de un pedacito sacado de una hoja. Con estos
elementos, se desarrolló el sistema básico para la ingeniería genética de plantas
(véase el recuadro IV.5). Uno de los protagonistas del desarrollo de esta tecnología
fue el investigador mexicano, doctor Luis Herrera-Estrella, a principios de
los ochenta, quien se encontraba en aquel momento en Bélgica. Es curioso que
algunas de las contribuciones de importancia internacional de este otro científico
mexicano consistieron en el desarrollo de vehículos de clonación (véase en el
capítulo II, "Concepto de clonación molecular").
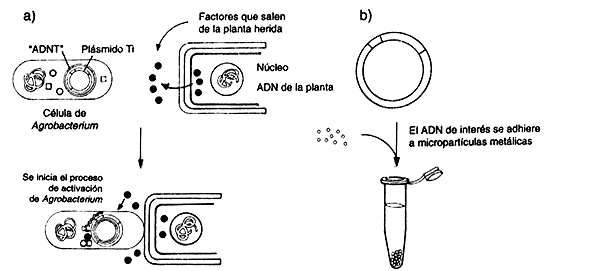
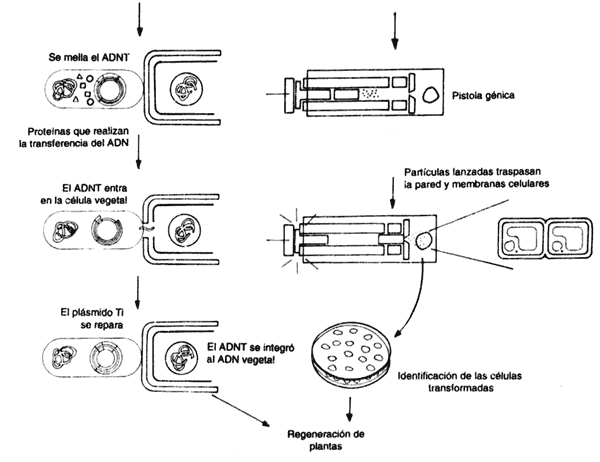
La posibilidad de regenerar plantas con genes exógenos insertados, se ha constituido en un factor crucial para su estudio. Este mecanismo permite acortar significativamente el tiempo de muchas investigaciones que anteriormente requerían experimentos de larga duración, pues dependían de la fertilización convencional de las plantas, y esperar que éstas crecieran y formaran semillas en cuestión de meses. Desafortunadamente, no todas las especies vegetales son susceptibles a la infección por Agrobacterium, por lo que la manipulación de muchas de éstas fue retrasada. En los últimos años, sin embargo, se han establecido mecanismos para transformar y regenerar plantas de muchas nuevas variedades, incluyendo especies de gran importancia agrícola, como los cereales. Una de las maneras como se introducen genes hoy en día, es lo que se ha dado en llamar biobalística. Con esta técnica, el ADN se hace penetrar a las células vegetales atravesando la pared celular; viajando en proyectiles microscópicos que simplemente se disparan hacia los pedacitos de hoja.
Otro de los aspectos más interesantes que se asocian a la biología molecular
vegetal tiene relación con los genes móviles o saltarines. Ya hemos comentado
que la visión clásica consideraba al material genético como un elemento fundamentalmente
estable (véase en este capítulo, "Los anticuerpos y cómo se producen"). Este
material es responsable de la determinación de las características de los nuevos
seres que permanecen de generación en generación. Se pensaba que los procesos
que daban origen a la evolución de las especies eran fenómenos azarosos de cambio
donde se alteraban unas cuantas bases del ADN. En los años cincuenta,
sin embargo, una genetista de plantas, la doctora Barbara McClintock, describió
fenómenos de variación en el maíz y propuso que la causa debería ser el movimiento
o salto de genes, de un lugar a otro del genoma. El trabajo de la doctora McClintock
no fue claramente apreciado hasta que este fenómeno se observó también en la
mosca de la fruta. Finalmente, en 1983, Barbara McClintock fue galardonada con
el Premio Nobel de fisiología o medicina, premiando así su trabajo que conjuntaba
lógica y método impecables, pero también preclara intuición. Los elementos móviles
han sido de gran utilidad para manipular el genoma de algunas variedades de
plantas, y también explican algunos de los fenómenos que en ellas observamos.
Una de las historias más notables que he escuchado se refiere precisamente a esto. Nos la relató la doctora Virginia Walbot, investigadora de la Universidad de Stanford, en una reciente visita al Instituto de Biotecnología de la UNAM.
Entre los objetos de estudio de la doctora Walbot se encuentran los elementos móviles del maíz. En sus investigaciones había descubierto que algunos de estos elementos se mueven de manera excesiva en muchas variedades de maíz, ocasionando que se produzca un elevado número de mutaciones. En su búsqueda del origen de estos elementos, la doctora Walbot descubrió que éste se podía identificar en una variedad de maíz proveniente de la zona zapoteca, del estado de Oaxaca. La variedad llamada zapalote chico contiene estos elementos móviles, pero los puede mantener bajo control. Si se cruza el zapalote chico con otras variedades de maíz, el elemento movil pasa al entorno genético que ya no lo controla, causando su degeneración. Antiguas leyendas que datan de hace muchos siglos amenazaban a los enemigos del pueblo zapoteca con las terribles consecuencias de robarse su maíz sagrado. ¡Los dioses castigarían al ladrón haciendo que sus cosechas se echaran a perder! Hoy día estamos conociendo las causas moleculares de la mitológica maldición.
Así pues, aunque durante milenios, el hombre se ha dedicado a mejorar las variedades de plantas que cultiva, la ingeniería genética ha hecho posible, en poco más de una década, que se abra un nuevo panorama para el conocimiento y la manipulación de este patrimonio fundamental de la humanidad.
EL GRAN PROYECTO PARA SECUENCIAR EL GENOMA HUMANO
Una vez revisados algunos de los logros recientes de la biología experimental, resulta más fácil entender la convicción de los biólogos moleculares de que el ADN recombinante irá permitiendo abordar grandes problemas, que previamente se percibían inaccesibles. Esto ha dado como resultado que muchas de las áreas de investigación que en el presente manifiestan intensa actividad fueran propuestas desde mucho antes: éste es el caso del estudio molecular del genoma humano. Desde inicios de los años ochenta, hubo iniciativas para generar un mapa físico y genético de todo el genoma humano de manera sistemática. La meta de largo plazo seria, obviamente, el conocimiento íntimo de todo el genoma, es decir; conocer toda su secuencia. Pero veamos paso a paso lo que significa esto.
Todos sabemos que nuestro genoma está repartido en 23 pares de cromosomas: éste es el nivel más grueso de mapeo genético. Pero ¿qué podemos decir respecto a qué genes se encuentran en cuáles cromosomas? ¿Qué genes están cerca de cuáles otros? Esta localización relativa de los genes constituye un paso siguiente de finura o resolución del mapa. Finalmente, la descripción de la secuencia de bases de cada uno de los cromosomas, desde su principio hasta su final, es el grado máximo de precisión alcanzable.
Pero ya sabemos que la longitud de cada cromosoma es realmente gigantesca. Llegar a conocer la secuencia completa de siquiera uno de ellos resulta una labor titánica. El más pequeño de los cromosomas humanos, el cromosoma Y está constituido por una hebra de alrededor de 60 millones de bases. Reconociendo lo formidable de la tarea de secuenciar todo el genoma, el progreso del conocimiento en lo que se refiere a secuencias de genes humanos fue desarrollado de manera independiente por grupos de investigación interesados en genes específicos. Esto es, hasta 1986, fue cuando empezó a gestarse una iniciativa para lanzar un gigantesco proyecto internacional que lograra descifrar toda la secuencia de los cromosomas humanos en un plazo razonable.
Como todo gran proyecto, el del genoma humano ha generado buen número de controversias. Muchos pensaron que no tenía sentido lanzar un gran programa para secuenciar algo que de todas maneras no íbamos a saber interpretar de inmediato. Además, sabemos de antemano que el 95% de las secuencias presentes en nuestros cromosomas no codifican para ninguna función directamente y que muchas de ellas no son mas que múltiples repeticiones de lo mismo. Algunos señalaron que era insensato canalizar los fondos, que normalmente se otorgaban a investigaciones bien cimentadas, en las que se persiguen y estudian genes y funciones específicas, para financiar un gran proyecto que pretende estudiar algo que en el 99% de los casos no vamos a saber entender ni aplicar.
Los argumentos anteriores son bastante sólidos y quizá fueron compartidos por gran parte de la comunidad científica. Pero entre los promotores de la idea de secuenciar el genoma humano, se encontraban algunos de los personajes más notables y respetados entre los biólogos moleculares (por ejemplo, James Watson, ni más ni menos). Ellos también tenían argumentos muy interesantes. Para los promotores de la idea, el desvío de los fondos no era una realidad, ya que el Proyecto se iría financiando con fondos nuevos, que en realidad ganaría para sí la ciencia biológica, y se accedería al mismo tipo de status que tenía la física, con sus proyectos internacionales multimillonarios. Según este grupo, el conocimiento que se deriva de estudiar el genoma como un todo, es mucho más atractivo que el que se obtiene de irlo estudiando por partes. Quizá uno de los elementos mas interesantes en favor de la idea es que este gran proyecto internacional promovería de manera definitiva el avance de todas las técnicas relacionadas. De hecho, desde un principio se planteó el proyecto sobre la base de que la eficiencia de las técnicas utilizadas para determinar la secuencia del ADN tendría que incrementarse entre 100 y mil veces. Pero esto se consideraba posible lograrlo en un plazo de menos de 10 años a partir del inicio del proyecto.
Finalmente se impuso el grupo a favor y el Proyecto Genoma Humano ha arrancado desde inicios de esta década. Se han organizado consorcios internacionales en los que participan laboratorios de muchos países. Creo que rápidamente hemos atestiguado la aparición de un fenómeno muy interesante, que es la organización de colaboración a gran escala. De esta Organización empezaron a surgir ideas nuevas sobre cómo distribuir el trabajo, qué requisitos técnicos hay que cubrir antes de empezar a secuenciar en gran escala, etc. En efecto, el gran proyecto internacional ha empezado a generar avances que de otra manera quizá no hubieran ocurrido. Algunos de los aspectos más notables de modo de operar de este gran proyecto internacional ilustran su envergadura y se describen a continuación.
Nuevas técnicas de secuenciación
Los biólogos moleculares se han dedicado con ahínco a secuenciar genes durante
los últimos 15 años. Consideremos, sin embargo, que la gran base de datos acumulada
hasta el momento está constituida por unos 550 mil segmentos de genes y alrededor
de 385 millones de pares de bases en total.4![]() Las cifras anteriores incluyen genes de todos los organismos
para los que se ha obtenido alguna secuencia. El esfuerzo para secuenciar todo
el genoma humano tendría una magnitud unas 10 veces de lo que se ha hecho hasta
la fecha. Claramente, se requiere acelerar y abaratar los métodos de secuenciación.
Las cifras anteriores incluyen genes de todos los organismos
para los que se ha obtenido alguna secuencia. El esfuerzo para secuenciar todo
el genoma humano tendría una magnitud unas 10 veces de lo que se ha hecho hasta
la fecha. Claramente, se requiere acelerar y abaratar los métodos de secuenciación.
Impulsados por los fondos y el entusiasmo generado por el Proyecto Genoma Humano, diversos laboratorios y compañías se han dado a la tarea de desarrollar métodos nuevos que en muchos casos representan gran originalidad y ambición.
En estos momentos, por ejemplo, se están activamente desarrollando sistemas para secuenciar utilizando un chip, mediante técnicas de hibridación de ADN. Estos chips consistirían en arreglos de oligonucleótidos pequeños, dispuestos en filas y columnas casi microscópicas. Se ha podido demostrar que si se hace hibridar un segmento de ADN de secuencia desconocida con uno de estos arreglos, la especificidad de la asociación permite observar un patrón de casillas positivas (donde el ADN desconocido se asoció) y negativas (donde no se asoció), a partir del cual es posible deducir, inequívocamente, la secuencia desconocida. La lectura del patrón puede ser miniaturizada y automantizada, utilizando lectores de fluorescencia láser, conectados a una computadora.
En otro notable desarrollo se plantea utilizar sistemas de electroforesis, pero en una novedosa versión que utiliza tubos capilares (de diámetros microscópicos). Este tipo de electroforesis puede ser completamente automatizada, y los experimentos se realizan en mucho menos tiempo que con la electroforesis convencional. Se están construyendo y desarrollando máquinas que contienen arreglos de decenas de capilares, que se utilizan simultáneamente y, nuevamente por fluorescencia láser, se detectan cantidades mínimas de la muestra y alimentan la computadora.
Existen varios diseños adicionales, muy ingeniosos, cuya finalidad es la misma: abaratar y acelerar la adquisición de datos de secuencia. Es importante considerar que antes de iniciar el Proyecto Genoma Humano, la técnica de secuenciación de ADN tenía un costo de entre uno y dos dólares por cada base. ¡Secuenciar el genoma humano con estas técnicas costaría cerca de 5 mil millones de dólares! Los laboratorios que forman parte del consorcio Genoma Humano ya están secuenciando (con técnicas convencionales) con costos de alrededor 20 cts. de dólar por base, pero todavía queda mucho por avanzar.
Técnicas de mapeo y clasificación
Muy pronto quedó claro para los participantes del gran proyecto que el esfuerzo de secuenciación masivo debía diferirse y que, como prerrequisito, se debía tener un mapa y un conjunto de fragmentos donados que representaran segmentos de los cromosomas identificados en ese mapa. Para lograr este objetivo, se han llevado a sus límites otras técnicas muy interesantes.
En primer término, se dispone ahora de métodos automatizados para separar cromosomas
individuales. Se puede utilizar una máquina llamada FACS (fluorescence
activated cell sorter). Con este equipo se produce un flujo de gotitas microscópicas,
a partir de una preparación que contiene cromosomas previamente extraídos de
células vivas. A través de una sonda de ADN que hibrida específicamente
con un cromosoma determinado (recordemos que hay ya un buen número de genes
identificados para los que se sabe en qué cromosoma se encuentran), y que lleva
una marca fluorescente, se puede lograr que el dispositivo FACS
identifique qué gotitas contienen un cromosoma marcado, induzca una carga electrostática
en esas gotitas y, mediante un campo eléctrico de corta duración, las saque
del flujo principal para recolectarlas en un tubo separado. Esta operación se
puede realizar miles de veces por segundo, logrando así colectar cantidades
importantes de cromosomas puros que fueron seleccionados uno por uno. El siguiente
paso sería someter el ADN de estas preparaciones a alguna enzima
de restricción y clonarlo para obtener una genoteca. El problema que se enfrenta
es que con un vehículo molecular convencional se pueden albergar solamente un
máximo de alrededor de 10 mil pares de bases, así que el más pequeño de los
cromosomas humanos requeriría de más de 6 mil clonas para estar representado
en la genoteca. Se hizo necesario desarrollar métodos más ambiciosos para tener
genotecas más manejables. Uno de los métodos utilizados consiste en generar
cromosomas artificiales de levaduras (YACs, por sus siglas en inglés
yeast artificial chromosome). Los YACs pueden contener cientos
de miles o millones de pares de bases. Así, un cromosoma puede ser dividido
en sólo algunas decenas o cientos de YACs .
La comunidad internacional del proyecto se ha distribuido el estudio de los cromosomas humanos, y cada laboratorio se está dedicando a generar una genoteca de YACs representativa del cromosoma que le fue encomendado. La actividad de los consorcios internacionales dedicados al Proyecto Genoma Humano constituye un ejemplo muy interesante de la labor científica en gran escala. Aquí, por ejemplo, participan diversos especialistas que provienen de disciplinas variadas. La identificación de "marcadores" físicos y genéticos en los cromosomas humanos ha sido una actividad que se llevó a cabo por mucho tiempo. La descripción del significado preciso de estos términos está fuera de los alcances de este libro; pero baste señalar que existen estos antecedentes, basados en técnicas complejas, y que hoy día se complementan con metodología de ADN recombinante. La reacción en cadena de polimerasa (PCR), por ejemplo, se está utilizando activamente para obtener una clasificación reproducible y confiable de pequeños segmentos marcadores presentes en estas colecciones.
En el momento de la impresión de este libro, acicateada por la publicación de la secuencia de los primeros genomas bacterianos completos (véase la siguiente sección), la organización del Proyecto Genoma Humano estaba considerado iniciar la etapa de secuenciación a gran escala. La expectativa es que se pueda completar la empresa a tiempo, antes del objetivo de 10 años en el año 2003.
El lector seguramente observará que la actividad descrita ha ido generando un gigantesco cúmulo de datos que, ya en el momento presente, sólo son manejables con técnicas computacionales.
Imaginemos la tarea que representa descifrar el mensaje de un solo gene. Nos enfrentamos con una larga secuencia de, digamos 1 000 bases. A simple vista sólo vemos una interminable lista de As, Gs, Ts y Cs. Parte de esta secuencia constituye señalamientos para encender y apagar la expresión del gene (véase el recuadro IV.1). El gene estructural contiene el mensaje que determina la secuencia de la proteína codificada. Nos llevaría un buen rato, utilizando el código genético, realizar la simple tarea de escribir la secuencia de aminoácidos de la proteína. Realmente el cerebro humano no es muy eficiente para estos menesteres. Las computadoras son, en cambio, extremadamente eficientes. Una computadora casera haría la traducción en una fracción de segundo.
Afortunadamente, el crecimiento de la eficiencia de las computadoras ha mantenido, y excedido, el paso que lleva la acumulación de datos. El manejo de secuencias biológicas se ha hecho siempre en las computadoras disponibles más potentes. Cuando la base de datos era pequeña (unos cuantos miles de pares de bases) a principios de la década, para su análisis eran suficientes las computadoras de entonces. Hoy día, el análisis de cientos de millones de pares de bases requiere de computadoras muchas veces más poderosas, y que afortunadamente ya existen.
Desde su inicio, por lo tanto, el Proyecto Genoma Humano ha puesto una gran atención en el desarrollo de la herramienta computacional necesaria para el análisis de los datos que se están generando. El consorcio internacional organiza, de hecho, una red de intercambio de datos, por medio de la red Internet, que enlaza millones de computadoras en todo el mundo. Esta gran comunidad esta así conectada continuamente, y puede acceder a los datos generados en todos los demás laboratorios casi simultáneamente a la producción de ellos. Se puede decir, incluso, que el fomento de una cultura de colaboración electrónica internacional en el área biológica, se ha debido en gran parte a la organización y manejo de estas grandes bases de datos.
Secuenciación de otros genomas
De manera simultánea al lanzamiento del Proyecto Genoma Humano, se dio gran impulso a los esfuerzos para secuenciar otros genomas de organismos modelo. Como es de esperarse, estos organismos modelo son los que se han seleccionado para su estudio desde hace varias décadas (véase en el capítulo anterior, Diversos niveles de complejidad"): Escherichia coli (bacteria), Sacharomyces cereviciae (levadura), Caenorhabditis elegans (gusano simple), Drosophila sp. (insecto), Arabidopsis thaliana (planta), Mus musculus (mamífero).
A la fecha de la última revisión de este libro, la comunidad internacional de biología molecular se encontraba celebrando la aparición de la secuencia completa de los genomas de algunos organismos simples: la organización TIGR (The Institute for Genomic Reserach influenzae y Microplasmas genitalium. Se dispondrá también de la secuencia completa del primer eucarionte, Sacharomyces cereviciae (levadura de pan). Claramente, ha surgido un nuevo modo de ver el patrimonio genético, ahora como un todo.
La ventaja que representa el conocimiento de estos genomas es clara. En primer lugar, es posible realizar experimentación, altamente sofisticada, con estos organismos. En segundo lugar, representan modelos más simples de organización, donde los fenómenos pueden resultar más fáciles de entender e interpretar.
Uno de los aspectos más gratificantes del estudio de organismos modelo proviene del hecho, descrito en el capítulo I, de que los organismos se parecen mucho unos a otros en el nivel molecular. Así, el descubrimiento de genes en una bacteria, levadura o insecto, con frecuencia puede decirnos algo sobre genes humanos, ya que es muy frecuente que exista una contraparte humana para el gene en cuestión. Esta reflexión nos da paso al tema de la siguiente sección, en donde se ve el tipo de conocimientos que se pueden derivar de comparar unas secuencias biológicas con otras, sea dentro de una misma especie, o entre especies diferentes.
COMPARACIÓN DE SECUENCIAS: FILOGENIA Y PALEONTOLOGÍA MOLECULARES
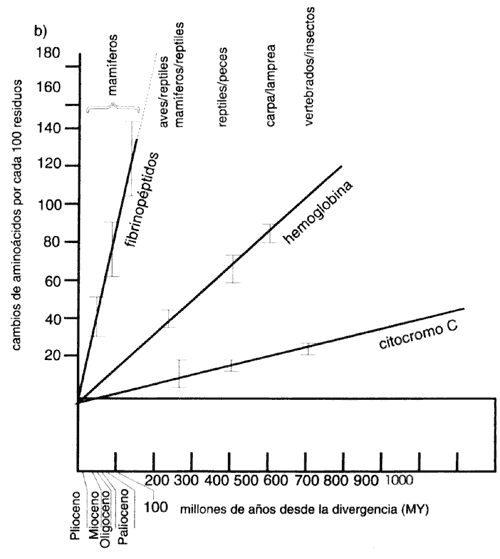
"Nada tiene sentido en biología, si no es a la luz de la evolución", dice Theodosius Dobshansky, el célebre evolucionista. La verdad de esta afirmación es particularmente aparente al comparar las secuencias de las moléculas biológicas. Charles Darwin, fundador de la etapa científica de la teoría de la evolución previó que algún día podríamos establecer todas las relaciones filogenéticas existentes entre los organismos vivientes. La posibilidad de utilizar secuencias de ADN y proteínas nos acerca, como nunca antes, a esta meta.
Quizá sea fácil concordar con la idea de que un perro y un lobo tienen mucho en común. Cuando se afirma que hace algunos millones de años no había lobos ni perros, sino un canino que originó a ambos, ésta parece una aseveración bastante lógica. Sin embargo, si alguien dice que tenemos más en común con una levadura de pan que con las bacterias del yogurt podría intrigar más en qué se basa quien afirme tal cosa. Desde luego que los estudiosos de la evolución han observado, desde hace más de un siglo, mucho más que la apariencia externa y otros caracteres superficiales para establecer relaciones filogenéticas. Pero la aportación de la biología molecular y el ADN recombinante ha revolucionado este campo en los últimos años.
El mismo hecho que posibilita el ADN recombinante sugiere la similitud molecular de los organismos vivos. Todos contienen ADN, proteínas, ribosomas, y se rigen por el mismo código genético. Aunque este hecho era aceptado desde hace varias décadas, la acumulación de datos de la secuencia que surge del empleo de la ingeniería genética le ha dado cuerpo y solidez. Hoy día podemos comparar las secuencias de diversos genes y proteínas, a lo ancho de toda la escala filogenética, y responder preguntas extremadamente interesantes.
El parecido entre un organismo y otro, revelador de un origen común, se manifiesta
en las secuencias de sus genes. Por ejemplo, entre una persona y otra, más del
99.9% de las secuencias son iguales. Entre un humano y un chimpancé, alrededor
de 99% de la secuencia es igual. (Tomemos en cuenta, sin embargo, que una diferencia
de 1%, en un genoma de 3 mil millones de bases, constituye todavía una cifra
elevada: 30 millones de bases distintas. No sabemos cuántas o cuáles de las
diferencias, a nivel del genoma, son importantes para dar origen a las diferencias
observables entre una especie y otra.) Estas similitudes y diferencias se pueden
analizar por técnicas que evalúan el parecido global del ADN, mediante
técnicas de velocidad de reasociación entre un ADN y otro, pero
se pueden analizar a un nivel más fino, comparando las secuencias de genes individuales.
Hay genes (y sus correspondientes proteínas) que difieren más que otros cuando
se comparan dos especies. Algunas de las proteínas componentes de las células
requieren conservar su estructura y secuencia de manera precisa, mientras que
otras pueden mantener su función útil aunque su secuencia varíe considerablemente.
Lo interesante de esta situación es que podemos utilizar ciertos genes para
comparar especies muy cercanas (o poblaciones dentro de una misma especie) y
otros para comparar especies muy lejanas (véase el recuadro IV.6). Por medio
de la comparación de secuencias de genes se han realizado hallazgos que empiezan
a revolucionar nuestra concepción de las relaciones entre los seres vivientes.
Tal como lo afirma Russell Doolittle, reconocido analista de secuencias moleculares,
quien se considera un paleontólogo, pero no usa palas ni desentierra huesos;
sus herramientas son una computadora y los datos en secuencia que se generan
en cientos de laboratorios alrededor del mundo.

Para comparar poblaciones humanas, se pueden usar secuencias de las mitocondrias
(que son organelos celulares que se encargan de transformar energía). Estos
organelos contienen ADN probablemente de una simbiosis en la que
una bacteria colonizó el interior de una célula eucarionte. Las mitocondrias
se heredan fundamentalmente de la madre, dado que los espermatozoides contribuyen
con una ínfima proporción de las mitocondrias al fecundar el óvulo. Esto hace
que sea más fácil establecer relaciones entre grupos cercanos utilizando secuencias
de ADN mitocondrial.
En un estudio reciente, cuyas conclusiones son aún controvertidas, se utilizaron
secuencias de ADN mitocondrial para establecer que todas las poblaciones
humanas provienen de una hembra que vivió en África hace unos 200 mil
años. Esta interpretación ha establecido un serio cuestionamiento sobre los
conceptos clásicos de la paleontología humana. Es decir, la afirmación de que
todos los humanos que vivimos actualmente descendemos de una hembra en particular
(por lo que se refiere el estudio a Eva), realmente debe interpretarse como
que se analizó el linaje femenino, que se observa en las mitocondrias, pero
desde luego en todo el proceso, y desde incontables generaciones atrás, los
humanos descienden de machos y hembras reproduciéndose de manera normal.
Por otra parte, lo que el estudio realmente puede indicar, es que todos los humanos descienden de una población africana que vivió hace unos 200 mil años.
La controversia que este tipo de estudios suscita ilustra un aspecto muy interesante del quehacer científico. En este caso, los paleontólogos tradicionales cuentan con un importante cúmulo de datos, proporcionados por el registro fósil, y toda una serie de teorías e interpretaciones de estos datos. La revolución establecida por la disponibilidad de secuencias de moléculas biológicas resulta de que algunas de las teorías en boga se ven reforzadas, mientras que otras se ponen en entredicho. Los científicos, al igual que todos los seres humanos, establecen lazos afectivos con sus creaciones (por ejemplo, sus teorías), y se apegan a los métodos de análisis que les son familiares. Seguramente, la irrupción de los biólogos moleculares en la escena de la evolución natural y, en particular, la humana, continuará suscitando controversia. A la larga, será de la integración de las diversas disciplinas de donde obtendremos las respuestas más satisfactorias a nuestra curiosidad sobre el origen del hombre.
Refinamiento de la clasificación de organismos
El uso de ADN mitocondrial permite la comparación de poblaciones
humanas, usando secuencias de genes que han cambiado en forma extraordinariamente
lenta se puede atizbar en las relaciones entre organismos mucho más distantes.
Los genes que codifican para el ARN, que forma parte de los ribosomas
(véase el capítulo I, el recuadro I.2), se parecen tanto que podemos usarlos
para comparar grupos tan lejanos como plantas con animales, o bacterias y hongos,
por ejemplo.
Los datos generados por la comparación de secuencias de ARN ribosomal indican que la clasificación clásica de los reinos (plantas, animales, hongos, monera y proroctista), no corresponde a las ramas más primigenias. En su lugar se sugiere introducir una nueva clasificación, usando un nuevo concepto de tres dominios: bacteria, arquea y eucaria. De manera similar, mediante la comparación de secuencias de genes, cuya divergencia es intermedia, se está revisando la clasificación a todos los niveles, estableciendo relaciones más seguras entre diversas plantas, animales, etcétera.
Recuperación de ADN fósil
mediante la reacción en cadena de polimerasa (pcr)
Como ya mencionamos en el capítulo II, la reacción en cadena de polimerasa
permite obtener grandes cantidades de ADN a partir de cantidades
minúsculas, incluso de una sola molécula. Así pues, basta el material genético
de unas cuantas células para poder estudiarlo. Esta potente herramienta está
siendo utilizada en todos los ámbitos del ADN recombinante, en
sus facetas básica y aplicada, a menos de 10 años de su creación.
Una de las aplicaciones más sorprendentes del método de la PCR lo constituye la recuperación y secuenciación de segmentos de genes de muestras muy antiguas. Varios investigadores pensaron que era concebible que material biológico preservado por diferentes procesos de fosilización contuviera aún cantidades detectables de ADN. Así pues, se dieron a la tarea de tratar de aislar algunos de estos segmentos de material genético usando la PCR. Para sorpresa de muchos, efectivamente ha sido posible amplificar y secuenciar diversos segmentos de ADN a partir de muestras de increíble antigüedad. El proceso de fosilización por ámbar ha sido especialmente benigno para la preservación del material biológico. El ámbar es un material resinoso, producto secretado por los árboles, que en ocasiones atrapa materiales biológicos como hojas o pequeños insectos. Algunas de las muestras preservadas en ámbar tienen varios millones de años de antigüedad y muestran con toda claridad los caracteres morfológicos de los seres que ahí quedaron atrapados. Entre los ejemplos más notables de muestras estudiadas hasta la fecha se encuentran los restos de una planta de unos 40 millones de años de antigüedad, y de un gorgojo que cuenta con alrededor de 120 millones de años.
Es importante hacer notar que todavía existe controversia acerca de la validez
de este tipo de estudios. Es de esperarse que el ADN en muestras
de millones de años de antiguedad se encuentre severamente dañado y fragmentado
por procesos químicos espontáneos. En opinión de algunos, aun la increíble preservación
del aire y el agua que el ámbar parece ofrecer no sería suficiente para mantener
el ADN en un estado tal para ser estudiado. Asimismo, se acepta
unánimemente que la recuperación de ADN dé muestras, aun
de las relativamente reciente (por ejemplo, momias o animales extintos hace
unos pocos miles de años), será siempre fragmentaria, incompleta.
Esta capacidad parcial de atisbar en el pasado de la evolución biológica promete una gama muy interesante de estudios que indudablemente contribuirán a nuestros conocimientos. Ya sabemos, por ejemplo, que los mamuts eran parientes más cercanos del elefante africano que del asiático. O que el tigre dientes de sable pertenecía a la familia de los grandes felinos, muy cercano a leones y tigres, pero más alejado de los pumas.
Las posibilidades de esta tecnología también ha incitado a la imaginación. La idea central de la novela Parque Jurásico, del escritor Michael Crichton, se basa, precisamente, en esta capacidad real de recuperar ADN muy antiguo. Evidentemente, la distancia que existe entre aislar y secuenciar algún pequeño fragmento de ADN prehistórico y la de reconstruir el genoma completo de un organismo es abismal. Un obstáculo quizá todavía más importante lo constituye el hecho de que el ADN, por sí mismo, no es capaz de realizar función biológica alguna. Requiere estar empaquetado en cromosomas y con toda la maquinaria celular a su disposición para poder expresarse. Durante miles de millones de años una célula sólo ha provenido de otra célula, llevando consigo, no sólo el ADN, sino también una buena parte del resto de los componentes celulares. Creo que no es difícil predecir que la reconstrucción total de la más simple de las células (por ejemplo, la de una bacteria), a partir de sustancias químicas aisladas, es una empresa de tan gigantescas proporciones que habrán de pasar muchas décadas antes de que pueda lograrse.
REFLEXIÓN SOBRE EL IMPACTO DEL CONOCIMIENTO BIOLÓGICO
En el capítulo que aquí concluye, se presenta una muestra mínima de la avalancha
de conocimientos generados a partir del ADN recombinante. Si nos
detenermos un minuto a reflexionar, las mujeres y los hombres que vivimos hoy
somos los primeros que tenemos acceso a un conocimiento más o menos coherente
de nuestra naturaleza biológica. La fechas que marcan la formulación y demostración
de conceptos verdaderamente fundamentales son todas increíblemente recientes!
Las decenas de miles de generaciones de Homo sapiens que nos precedieron, han centrado sus reflexiones en las facetas trascendentes (religiones, sistemas filosóficos) y sociales de la naturaleza humana. Dos conceptos inherentes a nuestra naturaleza psicológica y biológica fueron (por necesidad) de índole tentativa o especulativa. Sólo ahora contamos con los elementos para integrar una nueva concepción del cosmos y de nuestra naturaleza. Es éste un gran reto para nuestra generación y las que nos sucederán.