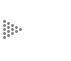II. LAS UNIDADES Y LA ESTRUCTURA DE LAS MEMBRANAS
PARA formar las membranas biológicas, la naturaleza aprovechó las propiedades de unas sustancias que no pueden incluirse estrictamente dentro de las que hemos llamado polares o no polares. Se trata de compuestos que en su misma molécula tienen una parte polar y otra no polar. A este grupo pertenecen, por ejemplo, los jabones y los detergentes, que están formados por una cadena apolar, o "cola", de átomos de carbono e hidrógeno, y una "cabeza", que tiene una carga negativa o positiva. Hay un grupo numeroso de estas sustancias, a las cuales se llama anfifílicas o anfipáticas (del griego anfi: ambos), porque parecen mostrar simpatía o, en términos reales, capacidad para interactuar, por una parte, con sustancias polares y, por otra, con sustancias apolares. A continuación veremos cómo se comportan estas sustancias en diferentes condiciones, pero principalmente en el agua, que es el solvente principal de los seres vivos, así como las influencias que algunos otros componentes de las soluciones en que se encuentran, como los iones, pueden tener en su comportamiento.
Si disolvemos moléculas de jabón en agua, como se muestra en la figura 9, lo que se produce no es una solución verdadera; de hecho no es transparente, y al colocarla contra la luz la vemos más o menos turbia u opalina, porque las moléculas realmente no se han disuelto, sino que se han organizado en forma de partículas muy pequeñas. Si la vemos iluminándola de un lado en una habitación oscura nos daremos cuenta que dispersa la luz (se ve luminosa al observarla de un lado), precisamente por la presencia en ella de esas partículas de cierto tamaño. El fenómeno es semejante a la luminosidad de las partículas de polvo iluminadas por un rayo de sol en una habitación más o menos oscura.
La formación de esas partículas tiene lugar porque las cadenas de átomos de
carbono o "colas" hidrofóbicas de las moléculas, como cualquier cadena hidrofóbica,
son rechazadas por el agua; esto provoca que todas se agrupen, dando lugar a
las estructuras llamadas micelas, que son minúsculas esferas en las cuales las
"colas" o porciones apolares forman la parte central, y hacia el exterior se
sitúan las "cabezas" o porciones polares, que sí pueden interactuar con el agua
(Figura 9). Esta propiedad de formar micelas no es más que una de las manifestaciones
de las interacciones de los detergentes y jabones con el agua. Si colocamos
jabón o detergente sobre la superficie del agua, se puede formar una capa monomolecular
(de una sola molécula de grueso), en la cual las "colas" hidrofóbicas, rechazadas
por el agua, quedan expuestas al aire, y las "cabezas" hidrofílicas sumergidas
en el agua. Pero más aún, las paredes de las burbujas de jabón no son otra cosa
que una doble capa de jabón o detergente con las colas orientadas hacia el aire
y las cabezas hacia la capa de jabón, en contacto también con una finísima capa
de agua, que, además, se evapora con facilidad y da como resultado la pérdida
de la estructura y la ruptura de la burbuja a pocos segundos de que se ha formado.
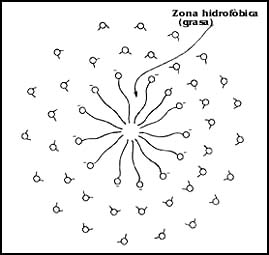
Figura 9. Las micelas, estructuras que forman las moléculas de jabón en
el agua, y su relación con ésta.
No es difícil imaginar ahora que las propiedades de las sustancias anfifílicas difieran según su estructura individual, y que estas diferentes propiedades modifiquen también las de las micelas u otras estructuras que forman. El estudio de las propiedades de las micelas, por otra parte, ha servido de base para suponer en un principio y confirmar luego muchas de las propiedades de las membranas biológicas que, como veremos a continuación, obedecen a las mismas reglas generales de comportamiento y comparten algunas de las propiedades generales con las micelas.
Las micelas también han permitido hacer estudios sobre las características generales de las superficies que ofrecen al medio, no sólo frente al solvente mismo, sino también frente a otros componentes o condiciones del medio ambiente, como la acidez o la alcalinidad (pH), o la presencia de otras sustancias, principalmente iones de diferentes cargas o valencias, etc. Esta información en muchas ocasiones ha permitido establecer principios o normas generales de comportamiento que se pueden aplicar a otras estructuras semejantes, incluyendo a las membranas biológicas.
Parte de la información básica que se ha obtenido de las micelas es la relativa al estado físico de las porciones apolares rechazadas por el agua. El estudio de la fluidez del interior de las micelas ha permitido predecir propiedades importantes de las membranas biológicas por extrapolación, en cuanto a su estado físico, acomodo, efectos del largo de la cadena apolar, de la presencia de dobles ligaduras en ella, de la carga de las cabezas, de la presencia de iones en el agua, etcétera.
LAS MEMBRANAS COMO CAPAS DOBLES DE FOSFOLÍPIDOS
Hay un tipo de sustancias anfifílicas o anfipáticas que tienen semejanza con las mencionadas y mostradas en la Figura 1; sin embargo, difieren de ellas esencialmente porque en lugar de una "cola" apolar, tienen dos. La naturaleza ha elaborado diversas clases de ellas, y corresponden al grupo de los fosfolípidos.
El nombre proviene del hecho de que la cola hidrofóbica corresponde en su estructura a la de una grasa (lípido), y en la "cabeza" polar se encuentra un grupo fosfato (fosfo) que contiene fósforo (representado por el símbolo P) en su estructura.
Uno de los hechos interesantes en relación con estas moléculas es que la mayoría
de ellas comparte propiedades comunes, pues tienen dos cadenas hidrofóbicas
(aunque pueden ser más), que son de longitud variable, pero las más comunes
tienen entre 16 y 18 átomos de carbono y un número variable de dobles enlaces
entre los átomos de carbono. En la composición de la "cabeza" hay una mayor
variabilidad, no sólo en la estructura misma, sino también en cuanto al número
y al signo de las cargas eléctricas. En la Figura 10 se representan sólo tres
de estos fosfolípidos, y se intenta señalar que puede haber diversidad en las
cabezas y en las colas. De hecho, en una molécula de un fosfolípido, las dos
cadenas de átomos de carbono no son necesariamente iguales.
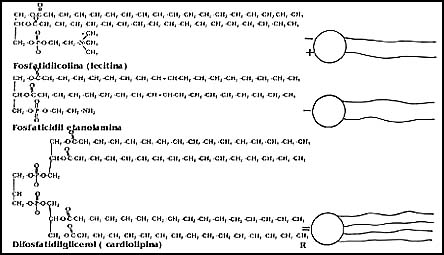
Figura 10. Distintos tipos de fosfolípidos. Nótese que para la lecitina
mostrada, las "colas " son dos moléculas de ácido oleico, de 18 átomos de carbono
con una doble ligadura. Para la cardiolipina se presenta una molécula que tiene
cuatro moléculas de ácido palmítico, de 16 átomos de carbono.
¿CÓMO SE COMPORTAN EN EL AGUA?
Estas moléculas no se dispersan con facilidad en el agua como los detergentes
y los jabones; sin embargo, por medios físicos, como con el ultrasonido, se
puede lograr su dispersión. Como resultado de ello, éstas se organizan formando
vesículas pequeñísimas, cuyas paredes están formadas por una doble capa de fosfolípidos,
como se muestra en forma esquemática en la Figura 11. A estas vesículas se les
ha dado el nombre de liposomas, y muestran propiedades interesantes, que se
asemejan muchísimo a las membranas biológicas.
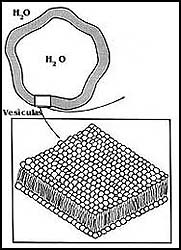
Figura 11. Esquema que muestra las vesículas que forman los fosfolípidos,
así como el acomodo de éstos para formar la estructura membranosa.
Las vesículas formadas artificialmente con fosfolípidos o liposomas, son estructuras cerradas que no permiten el paso de sustancias iónicas o polares ni de moléculas grandes en ningún sentido. Curiosamente, tal y como sucede con las membranas biológicas, sí son permeables al agua. Estas características han hecho que los liposomas, formados con diversos tipos de fosfolípidos, se hayan utilizado muchísimo para estudiar las propiedades de las membranas. Se ha llegado inclusive a reproducir en ellos muchos de los fenómenos que se encuentran en las membranas naturales de las células (véase el capítulo VIII).
LAS MEMBRANAS CELULARES ESTÁN FORMADAS CON CAPAS DOBLES DE FOSFOLÍPIDOS
Desde 1925, los investigadores Gorter y Grendel propusieron que las membranas de las células están formadas por una capa doble de fosfolípidos. La propuesta resultó de un experimento muy sencillo: estos autores aislaron los fosfolípidos de una cantidad conocida de glóbulos rojos (eritrocitos) y los colocaron sobre la superficie del agua, de manera que formaran una capa de una sola molécula de grueso, y midieron el área. Tomando en cuenta la forma y las dimensiones de los glóbulos rojos, calcularon el área de la superficie de los eritrocitos de donde provenían los fosfolípidos y encontraron que el área de los fosfolípidos dispersos era el doble de la calculada para las células. Con este sencillo experimento y una serie de consideraciones teóricas, basadas en las propiedades de las moléculas, tal y como las hemos descrito hasta ahora, estos investigadores propusieron entonces que los fosfolípidos deberían estar formando una capa doble en la membrana, además, con las porciones apolares opuestas y alejadas del agua, como se mostró en la Figura 11. Esta proposición no recibió la atención debida, y pasaron muchos años antes de que se le diera la importancia y el valor que merecía.
Una buena parte de la evidencia de que las membranas celulares están formadas por una capa doble de fosfolípidos proviene de experimentos llevados a cabo con bicapas artificiales de fosfolípidos, en las que se puede simplificar enormemente el problema y hacer estudios que en las células íntegras serían complicados. Pero la mayor parte de las observaciones realizadas en las bicapas artificiales se pueden aplicar a las membranas de las células.
Cuando se muestra la imagen de una membrana obtenida por microscopía electrónica, se observa una doble sombra que hace ya muchos años se interpretó en el sentido de que los lípidos estaban en el centro de la membrana y que sus dos superficies estaban recubiertas con proteínas. Sin embargo, esta misma imagen se observa en las bicapas artificiales, en las cuales no hay proteínas; se concluye que la doble sombra no representa otra cosa que las superficies de las dos capas de fosfolípidos.
Hay un procedimiento en el cual se puede congelar una muestra para observarla a través del microscopio electrónico, pero antes se le fractura en lugar de teñirla directamente, generando dos superficies. Todas las observaciones realizadas con membranas permiten concluir que la doble capa se fractura en la parte central que tiene menor resistencia, en donde la fuerza de cohesión sólo está dada por la débil atracción de las porciones polares de los fosfolípidos.
La conclusión de que las membranas de las células están formadas por una doble capa de fosfolípidos no se ha alcanzado por medio de argumentos aislados. Como ante cualquier problema científico, sólo es posible concluir algo si evidencias numerosas señalan lo mismo. Lo mencionado es sólo una parte de las pruebas de que las membranas son así. Hay muchos otros elementos, tanto teóricos como experimentales, que coinciden, de modo que en la actualidad no hay duda de que las membranas de las células tienen como estructura básica esta doble capa de fosfolípidos.
También debe quedar claro que esta estructura existe cuando las células se encuentran en el agua. Es sólo la relación recíproca del agua y de los fosfolípidos la que da lugar a la organización mencionada; si el agua desaparece, deja de existir la doble capa. Aunque por ejemplo hay manera de secar ciertos microorganismos, la realidad es que no puede hacerse completamente; siempre es necesario dejar un mínimo de agua para mantenerlos vivos.
¿QUE PROPIEDADES TIENEN LAS BICAPAS?
Impermeabilidad. Dado que la más importante de las funciones de las membranas es tal vez la de formar una capa protectora alrededor de las células, la impermeabilidad es una de las principales propiedades de las bicapas que se debió demostrar. No sólo ha sido posible demostrar la impermeabilidad, sino también algo que debía esperarse a partir de la composición grasa o lipídica de la membrana. Hace aproximadamente 40 años, Collander, haciendo investigaciones con el alga Nitella, encontró que las sustancias pueden pasar a través de la membrana en muy estrecha relación con su solubilidad en las grasas. Esto es precisamente lo que debía esperarse si la membrana equivale en cierta forma a una capa de grasa que rodea a la célula.
Por otra parte, las propiedades de las membranas no sólo son de interés para satisfacer la curiosidad de los investigadores. El conocimiento puede ser interesante hasta en el caso, por ejemplo, del diseño de medicamentos que deben actuar dentro de las células; para ello lo ideal es que su estructura les permita penetrar a las células porque se "disuelven" en la membrana. Esto también es cierto para algunos tóxicos, cuyos efectos se deben en gran parte a los efectos que tienen sobre las membranas. Uno de estos casos es la lesión que el tolueno y otros solventes orgánicos producen sobre las membranas de las células, y que parecen compartir con los solventes industriales que algunos individuos inhalan, utilizándolos como enervantes.
Sin embargo, en el mismo experimento realizado por Collander fue muy claro que el agua difiere del comportamiento general de otras sustancias. Siendo poco soluble en aceite, entra con gran velocidad a las células. Este comportamiento no tiene una explicación clara hasta ahora, pero sí es un hecho que tanto las membranas biológicas como las bicapas artificiales tienen una gran permeabilidad al agua. En los organismos animales, por ejemplo, no hay membrana celular alguna que el agua no pueda atravesar; inclusive hay algunas que además tienen mecanismos para aumentar esta permeabilidad; como en el caso, por ejemplo, de la piel de la rana o de algunas células de riñón en las cuales los movimientos del agua además se pueden regular por medio de hormonas.
Otra propiedad importante de las membranas es la elasticidad. Las células y otras estructuras membranosas contenidas en ellas deben ser elásticas. Por numerosas razones sería inconveniente tener membranas rígidas, que impedirían el movimiento o al acomodo de las células. La Figura 12 muestra una imagen de una amiba y la de un alga, obtenida con el microscopio electrónico de barrido; es claro que esta estructura no se lograría con una membrana rígida.
La elasticidad de la membrana parece deberse a una razón muy simple: sucede que las cadenas de átomos de carbono de los fosfolípidos que se encuentran en el centro de la doble capa se comportan como las moléculas de cadenas libres, y tienen una gran movilidad, como si se encontraran en estado líquido. Esto, por otra parte, se puede demostrar por medio de diversos procedimientos; además, es posible ver que esta "fluidez" o estado físico de las membranas, como para cualquier sistema, depende de la temperatura.
Resistencia. Aunque pudiera pensarse que las membranas celulares son frágiles, éste no es el caso; su resistencia es considerable; esto es cierto inclusive para las membranas formadas artificialmente por un sólo tipo de fosfolípido, como la fosfatidiletanolamina (Figura 12).
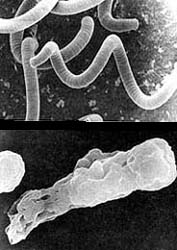
Figura 12. Micrografía de una amiba y de un alga (Spirulina maxima) tomada
en el microscopio electrónico de barrido por el Dr. Alfonso Cárabez, del Instituto
de Fisiológia Celular de la UNAM. Es de hacer notar la gran capacidad
que la membrana tiene para plegarse sin romperse.
Pero hay otros elementos que pueden agregar resistencia a la bicapa, como pueden
ser, por ejemplo, las fuerzas de atracción entre las cabezas de los fosfolípidos
con diferente signo en su carga. Tal es el caso de las moléculas de lecitina
(Figura 10), que tienen una carga positiva y una negativa en la porción polar
y se atraen entre sí. Aún en el caso de moléculas con un solo signo en la carga,
como la fosfatidiletanolamina, que lógicamente presentan repulsión de sus cabezas
cuando se encuentran en presencia de Ca2+ o de Mg2+, estos
cationes, no sólo neutralizan las cargas, sino que forman además puentes entre
ellas, transformando la condición desfavorable para aumentar la atracción y
con ello la resistencia de la membrana (Figura 13).
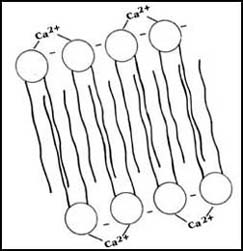
Figura 13. Esquema que muestra cómo el ion de calcio (Ca2+),
que tiene dos cargas positivas, puede aumentar la estabilidad de una membrana
formada por fosfatidil serina, cuyas moléculas tenderían a repelerse por tener
carga negativa.
Pero las propiedades generales mencionadas se pueden obtener prácticamente con cualquiera de los fosfolípidos. Sin embargo, en las distintas clases de células hay un número muy grande de diferentes fosfolípidos; sin duda esto debe tener una significación, pero sólo en algunos casos se ha llegado a demostrar que, por ejemplo, ciertos sistemas de transporte requieren precisa y estrictamente algunos tipos de fosfolípidos. En el caso, por ejemplo, del transporte del azúcar galactosa en Escherichia coli (el colibacilo), éste sólo funciona con membranas preparadas con los fosfolípidos que tiene la bacteria. Al cambiarlos por otros, el transpostador no funciona. Algo semejante se ha encontrado en algunos componentes de los sistemas respiratorios mitocondriales y en algunas bacterias que forzosamente requieren de ciertos fosfolípidos para su buen funcionamiento. Aunque el asunto no se ha estudiado con suficiente detalle, en la naturaleza es difícil encontrar sustancias o fenómenos que no tengan un significado e importancia precisos. El caso de los diferentes tipos de fosfolípidos de seguro no escapa a esta regla general.
LAS PROTEÍNAS DE LAS MEMBRANAS
Aunque nos hemos limitado a mencionar con detalle a los lípidos como los componentes de las membranas de las células, esto es simplemente una forma de presentación del tema. Los lípidos constituyen la estructura básica de la membrana, actúan como una película de recubrimiento y aislante, que evita el paso de la mayor parte de las sustancias que se encuentran dentro o fuera de una célula. Pero en términos de utilidad, de nada serviría una membrana concebida así; ya señalamos que se requieren elementos que permitan, entre otras cosas, el intercambio con el exterior, pero no sólo de sustancias, sino también de otros tipos de interacciones, como la comunicación, el contacto, etc. Es aquí donde intervienen las proteínas.
Las proteínas son tal vez las estructuras más complicadas que hay en las células. Están formadas por la unión de 20 unidades diferentes, los aminoácidos. A partir de estas unidades se forman moléculas más grandes, unidas siempre de acuerdo a un orden definido en cada célula para cada proteína. El tipo de enlace es el peptídico.
Dado que cada aminoácido tiene características especiales de tamaño, signo y número de cargas, hidrofobicidad, etc., al formarse cadenas entre ellos, se produce una organización especial que se traduce en estructuras de gran complejidad, cuyo papel dentro de las células es realizar funciones complicadas. Por ejemplo, cada transformación química de una sustancia en otra es prácticamente promovida o catalizada por una proteína, llamada enzima, y hay miles diferentes de ellas.
Las proteínas son en realidad lo que podríamos considerar como las piezas mismas de la maquinaria celular, y las que se encargan de mover cada paso del complicadísimo mecanismo que da lugar al funcionamiento integral de las células.
La situación es variable, pero el contenido de proteínas de las membranas varía entre un 25 y 75%; esto parece depender del grado de especialización y actividad de cada una de ellas. Una de las que tiene mayor contenido de proteínas es la mitocondrial, y es explicable, pues este organelo celular tiene a su cargo entre otras cosas, nada menos que la función de proveer de energía al resto de la célula.
Se utiliza a las proteínas en las funciones membranales, de la misma forma que para realizar directamente las funciones más complicadas de las células. Sólo estas sustancias alcanzan un grado de complejidad suficiente para realizar tan delicadas funciones como, por ejemplo, colocarse en la superficie de la célula y reconocer primero en el exterior a una molécula de glucosa, tal vez entre otras 1 000 o 2 000 sustancias, capturarla luego y finalmente introducirla a la célula, en ocasiones en contra de su concentración y tendencia natural a moverse, debido a que en el exterior puede estar menos concentrada que en el interior.
Las proteínas, como componentes de la maquinaria celular, también son las que ejecutan en las membranas la mayor parte de las funciones propiamente dichas. Sobre la estructura básica esencial de una doble capa de lípidos, impermeable y relativamente inerte, fue necesario implantar las unidades funcionales que dieran sentido a las membranas como fronteras dinámicas y que les permitieran un contacto eficiente y adecuado con el exterior. Sólo las proteínas tienen la capacidad de alcanzar este grado de especialización.
En la Figura 14 se presenta de manera esquemática el acomodo de las proteínas
en las membranas. Aunque algunas pueden estar simplemente adosadas a ellas,
la mayoría las cruzan de un lado al otro, porque así lo requieren las funciones
que realizan en la Figura 1 de la introducción, ya se mostró un esquema de algunas
de las funciones de estos componentes de las membranas.
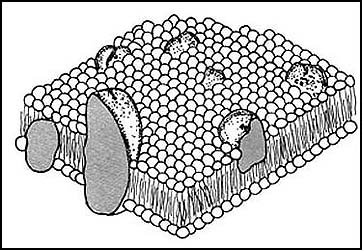
Figura 14. Esquema que muestra cómo se intercalan las proteínas en las membranas.
Las proteínas se presentan en forma más o menos compacta, aunque en realidad
están constituidas como se mostró en la figura 11.
LOS CARBOHIDRATOS EN LAS MEMBRANAS
Son numerosas las membranas celulares que contienen carbohidratos (azúcares). En la mayoría de los casos se trata de cadenas de los azúcares simples o sus derivados, llamados polisacáridos, que llegan a ser extremadamente complejas. Una de las funciones importantes de estas sustancias parece consistir en servir de elementos de reconocimiento por parte de otras células u organismos. Tal vez el caso más conocido sea el de los polisacáridos de los glóbulos rojos, que por la presencia en sus membranas dan lugar a los grupos sanguíneos. La cuestión es muy simple, hay dos polisacáridos, el A y el B; la presencia de ninguno, uno o ambos puede dar lugar a los tipos sanguíneos O, A, B, y AB, con las consecuencias que ello tiene para las transfusiones sanguíneas. Al administrar un tipo de glóbulos rojos diferente a los que tiene un individuo, su organismo los desconoce y los destruye, y el mecanismo de reconocimiento tiene como base la presencia de estos polisacáridos.
Es impresionante el gran número de cadenas de azúcares que sobresalen de la superficie de un eritrocito aunque no se conoce el papel más que de una parte de ellas.
En resumen, las membranas no sólo contienen lípidos; hay también carbohidratos y proteínas en proporciones variables: sin embargo, las funciones "activas" de las membranas parecen estar a cargo de las proteínas, como es el caso de un gran número de otras funciones celulares.