VI. CÓMO SE COMUNICAN LAS CÉLULAS
Es fácil pensar en la necesidad de comunicación entre las células de distintos tejidos, órganos o sistemas de nuestro organismo; se requieren estructuras encargadas de regular las funciones de muchos órganos y tejidos. No nos cuesta trabajo imaginar, por ejemplo, la función del sistema nervioso central, como una especie de sistema de gobierno que centralice e influya o maneje las funciones de muchos otros órganos. Sin embargo, nuestro organismo representa uno de los estadios más evolucionados que existen en la naturaleza, pero este grado de complicación enorme debe de haber surgido originalmente de la asociación de organismos unicelulares que poco a poco fueron desarrollando la capacidad de asociarse y de comunicarse.
Algunos estudios realizados nos muestran que ni siquiera es necesario que exista
la formación de organismos pluricelulares en el sentido estricto de la palabra
para que haya comunicación entre distintas células. Existe un microorganismo
llamado Dictyostelium discoideum que es unicelular, pero en ciertas condiciones
distintas células que representan realmente organismos diferentes, por alguna
razón se reúnen y forman grupos o cúmulos de células que empiezan a funcionar
como una especie de colonia (Figura 39). Todo parece indicar que fuera de la
comunicación inicial, que parece tener como finalidad la agrupación de las células,
una vez que se ha formado esta especie de colonia no hay ninguna otra manifestación
clara de intercomunicación. Es claro en este caso que, ante condiciones adversas,
la asociación de grandes números de células les ofrece mayores posibilidades
de sobrevivencia; simplemente, cuando se agota el alimento, algunas de las células
pueden proceder a formar esporas, formas de vida latente, resistente a la falta
de alimentos y otros materiales necesarios para su desarrollo y crecimiento.
La formación de esporas por parte de algunas de las células se lleva a cabo
gracias a que otros miembros de la colonia se sacrifican a su vez para proporcionar
los materiales que aseguren al final la sobrevivencia de la especie.

Figura 39. El comportamiento "social" de Dictyostelium discoideum.
Al agotarse el alimento, los individuos se asocian en grupos. Esto les permite
formar un esporangio y esporas que aseguren su sobrevivencia.
Todo el mecanismo tiene como punto de partida un sistema de comunicación según
el cual una célula es capaz de segregar una sustancia al medio, y es así que
atrae a otras y da como resultado que luego se mantengan juntas. Este mecanismo,
que nos puede parecer extremadamente sencillo, representa sin embargo una serie
larga de pasos que podríamos imaginar al menos de la manera siguiente (Figura
40):
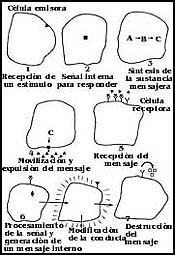
Figura 40. Generación de un mensaje y recepción en la comuncación celular. La célula emisora también obedece a algún estímulo o condición del medio para emitir una señal.
1) Las células deben contar con sistemas de detección para responder a ciertas condiciones con el envío de la señal de comunicación.
2) Estos sistemas de detección luego deben dar una señal interna para que se sintetice o produzca la molécula mensajera que se debe enviar a otras células.
3) Se inicia la síntesis de la sustancia en el interior de la célula, gracias a la existencia de enzimas y materiales adecuados.
4) La sustancia debe movilizarse hacia la membrana y luego expulsarse al exterior por un sistema especial.
5) La célula receptora debe tener moléculas en su superficie para detectar la presencia de esta sustancia o "mensaje".
6) Una vez recibido el mensaje, la célula receptora debe contar con un complejo sistema para interpretarlo primero y luego para modificar su conducta.
7) Finalmente, una vez captado el mensaje o señal, la célula receptora debe tener capacidad para destruirlo y evitar que continúe modificando indefinidamente su conducta.
En el caso del Dictyostelium, se ha llegado a aislar la molécula que
es relativamente sencilla, y recibe el nombre de AMP cíclico (CAMP),
que se produce a partir del ATP, que ya conocemos, gracias a una
enzima a la que se le ha llamado ciclasa, y se degrada gracias a otra, llamada
fosfodiesterasa (Figura 41). Ha sido posible provocar la respuesta de células
aisladas de Dictyostelium agregando una pequeñísima gota de esta sustancia
con un capilar muy fino. Se observa que una célula sometida a este estímulo
inicia luego un movimiento amiboideo hacia donde se encuentra la sustancia.
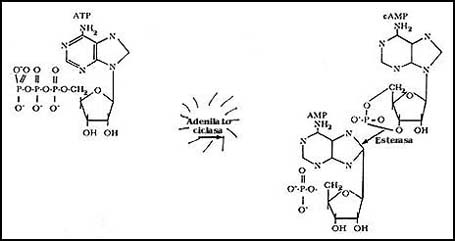
Figura 41. El AMP cíclico, su producción y su degradación.
Hay una enzima regulada por las señales externas que produce esta sustancia.
Una vez cumplida su misión como mensajero interno o segundo mensajero, el AMP
cíclico es destruido por una enzima especial.
El ejemplo de la señal química que da lugar al desplazamiento de células de Dictyostelium es probablemente sólo un caso sencillo y primitivo de intercomunicación celular, en la cual una célula modifica el comportamiento de otra igual, emitiendo o segregando una sustancia al medio en donde se encuentran ambas. Es claro que éste es un sistema que requiere, por una parte, de la participación de la membrana de la célula que emite la señal, pues el fenómeno de secreción es esencialmente membranal, y de otro fenómeno, también membranal, que consiste en la recepción y procesamiento de la señal recibida, para que posteriormente ésta se convierta en una modificación del comportamiento de la célula receptora.
LA COMUNICACIÓN ENTRE CÉLULAS VECINAS
Hay células que, no obstante encontrarse juntas en un órgano protegido, están aisladas por su membrana y no se comunican entre sí más de lo que lo harían células distantes. Hay órganos en los cuales las células se encuentran bien separadas por sus membranas, y fuera del intercambio que puede haber de ciertas sustancias, no puede hablarse estrictamente de un sistema de comunicación entre ellas. Este es el caso de muchos órganos en los cuales las células se encuentran agrupadas constituyéndolos, pero no tienen mecanismos de comunicación ni mucho menos sistemas como el antes mencionado, en el que la producción de una sustancia por parte de una de ellas sea capaz o esté diseñada para modificar el comportamiento de la otra.
En el otro extremo hay células que forman lo que se llama un sincicio, en las
cuales no hay realmente una membrana entre ellas, y puede hablarse más bien
de células multinucleadas, que de diferentes células. Esto se observa en algunos
epitelios o en las prolongaciones de algunos hongos. Como en el caso de células
que prácticamente comparten el mismo medio interno y están en todo caso sujetas
todas a las modificaciones o señales que les lleguen del medio externo, pero
estas células no forman ni requieren, mucho menos, de señales para comunicarse
unas con otras, dado que el citoplasma de una es el citoplasma de la otra (Fig.
42).
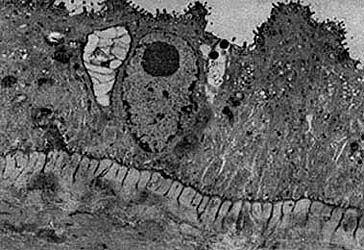
Figura 42. Micrografía de un sincicio. Cortesía del Dr. A. Cárabez, Instituto de Fisiología Celular, UNAM.
Hay tal vez un caso intermedio en el cual algunas células se comunican entre sí por medio de orificios o canales, que reciben el nombre de uniones permeables, y que permiten el paso de materiales de tamaño pequeño, con pesos moleculares hasta de 1 000 o 1 500 daltones (un dalton es la unidad de masa en las moléculas; el átomo de hidrógeno tiene una masa de un dalton). Sin embargo, estas comunicaciones parecen representar más bien mecanismos para intercambiar materiales y equilibrar sus concentraciones tratándose de sustancias que son comunes a todas las células. El sistema parece representar un mecanismo para que el funcionamiento de las células sea más uniforme habiendo una conexión física entre sus materiales.
Este tipo de relaciones entre las células de ninguna manera representa sistemas especializados de comunicación, concebida como el envío de señales de unas a otras, sino simplemente diversos mecanismos para intercambiar materiales, muchos de los cuales pudieran considerarse como nutritivos. Realmente puede hablarse de intercomunicación celular en los casos en que una célula es capaz de enviar una señal y modificar el comportamiento o la función de otra más o menos lejana. Existe inclusive el caso de células con un elevadísimo grado de especialización en este sentido, que son las células nerviosas, en las cuales no hay lejanía estrictamente, pues las células establecen contacto unas con otras a través de prolongaciones muy largas, que se llaman axones. Sin embargo, estas células sí pueden llevar señales a través de distancias que pudiéramos considerar enormes, no obstante que se encuentran en contacto a través de esas prolongaciones, tomando en cuenta el tamaño mismo de ellas. Puede haber axones extremadamente largos, y las señales son capaces de viajar distancias hasta de muchos centímetros, que son enormes si se piensa en las dimensiones de las células mismas.
Sin embargo, en ambos casos, ya sean células lejanas que se ponen en contacto a través de prolongaciones que pueden ser muy largas, o células entre las cuales median distancias de miles o millones de veces sus propias dimensiones, el elemento común de comunicación consiste en que una de ellas emite una sustancia al medio en que se encuentra. Ésta, al llegar a la otra célula, encuentra un receptor que es capaz de reconocerla entre muchísimas otras. Finalmente, al unirse la molécula que pudiéramos llamar mensajera con su receptor, la unión de lugar a una modificación en el comportamiento o función de la célula receptora. Las modificaciones en el funcionamiento de las células receptoras pueden ser extremadamente diversas; por ejemplo, las moléculas de epinefrina producidas en las glándulas suprarrenales que se encuentran localizadas sobre ambos riñones de los animales o los humanos, al llegar al hígado dan como resultado la liberación de una cantidad importante del azúcar glucosa a la sangre del individuo mediante una serie de procesos más o menos complicados. Por otra parte, la terminación nerviosa que viene desde el sistema nervioso central con una serie de relevos, y que llega a una fibra muscular, al liberar en una zona especial de ésta unas cuantas moléculas de la sustancia llamada acetilcolina, da lugar a que se contraiga. Estos son sólo dos casos de las modificaciones que podemos observar del comportamiento de una célula mediante una señal que le envía otra.
En el caso de las células lejanas, que no se encuentran unidas a través de sus prolongaciones, las señales que envían unas a otras están representadas por una serie grande de sustancias químicas relativamente sencillas, que llamamos hormonas. Éstas pueden ser consideradas como los mensajes que unas células envían a otras con objeto de que las segundas desempeñen o modifiquen cierto tipo de funciones.
CUADRO 1. Algunos tipos de hormonas.
| Hormona |
Producción |
Células "blanco" |
efectos |
| Insulina |
Páncreas |
Casi todos los tejidos |
Favorece la utilización delos azúcares,
estimula la síntesis de proteína |
| Epinefrina |
Cápsulas suprarrenales |
Hígado y músculo |
Hidrólisis del glucógeno. Liberación
de glucosa a la sangre |
| Tiroxina |
Tiroides |
Prácticamente todas |
Acelera el metabolismo |
| Pratohormona |
Paratiroides |
Hueso, riñón, intestino |
Regula el metabolismo del calcio y el fosfato |
| Testosterona |
Testículo |
Órganos sexuales |
Favorece la maduración y el funcionamiento |
Las hormonas se producen dentro de las células que las van a segregar al medio; en la mayor parte de los casos son almacenadas dentro de pequeñas vesículas de membrana que provienen a su vez de las estructuras membranosas del interior de la célula. Es claro que en un gran porcentaje de los casos, las hormonas y otras sustancias que sirven como señales de comunicación entre las células son liberadas por las células que las producen, mediante un mecanismo que se presenta en forma esquemática en la Figura 47. Las vesículas que contienen a las hormonas se encuentran cerca de la superficie de las células y al recibirse la señal de liberación se funden con la membrana celular; esta fusión da lugar a la exteriorización del contenido que en el caso de los animales superiores pasa al medio externo y de ahí a la circulación sanguínea, la cual eficiente y rápidamente las hace llegar a todo el organismo.
Es necesario señalar que muchas de estas hormonas son solubles en agua, y sólo unas cuantas son liposolubles. Parece haber una gran diferencia entre ambos tipos de sustancias; debido a su característica liposoluble al llegar a las células son capaces de atravesar la membrana celular y suelen contar con receptores en el interior, a través de los cuales producen diversos cambios en el comportamiento o el metabolismo.
Las hormonas que son solubles en agua son segregadas a la circulación y llegan prácticamente a todas las células del organismo. Para estas señales hay un sistema de recepción, es decir, hay moléculas en las membranas de las células receptoras que son capaces, primero, de reconocer la señal que les llega y distinguirla claramente de muchas otras emitidas por otras glándulas del organismo y que van dirigidas a distintas células. Pero esto no es todo. Las células que cuentan con receptores para esta señal, deben también tener un sistema de procesamiento, una serie de pasos posteriores a la recepción que haga posible que la señal produzca efectivamente una modificación del comportamiento de la célula. Así pues, hay dos elementos fundamentales en este sistema: la recepción misma de la señal y su reconocimiento entre otro gran número de señales y luego el procesamiento de la misma, para que se convierta en una acción definida por parte de la célula.
La recepción de la señal. Las células de un organismo que son sensibles a una determinada hormona o sustancia segregada al medio interno por otra célula cuentan en su superficie con moléculas proteicas, que son las encargadas de reconocer las señales con gran especificidad y sensibilidad. La especificidad se refiere a la capacidad que tienen estas moléculas receptoras de distinguir a una hormona entre un gran número de ellas que puedan encontrarse en la circulación; en el caso del organismo humano existen alrededor de unas 20 hormonas que se encuentran constantemente en circulación y en concentraciones pequeñas; los receptores de las células sensibles deben distinguir entre todas ellas de modo que no haya interferencia de una señal con otra, como debe ser el caso para un individuo normal de cualquier especie, si se pretende que su funcionamiento sea más o menos armónico. La situación es comparable a la de un receptor de radio que debe distinguir en un momento, cientos o tal vez miles de señales que hay en el ambiente.
La sensibilidad de los sistemas de recepción de las señales hormonales, es decir, de los receptores que se encargan de establecer contactos con las señales que llegan de otras partes del organismo debe ser enorme, pues es necesario tomar en cuenta el tamaño de las glándulas que las producen; la hipófisis por ejemplo pesa alrededor de un gramo; ambas glándulas suprarrenales aproximadamente cinco gramos; la glándula tiroides alrededor de uno o dos gramos; las glándulas paratiroides no llegan a medio gramo entre todas. Es obvio entonces que la cantidad que se produce de cada hormona es pequeña. Pero la señal emitida por una glándula, además de ser pequeña, se distribuye a través de la circulación por todo el organismo. Esto da lugar a una concentración extraordinariamente baja de las hormonas en la sangre y en el medio que rodea a las células en las cuales va a actuar. Nos puede dar una idea de la sensibilidad de los receptores de nuestras glándulas, la comparación entre la sensibilidad de éstos y la de nuestras papilas gustativas, por ejemplo. El sabor más o menos habitual de una taza de té o de café endulzada, se logra con una concentración 0.2 molar de azúcar en estas bebidas. El sabor salado del agua de mar se debe a una concentración molar de aproximadamente 0.7 de cloruro de sodio en ella. Una célula muscular es capaz de responder a concentraciones molares de 0.000 000 1 M de insulina. En otras palabras, esto quiere decir que los receptores de una célula muscular para la insulina, son aproximadamente un millón de veces más sensibles a la insulina que lo que nuestras papilas gustativas lo son al azúcar o la sal. Por otro lado, es posible imaginar también la concentración de las hormonas en el líquido que rodea a las células, y encontramos que ésta no es tan exageradamente pequeña; 10-7 M de insulina implica la presencia de alrededor de 60 000 millones de moléculas en un microlitro, que es a su vez la milésima parte de un centímetro cúbico o bien la millonésima parte de un litro. Habría entonces 60,000,000,000,000 (sesenta billones de moléculas en un centímetro cúbico). De cualquier forma, dentro de los estándares biológicos se considera que la sensibilidad de los receptores que se encuentran en las células de las membranas sensibles a las hormonas es extraordinariamente alta.
De las características anteriores salta a la vista la semejanza que hay entre los receptores y los sistemas de transporte de las células. De inmediato se antoja que las células y los organismos desarrollaron sus sistemas de recepción de señales aprovechando sistemas de transporte que ya existían, y simplemente los modificaron para que funcionaran, no para introducir sustancias a las células, sino simplemente para reconocerlas y luego para responder a su presencia con el procesamiento de la señal recibida (como veremos a continuación).
El procesamiento de las señales. La interacción de las hormonas con sus receptores representa lo que podríamos considerar como la culminación del proceso de comunicación. La hormona o señal enviada por otra célula llega y se une con su receptor que ha sido específicamente diseñado para reconocerlo. Pero una vez que la señal ha sido recibida, es necesario que la recepción produzca a su vez una modificación del estado funcional de la célula que la ha recibido. Podemos volver al ejemplo de la molécula de epinefrina que puede llegar a una célula hepática y dar lugar, en última instancia, a que esta célula libere glucosa al medio en que se encuentra; la liberación de glucosa por parte de la célula se hace a partir del almacenamiento de esta sustancia en forma de polímero, que se llama glucógeno. En otras palabras, el glucógeno es una sustancia formada por la unión de millones de moléculas de glucosa, y para que la célula libere glucosa al exterior, éste debe fraccionarse y dar lugar a las unidades que lo componen.
En términos muy simples, en la Figura 43 se muestra el proceso mediante el
cual funciona la hormona que nos ocupa. La epinefrina interactúa con su receptor
que se encuentra en la membrana celular. Luego hay un segundo paso que consiste
en la transformación inicial de la señal en otra, que también ocurre dentro
de la membrana. Una vez que la hormona se ha unido a su receptor, éste interactúa
con una molécula de proteína llamada proteína G, que a su vez, con la participación
de otro elemento más (el GMP), que se muestra en la figura, es
capaz de interactuar con una tercera molécula de proteína, que no es otra cosa
que una enzima. Esta es capaz de convertir al ATP, molécula que
ya conocemos, en AMP cíclico, otra molécula que fue presentada
al principio de este capítulo. En realidad, el proceso de recepción de la señal
y transmisión de una orden al interior de la célula consiste en forma completa
en la interacción de la hormona con su receptor que se traduce, se transforma
o se convierte en la producción de un cierto número de moléculas de AMP
cíclico en el interior de la célula.
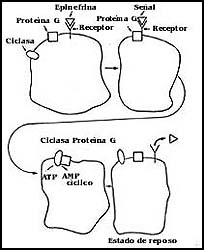
Figura 43. Esquema de la acción de una hormona, la epinefrina, para desencadenar
la producción de AMP cíclico. En la acción interviene, además del
receptor, una proteína llamada G, que actúa como intermediaria entre éste y
la cidasa, y permite después le desactivación del sistema.
Finalmente, el AMP cíclico que se genera en el interior de la célula es realmente lo que se ha dado en llamar el segundo mensajero, que sirve como elemento de transición o de comunicación entre la señal que se entrega a nivel de la superficie celular y los sistemas enzimáticos o metabólicos del interior de la célula.
Una vez que se ha liberado el AMP cíclico, éste puede interactuar
con otras enzimas del interior de la célula, que en forma encadenada dan finalmente
lugar a la ruptura del glucógeno y a la liberación de glucosa al exterior (Figura
44).
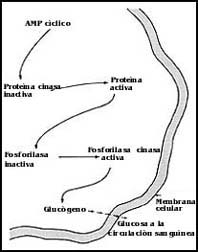
Figura 44. Efectos del AMP cíclico sobre una célula del hígado
para que libere glucosa a la sangre. La señal original fue una descarga de epinefrina.
Este ejemplo que hemos tomado sólo representa uno de los numerosos casos mediante los cuales una hormona producida en una determinada glándula da lugar finalmente a la respuesta de una célula que se encuentra, en términos de lo que son las dimensiones relativas de las células, a una gran distancia de aquella que envió la orden o la señal.
Un elemento extraordinariamente importante que debe tomarse en cuenta en el procesamiento y realización de la orden representada por una hormona que llega a una célula sensible es el hecho de que, para que la orden sea efectiva y útil al organismo, un pequeño número de moléculas de hormona que llegue a una célula debe dar lugar a que ésta, en el caso que nos ocupa, por ejemplo, libere al medio que la rodea miles o millones de moléculas de glucosa. El caso es el siguiente: la glándula suprarrenal libera en una descarga unos cuantos microgramos de epinefrina que se diluyen en todo el torrente sanguíneo. Una muy pequeña parte de estas moléculas, unos cuantos nanogramos, interactúa con los receptores de las células hepáticas y permite la producción de varios gramos de glucosa. Un gramo es un millón de microgramos; éste es entonces, en forma muy aproximada, el factor de amplificación. Pero para esto es necesario contar con un sistema que amplifique la señal. Es fácil imaginar la ineficiencia de este sistema de señales si cada molécula de glucosa liberada al medio requiriera para ello de una molécula de la hormona o señal enviada por la glándula responsable de este proceso.
La Figura 45 representa en forma esquemática cómo es posible que, en primer
lugar, una molécula de la hormona que representa la señal primaria enviada,
interactúe con una molécula y nada más que con una del receptor. Pero en seguida,
una molécula del receptor unido con la señal que ha recibido es capaz de activar
a la enzima adenilato ciclasa y a su vez cada una de éstas es capaz de catalizar
la conversión de un cierto número de moléculas de ATP en moléculas
de AMP cíclico. Para este paso de amplificación ya resulta que
una molécula de la hormona original es capaz de producir varios miles de moléculas
de AMP cíclico. Sin embargo la historia no termina ahí, cada una
de estas moléculas es capaz de activar a una enzima llamada proteína cinasa,
y da lugar a que ésta, actuando como una enzima, active muchas moléculas de
otra enzima, que en el caso que nos ocupa es la misma que se encarga de degradar
al glucógeno para liberar a la glucosa. Finalmente, cada molécula de esta enzima
es capaz de, actuando sobre su sustrato, en este caso el glucógeno, permitir
la liberación, a su vez, de varios miles o tal vez millones de moléculas de
glucosa.
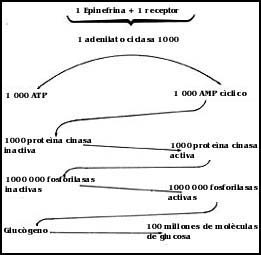
Figura 45. Esquema que muestra cómo se puede amplificar la señal de una molécula de epinefrina para que la célula receptora produzca millones de moléculas de glucosa. Las enzimas multiplican los efectos en cada caso.
De esta forma que hemos representado en forma esquemática resulta que una sola molécula de la señal original es capaz de producir una respuesta en forma de muchos millones de moléculas de glucosa liberadas al medio que rodea la célula y el procesamiento de la señal que así se inicia a nivel de la membrana celular consta en realidad de una serie de etapas de amplificación que dan lugar a que el efecto de unas cuantas moléculas de la hormona al final se presente en forma de millones de las moléculas que se pretendía liberar al medio, al enviarse la señal a la células adecuadas.
LOS MECANISMOS DE TRANSMISIÓN EN EL SISTEMA NERVIOSO
A pesar de que las señales utilizadas avanzan distancias enormes, las células
nerviosas se comunican entre sí por mecanismos que por una parte pueden considerarse
como existentes entre células vecinas. Esto se debe a que en el sistema nervioso
hay una situación especial: tal vez durante la evolución, el mecanismo seleccionado
más eficiente para la transmisión de órdenes de unas células a otras consistió
en la modificación de células distantes unas de otras, que resolvieron el problema
de su lejanía emitiendo largas prolongaciones, que podían permitirles entregarse
sus mensajes a través de ellas. Las prolongaciones desarrolladas por las células
nerviosas reciben en nombre de axones, y llegan a establecer contactos
con otras células a través de modificaciones especiales de la membrana celular
que permite establecer conexiones eficaces entre la célula efectora y la célula
receptora; estas estructuras que se desarrollan, y que representan modificaciones
tanto de la membrana de la célula emisora como de la receptora, reciben el nombre
de sinapsis. La figura 46 presenta una de estas células, en la que hay claramente
una modificación estructural. Primero, el cuerpo muestra prolongaciones de dos
tipos, las más cortas son las dendritas, que sirven para recibir señales, y
el axón, una gran prolongación, que sirve para enviar las señales. Estas se
transmiten gracias a lo que probablemente representa la manifestación de la
especialización de diversas regiones de la superficie celular, ya sea del cuerpo
celular, y/o de los axones y las dendritas para emitir por una parte y recibir
por otra mensajes de una a otra. El punto de contacto entre las células representa
una estructura especializada de las membranas de ambas que recibe el nombre
de sinápsis.
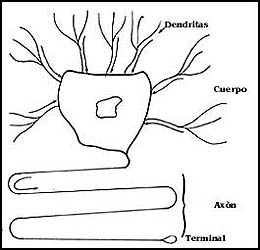
Figura 46. Esquema de una célula nerviosa. Los impulsos suelen llegar por las dendritas, terminaciones cortas que alcanzan al cuerpo celular. Los impulsos salen por el axón, que se representa plegado para indicar que es mucho más largo, hasta llegar a la sinapsis.
Para que se lleve a cabo la sinapsis en la porción que corresponde a la célula efectora o emisora, se acumulan vesículas recubiertas por membranas, en las cuales se encuentran las sustancias que habrán de servir como mediadores en la transmisión de las señales entre una célula y otra. Las células nerviosas, no obstante que establecen contactos unas con otras a través de sus axones, no se comunican en forma directa, sino de forma semejante a como lo hacen las glándulas del organismo que producen una hormona para modificar el funcionamiento o comportamiento de otra célula a distancia. En las células nerviosas las moléculas que representan propiamente la señal emitida de una célula a otra son liberadas también como respuesta a señales que ellas han recibido de otras células, de forma semejante a las hormonas. Las vesículas cargadas con la sustancia que representa la señal pueden fundirse con la membrana que se encuentra en la sinapsis y liberarla al exterior, para que ésta sea recibida por la célula receptora.
En el caso específico de las células nerviosas, la sustancia que se libera en el espacio de la sinapsis recibe el nombre de neurotransmisor, para distinguirla de las hormonas que son sustancias que viajan a través de la circulación para llegar a las células receptoras. Es muy claro, pues, que si el sistema de transmisión de los impulsos nerviosos de una célula a otra se vuelve mucho más efectivo y rápido que el sistema de transmisión de señales con sustancias del tipo de las hormonas. Las ventajas del sistema de transmisión de señales que existe en el sistema nervioso central son esencialmente dos: en primer lugar, la entrega de la señal directamente de la superficie de una de las células a la célula receptora, hace muchísimo más rápido el fenómeno mismo de la transmisión; la sustancia no debe viajar grandes distancias para llegar a la célula que ha de recibirla. En segundo lugar, dado que la sustancia transmisora de la señal se libera en un espacio extremadamente pequeño, basta con que la célula efectora o emisora libere cantidades relativamente pequeñas del neurotransmisor, para que éste alcance concentraciones muchísimo mayores que las que puede uno imaginar para una hormona; así este mecanismo se vuelve bastante más eficiente que el existente para las hormonas.
LA DESACTIVACIÓN DE LAS SEÑALES
Es muy claro que en las células deben existir mecanismos no solamente para
activar determinados procesos, sino también otros para desactivarlos. No es
posible imaginar que una señal enviada de una célula a otra no cuente con un
mecanismo de desactivación; las señales enviadas deben tener una duración limitada
para que cada una de ellas produzca modificaciones transitorias en la célula
receptora. Además, las células deben contar con mecanismos para desactivar las
señales; en el caso de las hormonas existen con frecuencia y en su mayor parte
para destruir a las moléculas efectoras que han llegado a las células, y así
hacer desaparecer la señal original. Por otro lado, las moléculas que han sido
activadas como resultado de la recepción de la señal también cuentan con mecanismos
propios de desactivación.
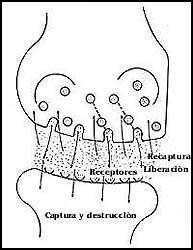
Figura 47. Los principales mecanismos de recepción e inactivación de los neurotransmisores a otras señales de comunicación. El transmisor debe ser eliminado para impedir que persista su efecto.
En los transmisores lo más frecuente es que se haga desaparecer la señal enviada de la sinapsis, sea por su destrucción o por la captura, sea por la misma célula que la emitió o por la célula receptora. Estos son los mecanismos que evitan que las señales se vuelvan modificadores perpetuos del funcionamiento de las células que las reciben.
Es posible que las magnitudes de los cambios producidos por diferentes neurotransmisores sean diferentes. Pero además, hay transmisión de impulsos excitadores e inhibidores, positivos o negativos, a carga de diferentes neurotransmisores. El sistema de los neurotransmisores se convierte así en algo semejante a los circuitos de cómputo, con señales positivas o negativas como base esencial del funcionamiento de cada unidad. La complejidad del funcionamiento se puede imaginar por algo así como 10 000 millones de neuronas, de las cuales cada una se comunica con otras 1 000, aproximadamente. Esto da en total una cifra aproximada de diez billones (10,000,000,000,000) de conexiones.
Otro de los elementos importantes en la comunicación de las neuronas es el
envío de las señales del cuerpo celular a las terminaciones sinápticas al final
del axón. Un axón puede tener varios centímetros de longitud, y el impulso nervioso
debe recorrer este camino para ser entregado en la terminación de axón a otra
célula. Una célula nerviosa envía sus impulsos a través del axón en la forma
de una onda que se llama "despolarización", y que consiste simplemente en permitir
el paso de iones de sodio y potasio por canales especiales y rápidos a través
de la membrana, dando lugar a que se invierta la corriente de estos iones acumulados
en el interior (K+) o expulsados al exterior (Na+) por
la acción de la ATPasa de Na-K. Esta despolarización se propaga
en forma de onda a una velocidad semejante a la del sonido a todo lo largo del
axón para terminar en la sinapsis, relevándose al ser convertida en la liberación
de un neurotransmisor a otra neurona. De esta manera se asegura una gran eficacia
y velocidad en la transmisión de los impulsos nerviosos a través de distancias
que en algunos casos son extremadamente grandes.