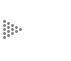VII. OTRAS FUNCIONES DE LAS MEMBRANAS
HASTA este punto se ha descrito toda una serie de funciones de las membranas que son más o menos importantes y claras para la mayoría de las personas, o que por su importancia resaltan sobre muchos otros procesos celulares, cada uno de los cuales puede resultar de tal interés que merezca la elaboración de un capítulo especial. Sin embargo, dentro de las funciones celulares que se localizan en las membranas todavía hay un número importante que merece mención especial, y que puede dar una idea más completa sobre la gran complejidad de su estructura y funcionamiento. Si lo mencionado hasta el momento no es suficiente para borrar en definitiva la vieja idea de las membranas biológicas como simples envolturas de las células o de organelos contenidos en ellas, tal vez la mención de unas cuantas funciones más podrá eliminarla sin lugar a dudas.
LA FORMACIÓN DE LA PARED CELULAR
Aunque —como hemos visto— todas las células tienen una membrana celular que las rodea, y ésta desempeña numerosas funciones como aislar y dar cierta protección al interior de la misma, hay cosas en las cuales esta protección debe ser en contra de factores adicionales, en especial los mecánicos y otras variaciones en el medio externo y el interno. Es fácil comprender la necesidad de una cubierta protectora para una célula ante los agentes mecánicos, pero hay un factor adicional que es difícil de imaginar, y lo podemos considerar de la manera siguiente: en el capítulo IV ya se mencionó el hecho de que los materiales disueltos en un líquido se comportan de manera semejante a un gas que se encuentra confinado en un cierto recipiente. La comparación que puede hacerse es válida también en términos de las presiones que ejercen los gases sobre las paredes de los recipientes que los contienen, y las soluciones cuando éstas se encuentran encerradas dentro de una membrana.
De acuerdo con lo expuesto en el capítulo IV, si tomamos un mol de oxígeno (02), que equivale a 32 gramos y que contiene un total de 6.02 x 1023 moléculas de este gas, encontraremos que a una temperatura de 25ºC y a una presión de una atmósfera ocupa un volumen de 22.4 litros. Esta misma situación ocurre si tomamos un mol de hidrógeno (H2), que equivale a sólo 2 gramos del gas. Para cualquier gas, en resumen, podemos tomar una cantidad en gramos, representada por el valor de su peso molecular, y la situación será exactamente la misma; un mol de él siempre ocupará un volumen de 22.4 litros a la temperatura de 25º C y una atmósfera de presión.
Por otra parte, sabemos bien que la presión de un gas es inversamente proporcional al volumen que ocupa. Esta es la razón por la cual aumenta su presión si lo comprimimos. Lo que se hace al inflar la rueda de un coche es precisamente aumentar la presión del aire, comprimiéndolo en su interior. Los valores del volumen de un gas y la presión a la que se le debe someter para mantenerlo son inversamente proporcionales; es decir, mientras mayor es el volumen menor es la presión, y mientras menor es el volumen mayor es la presión; además, la relación es tal que si se duplica la presión el volumen disminuye a la mitad o a la tercera parte y viceversa. Una forma simple de analizar este concepto es la siguiente: si un mol de oxígeno ocupa 22.4 litros a una presión de una atmósfera, al comprimir este gas en un volumen de un litro, la presión que ejercerá será de 22.4 atmósferas. (Una atmósfera es la presión que ejerce la capa de aire que nos rodea sobre todo lo que existe en la superficie de la tierra, por el simple efecto de la gravedad, y equivale a cerca de un kilogramo de fuerza por cada cm² de su superficie de cualquier cuerpo.)
De la misma manera que un gas ejerce una presión sobre el recipiente que lo
contiene, cuando se incluye una sustancia disuelta dentro de un volumen pequeño
ejerce también una presión sobre la pared o la membrana que lo rodea. Esto es
exactamente lo que sucede con la célula, y el experimento siguiente ilustra
la situación: si tomamos glóbulos rojos de cualquier animal o de un ser humano,
los separamos del plasma en que se encuentran con una centrífuga y los colocamos
en agua, debido a la cantidad de materiales disueltos que contienen y que no
pueden salir del mismo, se genera una presión de esas sustancias llamada presión
osmótica, la responsable de que, al no poder salir los materiales disueltos,
penetre una cantidad grande de agua, y las células terminen por estimar y perder
el material que contienen (Figura 48). Esto se puede evitar si simplemente se
colocan los eritrocitos en una solución de cloruro de sodio al 0.85%, también
conocida como "suero fisiológico" para que contrarreste la presión de los materiales
disueltos en el interior. La manera como nuestro organismo evita que esto suceda
es haciendo que el plasma sanguíneo, o sea el líquido en el cual se encuentran
los glóbulos rojos, tenga una concentración de sustancias disueltas semejante
a la que hay en el interior de los eritrocitos.
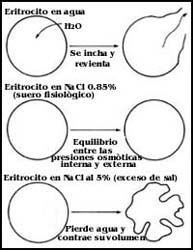
Figura 48. La hemólisis y su prevención. La falta de sales en el medio provoca
la entrada de agua; el eritrocito se hincha y revienta. El exceso de sal hace
que el glóbulo rojo pierda agua.
Este sistema de protección, que consiste en rodear a las células de soluciones más o menos concentradas de distintas sustancias, evita dañar a las células por los cambios bruscos del medio exterior, pero no existe ni en las plantas ni en muchísimos microorganismos de vida libre como las bacterias o las levaduras. Es muy frecuente que en el curso de su existencia, una bacteria o una levadura caigan en un medio que es prácticamente pura agua. Si su envoltura consistiera exclusivamente en la membrana celular, se romperían en el acto y dejarían de existir. Pero la naturaleza ha provisto a su membrana celular de enzimas que son capaces de producir polímeros, en general de azúcares, que adquieren forma fibrosa, se entrelazan alrededor de la membrana y llegan a constituir una capa protectora de gran resistencia.
Aunque hay muchas variantes, éste es en esencia el principio de la formación
y papel de la pared celular que hay en las células vegetales, en las bacterias,
hongos, etc. Su función consiste en evitar que la presión generada por la concentración
interna de los solutos, cuando las células se encuentran en soluciones diluidas,
dé lugar a su destrucción. La Figura 49 muestra la micrografía electrónica de
la pared de una célula, en la que puede verse el entrecruzamiento de las fibras
que la constituyen, y explica la gran resistencia de esa cubierta protectora.

Figura 49. Micrografía de una pared celular. Puede verse la estructura a
base de fibras entrelazadas, que adquiere gran resistencia. Cortesía del Dr.
Alfonso Cáravez; Instituto de Fisiología Celular de la UNAM.
La magnitud requerida de esta resistencia puede pensarse de manera muy simple. La concentración de solutos dentro de una levadura, por ejemplo, es de aproximadamente 0.7 molar, o sea 0.7 moles por litro. Si un mol por litro representa una presión de 22.4 atmósferas, esta concentración nos da una presión aproximada de 13.4 atmósferas. Para tener una idea cercana de la magnitud de esta presión baste pensar que la llanta de un automóvil se infla a una presión cercana a dos atmósferas. Así pues, la pared de un hongo debe resisitir más de 6 veces la presión que resiste el neumático de un coche.
Tal vez el caso más claro de la resistencia que una pared celular puede proporcionar a la célula esté representado por la de las esporas y la de los quistes, que son formas de vida latente, y proporcionan a diferentes organismos una capacidad enorme de resistir a condiciones adversas del medio ambiente. Inclusive hay enfermedades como la amibiasis y el tétanos que deben su abundancia precisamente a la resistencia que estas formas de los microorganismos adquieren ante el medio. Desde luego, en el caso de los quistes y las esporas no todo se debe a la pared celular; hay otros elementos desarrollados para aumentar la resistencia, pero tal vez el más importante sea la formación de una pared de extraordinaria resistencia.
Por otro lado, las paredes protectoras de las células representan barreras permeables para las sustancias que deben llegar a su interior. En su mayor parte permiten el libre paso de moléculas pequeñas, y sólo impiden el de moléculas grandes, como son las proteínas o polímeros de otras moléculas: los polisacáridos.
En las bacterias, por otro lado, los materiales de que está compuesta la pared celular con frecuencia les confieren propiedades que son de importancia, sobre todo desde el punto de vista de la respuesta o el reconocimiento que otros organismos pueden lograr cuando los invaden. Hay toda una serie de ellas, los neumococos, a los cuales los organismos animales reconocen o desconocen, gracias a los polisacáridos o polímeros de azúcares que constituyen su pared celular. Estas características son aún de importancia para que los médicos reconozcan el tipo de bacteria que puede haber invadido a un individuo.
La penicilina, por otro lado, actúa porque las bacterias resisten una gran variedad de condiciones gracias a su pared celular, sintetizada por las enzimas de su membrana. Lo único que hace el antibiótico es inhibir a una de las enzimas que sintetizan esa pared y evita que se forme la capa protectora; como resultado de ello las células mueren al encontrarse en un medio con una menor presión osmótica.
LAS CÉLULAS SE PUEDEN RECONOCER UNAS A OTRAS
Tal vez el caso más primitivo de reconocimiento celular esté representado por los microbios llamados mixobacterias, que son microorganismos del suelo y se alimentan de moléculas grandes insolubles. Lo interesante de ellos es que se asocian formando grupos grandes, en los cuales las células se encuentran unidas entre sí, y siempre se mueven juntas; al encontrar moléculas grandes, contribuyen todas a digerirla y a utilizar los productos de esa digestión.
Aparentemente hay dos elementos importantes en esta movilidad de las células como grupos. Por una parte, cuando una bacteria se desplaza deja una especie de rastro de una sustancia que produce y elimina al exterior, que puede ser reconocida por sus congéneres. Por otro lado, parece que cada célula se puede mover cuando se estimula por el contacto con otras. En el mecanismo de organización del grupo de células parecen existir genes que dan lugar a la formación de sistemas de señalamiento para atraerse entre sí; hay también otros que actúan como receptores de esas señales y finalmente deben existir los elementos mismos que dan lugar a la movilidad de las bacterias. En los estudios sobre el fenómeno se han utilizado mutantes en las cuales se pierden distintas capacidades cada vez, para llevar el fenómeno a su forma final.
Este comportamiento "social" de las mixobacterias parece modificarse inclusive cuando el alimento se agota. Cuando esto sucede, las células no sólo se mueven juntas sino que se agregan para formar grupos y constituir una especie de órgano primitivo en el cual se convierten en esporas y se cubren de una sustancia que las rodea. Así, las esporas son capaces de sobrevivir durante largos periodos de tiempo aún en condiciones totalmente adversas. Cuando llegan a encontrarse en condiciones favorables, germinan y producen nuevamente las bacterias. El carácter múltiple de estos órganos rudimentarios o esporangios asegura que al germinar las esporas se formará nuevamente un grupo de individuos y no individuos aislados.
También se han hecho experimentos para definir no sólo la capacidad que tienen
las células de reconocerse entre sí, sino también de adherirse unas a otras.
Se ha estudiado, por ejemplo, a las esponjas, que son tal vez los animales multicelulares
más sencillos que existen y que constan de sólo unos cuantos tipos diferentes
de células que se pueden disociar oprimiendo al individuo contra una malla.
Las células intactas se separan así para ver luego su capacidad de asociarse.
Al volver a mezclar las células disociadas, se agregan rápidamente, y eventualmente
se reorganizan para formar una esponja normal (Figura 50). Pero no sólo existe
la capacidad de reagregarse y reorganizarse, sino que las células de especies
diferentes también lo hacen, dependiendo de la especie de la que provienen.
Se pueden mezclar células obtenidas de esponjas de especies diferentes, y el
resultado final es que las células de especies diferentes se reconocen entre
sí, y no se asocian con las células de la otra especie (Figura 50).
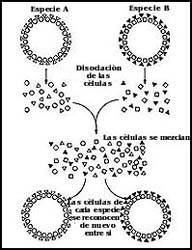
Figura 50. El reconocimiento y la asociación de las células de las esponjas.
Las células sólo reconocen a las de su misma especie, gracias a sus sistemas
membranales.
En el caso de la agregación de las esponjas se ha aislado inclusive la molécula
responsable producida por las esponjas mismas, que es muy grande y requiere
de calcio para funcionar como enlace entre las distintas células, que al parecer
se ancla a receptores específicos que existen en las membranas de las células
de la misma especie. Es posible que esta sustancia actúe entonces como puente
entre los receptores que se encuentran en la superficie de unas y otras células.
El fenómeno consta así de dos partes: primero, la presencia de receptores que
son específicos para cada especie, y luego, una molécula que es producida por
las mismas esponjas que es capaz de reconocerlos y formar puentes entre dos
células, uniéndolas (Figura 51).
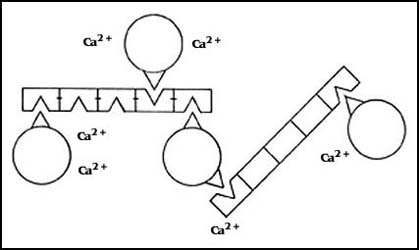
Figura 51. Las moléculas que unen a las esponjas. Son producidas por las
células. Son específicas para cada especie. Forman agregados que requieren calcio.
Las células tienen receptores específicos que las reconocen.
En los experimentos realizados, en los que se utilizan células embrionarias disociadas, también se ha logrado encontrar un fenómeno semejante, pero además de la impresión de ser específico para cada uno de los tejidos u órganos de un individuo.
LOS MECANISMOS DE RECONOCIMIENTO EN ORGANISMOS SUPERIORES
Se han hecho experimentos en los cuales se disocian las células embrionarias de animales vertebrados por diferentes medios. En ellos se utilizan estas células por el simple hecho de que las células adultas son muy difíciles de separar. Después de haberlas disociado se les vuelve a mezclar; si se toman células de dos tejidos diferentes, en un principio forman agregados mixtos, pero posteriormente empiezan a separarse, agrupándose de acuerdo con el tejido que les dio origen. Hay moléculas en la superficie que en forma específica les indican a cuáles células deben asociarse. Esto se deduce porque cuando se mezclan células embrionarias de dos especies diferentes, todavía se agrupan según el tejido de donde provienen, y se asocian sin importar la especie a la que pertenecen, sino esencialmente el tejido del que formaron parte.
UNA BACTERIA EN BUSCA DE SU ALIMENTO
Aunque parece muy simple que una bacteria se desarrolle si le toca en suerte encontrarse un medio rico en materiales nutritivos, y que muera si no es ésta su suerte, la realidad es diferente: hay muchas bacterias que son capaces de dirigirse a donde existen mejores condiciones para su alimentación. Este fenómeno es conocido con el nombre de quimiotaxis, y puede permitir a una bacteria acercarse al medio más rico en materiales nutritivos, o inclusive alejarse de ambientes que puedan resultarle nocivos. Es claro desde luego que esto sólo ocurre con las bacterias móviles que cuentan generalmente con flagelos.
La Figura 52 no representa esquemáticamente los movimientos de una bacteria
que se encuentra en presencia de una sustancia capaz de atraerla, en comparación
con el trayecto que sigue cuando se encuentra frente a otra que no es capaz
de atraerla, o bien esencialmente porque representa un material que no es nutritivo.
Cuando la bacteria se encuentra cerca de un material nutritivo, cuya concentración
aumenta en alguna región cercana, lo que hace es nadar hacia él de manera irregular;
para ello, hace girar sus flagelos en un sentido. Cuando los materiales del
medio les son indiferentes, lo que hace es dar saltos en direcciones al azar
haciendo girar sus flagelos en sentido contrario. Tal vez sea interesante señalar
que de ninguna manera se mantiene quieta, sólo que cuando no hay materiales
nutritivos o atrayentes, nada sin un rumbo fijo, como si explorara el ambiente
que la rodea. El estudio de este comportamiento ha permitido definir algunos
de los componentes involucrados. El primero de ellos es un receptor capaz de
"sentir" la presencia de un material atrayente; se han aislado mutantes que
son capaces de comportarse normalmente frente a prácticamente cualquier estímulo,
excepto alguno de ellos. Este resultado indica que la lesión reside solamente
en el receptor a estas sustancias, puesto que si simplemente se cambia una sustancia
por otra que sí atraiga a la bacteria, el comportamiento es normal en todos
sentidos. Hay otro tipo de mutantes que impiden que las bacterias respondan,
no a uno, sino a un grupo de sustancias atrayentes. Esto ha dado lugar a que
los receptores se agrupen u organicen en grupos y tengan un sistema de
interconexión con el sistema completo de la respuesta de la bacteria. Se piensa
que el receptor se encuentra conectado con otra proteína de la membrana, y que
funciona como intermediario para generar un mediador intracelular, que se comportaría
de manera semejante al AMP cíclico en el caso de los receptores
a algunas hormonas. Sería un segundo mensajero entre el sistema receptor y el
que hace funcionar a los flagelos para moverse según un esquema adecuado para
seguir ese estímulo.
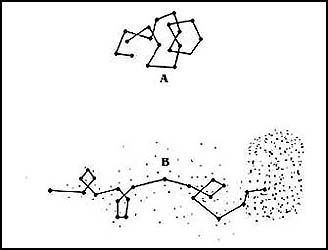
Figura 52. Una bacteria que no es atraída, se mueve al azar(A). Al ser atraída,
su movimiento, aunque no en línea recta, la lleva al sitio de mayor concentración
del material nutritivo (B). El movimiento, aun en ausencia de un material nutritivo,
le permite "explorar" el medio.
Pero hay otro elemento interesante en el comportamiento de las bacterias frente a una fuente alimenticia. Una vez que se encuentran en un ambiente adecuado, y sus movimientos no las llevan a regiones más ricas en el material que las ha atraído, parecen adaptarse a él y dejan de desplazarse, moviéndose nuevamente al azar y sin dirección fija en el mismo lugar. A este fenómeno se le conoce con el nombre de adaptación, y ha sido objeto de interesantes estudios que han permitido saber qué sucede cuando la bacteria llega a un medio ambiente en el cual ya no cambia la concentración del material nutritivo. Parece ser que cuando esto sucede, uno de los componentes del sistema se modifica químicamente, a través de un proceso de metilación, impidiendo que la bacteria continúe su búsqueda.
LOS MICROBIOS EN GUERRA, LOS ANTIBIÓTICOS Y LAS TOXINAS
Con frecuencia, el medio ambiente es incapaz de permitir la vida de un número grande de organismos. Aun un medio nutritivo rico tiene límites en cuanto al número de organismos que puede alimentar, y con frecuencia son diferentes tipos de ellos los que buscan disfrutarlo y desarrollarse en él. Cuando esto sucede, frecuentemente no hay otro camino que la guerra: entre animales de la misma o diferentes especie que no permiten a otros invadir su territorio, entre plantas que producen y liberan al suelo sustancias tóxicas para otras plantas; y por desgracia también suele suceder que la riqueza de una zona es la que provoca disputas y hasta guerras entre humanos.
Pero los microorganismos no por ser pequeños dejan de participar de esta regla general de comportamiento en la biología; es tal la abundancia y dispersión de ellos, que con enorme frecuencia no es una sino varias especies las que invaden un medio nutritivo rico. Por lo tanto, es también enorme la frecuencia con la que se enfrentan y disputan la posesión de ese ambiente dos o más especies de microorganismos, y para ganar la guerra muchos de ellos disponen de armas muy eficaces contra los demás. Estas son sustancias que segregan al medio y paralizan o causan la muerte de los demás microorganismos y hasta de plantas y animales; en conjunto reciben el nombre de antibióticos y toxinas.
Hay una gran diversidad de mecanismos por los cuales los antibióticos son capaces de bloquear el crecimiento de otros microorganismos o de detener el metabolismo de distintos tipos de células. Existen inhibidores de la respiración, inhibidores de la síntesis del ATP, inhibidores específicos de ciertas enzimas, pero hay algunos que específicamente producen efectos en relación con las membranas. Como ejemplos mencionaremos solamente el caso de dos de ellos: la penicilina, el primer antibiótico conocido, y cuyo uso ha persistido durante tantísimos años como un útil agente contra las infecciones; el otro tipo de sustancias corresponde a los antibióticos que son capaces de comportarse como sistemas de transporte y afectan a las membranas en forma directa.
Aunque en la vida diaria se habla de antibióticos que son benéficos y se administran a los humanos en la lucha contra las infecciones y otras enfermedades, es necesario saber que éstos son el resultado de una selección entre un gran, número de ellos que se han aislado. Una buena parte de los antibióticos que se han descubierto, además de impedir el crecimiento de otras bacterias, resultan tóxicos para las células animales o vegetales. Ello hace imposible su utilización para el tratamiento de infecciones u otras enfermedades en las que participan las bacterias. Sin embargo, hay unos cuantos que por diversas razones tienen la capacidad de atacar sólo a ciertos microorganismos sin producir ningún daño a las células animales, y se utilizan en caso de infecciones para destruir a las bacterias que las causan, sin dañar a las células animales. La penicilina, por ejemplo, es un antibiótico que no afecta precisamente a las membranas biológicas, sino a las enzimas involucradas en la síntesis de la pared celular de muchas bacterias. Este mecanismo consiste sencillamente en la inhibición de una enzima, responsable de uno de los pasos en la síntesis de la pared celular. Si las bacterias habitualmente se encuentran en ambientes en los que la concentración de sales es inferior al de su propio interior, como ya se mencionó, la pared representa un elemento fundamental para protegerse y mantenerse vivos. Por el simple hecho de inhibir la formación de la pared, la penicilina produce la muerte de las bacterias, debido a que impide la formación de la cubierta protectora que los defiende de los efectos de las diferencias de presión osmótica entre su interior y el medio en que se encuentran.
En el capítulo III ya se mencionó que algunos antibióticos pueden comportarse como acarreadores de iones en las membranas. Hay una variedad importante de ellos que no se utilizan en el tratamiento de las infecciones porque son tóxicos también para las células animales; sin embargo, su mecanismo de acción es interesante. La valinomicina, por ejemplo, es una molécula que puede colocarse en la membrana de cualquier célula por poseer un exterior hidrofóbico. Por otro lado, su interior es hidrofílico, y le permite abarcar un átomo del ion potasio, uno de los constituyentes más importantes, prácticamente, de todas las células.
De esta manera, cuando una célula se encuentra en presencia de la valinomicina, ésta se concentra en su membrana y es capaz de producir, por la diferencia de concentraciones entre el interior y el exterior, la salidad del ion potasio, produciendo así alteraciones en el funcionamiento de los sistemas enzimáticos celulares y finalmente la muerte de las células. Como ya se mencionó, la valinomicina no se utiliza en el control de infecciones debido a que es tóxica para casi todos los organismos y las células animales, por esa razón no puede utilizarse selectivamente contra las bacterias. Del mismo tipo de la valinomicina hay otros antibióticos como la gramicidina, la alameticina, la nonactina, etc., que tienen propiedades más o menos semejantes y todas actúan provocando la salida de los iones del interior de la célula, sobre todo el potasio. Es importante mencionar el caso de la gramicidina, cuyo comportamiento es semejante al de la valinomicína, pero sí se utiliza en el tratamiento local de algunas infecciones, como las amigdalitis bacterianas.
Otro grupo más de antibióticos es capaz de intercalarse en las membrana y producir poros grandes a través de los cuales salen prácticamente todas las moléculas pequeñas que contienen. Este es el caso de la filipina y la nistatina, sustancias utilizadas en el tratamiento de las infecciones por hongos, a los que parecen destruir por este mecanismo.
También hay otras sustancias producidas por bacterias que atacan a las células
animales. Aunque no hay diferencias importantes entre éstas y los antibióticos,
a través de los años se les ha diferenciado, llamándoles toxinas, con diferentes
acciones. Por ejemplo, la toxina producida por el bacilo del cólera tiene efectos
sobre las membranas celulares. Esta toxina actúa en forma importante sobre los
mecanismos de transmisión de señales; cuando esto ocurre se fija sobre la proteína
G, que activa y desactiva la adenilato ciclasa (capítulo VI), e impide la desactivación
del sistema completo, produciendo serias alteraciones sobre el funcionamiento
celular (Figura 53).
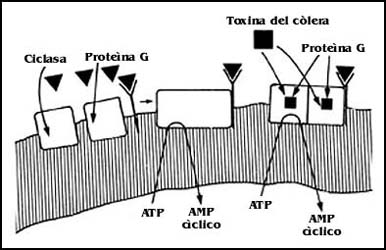
Figura 53. El efecto de la toxina del cólera. La toxina se fija a la proteína G, y le impide separarse. El sistema entonces no se puede desactivar. Esto provoca la salida de Na+ y agua al intestino y fuertes diarreas que pueden llevar a la deshidratación y a la muerte a los enfermos.
Hay muchas otras toxinas que no tienen efecto sobre las membranas, pero que también tienen extraordinaria importancia, como las de la difteria, la tosferina, la tifoidea, etcétera.