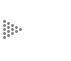III. EL PASO DE LAS SUSTANCIAS A TRAVÉS DE LAS MEMBRANAS
HAY SUSTANCIAS que las células no necesitan y deben ser eliminadas; hay otras en el exterior que deben tomar para nutrirse. A la gran mayoría de ellas les está prohibido, ya sea por su naturaleza polar o por su tamaño, cruzar la bicapa lipídica de la membrana. Antes que otras funciones más complicadas de las células, está la de su propia nutrición y excreción. Con este fin primordial se desarrollaron los sistemas de transporte; moléculas o grupos de ellas, generalmente proteínas, que en ocasiones funcionan como poros selectivos, permiten simplemente el paso de las sustancias, o en otras, inclusive, las "obligan" a entrar o salir, según las necesidades de la célula.
CÓMO SON LOS SISTEMAS DE TRANSPORTE
Fuera de que los sistemas de transporte conocidos están constituidos por moléculas de proteína, con qué velocidad se mueven y cuál es su afinidad por las sustancias que acarrean, etc., poco es lo que se sabe acerca de su mecanismo íntimo de funcionamiento. Nadie tiene idea en detalle de cuál es la estructura de una proteína que formada por una cadena de aminoácidos, como todas ellas, tenga la capacidad de reconocer y luego permitir, de manera selectiva, el paso de ciertas sustancias; sólo se han llegado a imaginar esquemas que aceptamos como modelos para seguir estudiándolos. Una parte de los resultados logrados en el área del transporte biológico proviene de investigaciones realizadas utilizando algunos antibióticos que, a pesar de que no son los sistemas de transporte celular en sí, se comportan como tales y han permitido imaginar algunas de las características de los transportadores naturales o verdaderos. Su estudio ha proporcionado información sobre algunos de los mecanismos para explicar la selectividad para ciertas sustancias o la estructura de un poro en la membrana. Su comportamiento es tan interesante, que vale la pena hacer un paréntesis sobre estas sustancias.
LOS ANTIBIÓTICOS, ARMAS BIOLÓGICAS Y HERRAMIENTAS EN LA INVESTIGACION
Durante varias décadas se han estudiado muy diversos tipos de antibióticos, que son sustancias producidas por ciertos microorganismos y capaces de alterar el funcionamiento de otros seres vivos o incluso de producir su muerte. Tal vez la mejor conocida de estas sustancias sea la penicilina, que detiene el desarrollo bacteriano al bloquear la síntesis de la cápsula que cubre a la membrana celular de numerosas bacterias. Al impedirse la formación de esta cápsula protectora, la bacteria se vuelve frágil y fácilmente muere por diversos factores, el principal de ellos es la presión que su contenido ejerce sobre la membrana, produciéndose la muerte por lisis.
Hay antibióticos que son dañinos para algunas bacterias e inocuos para los animales o los humanos, y son los que tienen utilidad para tratar las enfermedades. Pero en la búsqueda de antibióticos eficientes para el tratamiento de enfermedades se ha encontrado también un gran número de ellos que sí son tóxicos y, de hecho, no se pueden emplear porque, aunque sean efectivos contra las bacterias, también producen la muerte de las propias células de los animales o de los humanos. Esto es desde luego lógico, pues a fin de cuentas los antibióticos son las armas que permiten a los microorganismos asegurar su predominio en el medio ambiente sobre los otros seres vivos.
Es realmente raro encontrar antibióticos que no sean tóxicos para los animales en general, y los humanos en particular. En la búsqueda de sustancias de este tipo que puedan ser benéficas para el tratamiento de las enfermedades producidas por microorganismos, se han encontrado muchísimos antibióticos que sí son tóxicos para los animales, y que no se pudieron emplear nunca como agentes terapéuticos, pero al estudiarlos desde otros puntos de vista, se encontraron propiedades sorprendentes que explican su mecanismo de acción tóxica. Tal es el caso de una serie de ellos que han sido de gran utilidad para el estudio de los fenómenos de transporte en sistemas biológicos.
LOS ANTIBIÓTICOS LLAMADOS IONÓFOROS
En el año de 1964, Moore y Pressman iniciaron los estudios de la valinomicina
que producía la salida de potasio en mitocondrias obtenidas de las células de
hígado de rata, y esto era causa de muchas otras alteraciones consecuentes en
el funcionamiento de ese organelo celular. En forma resumida, los estudios de
estos autores se extendieron a otros sistemas biológicos, y se llegó finalmente
a la conclusión de que su efecto tóxico reconocía como mecanismo el que se presenta
en la Figura 15. Esta sustancia tiene una estructura semejante a una "dona"
o rosca, pero muy peculiar; es hidrofóbica en el exterior e hidrofílica o polar
en el interior: además, su diámetro interior corresponde con gran precisión
al del ion potasio (K+). Como otros iones, el K+ normalmente
no puede atravesar la membrana porque se encuentra rodeado de una capa de moléculas
de agua; esta característica es la que lo vuelve insoluble en la membrana y
le impide el paso (véase el capítulo II). Lo que hace la valinomicina es rodear
al K+ y, dado que por fuera es hidrofóbica, puede cruzar la bicapa de un lado
al otro. Cuando las mitocondrias se colocan en un medio en el que no hay potasio,
y tomando en cuenta que ellas tienen una gran cantidad de este ion en el interior,
es fácil imaginar que la valinomicina pueda tomarlo del interior y llevarlo
al exterior con facilidad. De esta manera, con una molécula relativamente pequeña
se pudieron hacer miles de experimentos que permitieron, además, sentar bases
más sólidas en cuanto al mecanismo de funcionamiento de los llamados acarreados
móviles.
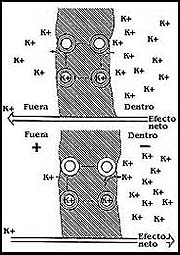
Figura 15. Esquemas que muestran lo que hace una molécula de valinomicina. En la parte superior se presenta el mecanismo por el cual el antibiótico puede vaciar el contenido de iones de potasio. En la parte inferior se ve que, si hay un potencial eléctrico negativo dentro, el antibiótico puede introducir K+ y acumularlo en concentración superior a la externa.
Con este sistema de transporte tan sencillo, se demostró que la molécula del antibiótico es capaz de reconocer al K+ con una afinidad 5 000 veces mayor que al sodio, y ello casi sólo se debe a las dimensiones del orificio interno de la molécula. Aunque pueden caber otros iones, esto sólo tiene lugar con dificultad cuando son demasiado grandes, o si son más pequeños, la fijación a los sitios del antibiótico se vuelve débil y no se pueden transportar o lo hacen con una eficiencia muchísimo menor.
También se demostró con este sistema que el antibiótico, por su característica hidrofóbica en el exterior, se fija a la membrana y ocupa la región hidrofóbica. Ahí puede "asomarse" al exterior y capturar a los iones de potasio. Luego se mueve con ellos en el interior de la bicapa y los suelta hacia el lado de menor concentración. Pero puede suceder algo que es aún más interesante; si existe una diferencia de potencial eléctrico a ambos lados de la membrana, la valinomicina puede soltar el catión en el lado de la membrana en que el potencial es negativo, aun en contra de lo que se esperaría por la diferencia de concentración; es decir, la valinomicina puede producir la acumulación del ion si hay un potencial eléctrico.
Otro de los antibióticos que se estudió posteriormente fue la gramicidina.
El caso de esta sustancia es diferente, porque, para empezar, es una cadena
lineal de aminoácidos; sin embargo, y sin entrar en los detalles de cómo se
realizaron los experimentos, se pudo demostrar que este antibiótico también
puede funcionar como ionóforo, pero su mecanismo de acción es distinto. Varias
moléculas de gramicidina se acomodan una sobre otra, enroscándose, de modo que
entre varias pueden formar un túnel que cruza la membrana, constituyendo un
poro que puede permitir el paso de los iones de determinado tamaño y características
(Figura 16).
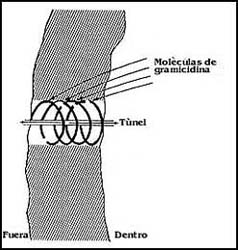
Figura 16. La gramicidina y su acomodo en la membrana. Este antibiótico, no obstante de ser una cadena de varios aminoácidos, puede enrollarse y asociarse con otras moléculas para formar un canal a través de la membrana.
Estos son sólo dos de los casos de antibióticos que se han estudiado; hay una larga lista de ellos que además son capaces de permitir el paso de distintos iones a traves de las membranas. Está el caso inclusive de la nistatina o la filipina, que al formar poros tan grandes en las membranas, por ellos llegan a salir componentes mucho mayores que los iones. El efecto de estos antibióticos es, por tanto, permitir el paso de todos los componentes pequeños de las células, y sólo les quedan las proteínas y otras moléculas grandes.
Con estos antecedentes y una gran cantidad de experimentos realizados por numerosos investigadores de todo el mundo, se ha llegado a concebir la existencia de dos tipos principales de acarreadores o sistemas de transporte: los canales o poros y los acarreadores móviles.
Como se pretende mostrar en la Figura 17, hay sistemas de transporte que se
imaginan como canales o poros. No se conoce el mecanismo preciso del funcionamiento
de ninguno de ellos; sin embargo, a través de su función se ha llegado a un
modelo imaginario. El poro o canal es más que nada la conceptualización de un
sistema rápido de transporte; es más fácil imaginar el movimiento rápido como
flujo a través de un túnel, que por un mecanismo de acarreo más complicado.
La idea del poro está representada en el diagrama de la figura; y se piensa
que debe tener antes que nada una especie de entrada o filtro capaz de discriminar
o escoger entre distintas sustancias o iones. Por ejemplo, hay poros que pueden
distinguir fácilmente el Na+ y K+ a pesar de su semejanza, pero que difieren
por el tamaño. Otra de las características importantes de los poros es la existencia
de una especie de "compuertas" o dispositivos que les permiten abrir y cerrarse
al paso de los iones.
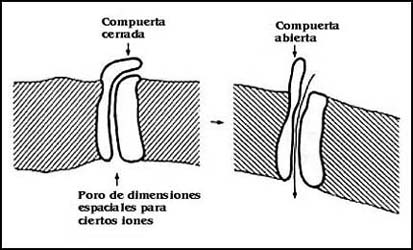
Figura 17. Esquema de un canal. Además de un túnel de dimensiones
precisas, los canales tienen compuertas que pueden abrirse o cerrarse por diferentes
mecanismos.
Es claro que el caso de la gramicidina es relativamente simple, pues esta sola sustancia debe satisfacer las características de selectividad (sólo acepta cationes monovalentes) y de comportamiento como compuerta (sólo forma el poro cuando hay una diferencia de voltaje en ambos lados de la membrana). Aunque el modelo que ofrece la gramicidina para su estudio ha sido muy interesante y ha permitido establecer ciertas bases, todo indica que los sistemas de transporte que existen, por ejemplo, en el músculo o el nervio, o cualquiera otra célula animal, son mucho más complicados que este antibiótico.
En el caso de sistemas de transporte más lento, se ha imaginado que se trata
de moléculas de proteína que situadas en la membrana cuentan con un sitio capaz
de reconocer a las sustancias que han de transportar, de manera semejante como
las enzimas tienen un sitio activo en el que se coloca el sustrato que van a
modificar. En este sentido no habría diferencia con los poros. Sin embargo,
como se muestra en la Figura 18, el sistema del paso de los iones de un lado
al otro sería diferente; sin que se conozca el mecanismo íntimo, se piensa que,
o bien la molécula de la vuelta y el sitio activo que estaba hacia un lado de
la membrana se desplaza al otro, o bien el ion u otra sustancia es movido al
otro lado por movimíentos peristálticos, semejantes a los del intestino, "exprimiendo"
a la sustancia transportada hacia el otro lado. El único hecho real que hay
es que estos sistemas de transporte son mucho más lentos que los canales o poros.
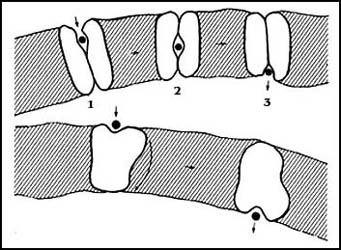
Figura 18. Esquemas de un acarreador móvil. Éstas son las representaciones de dos mecanismos que se han propuesto para el funcionamiento de los acarreadores móviles. La valinomicina es también un acarreador móvil (fig. 16).
Se ha tomado a la valinomicina como modelo de este tipo de acarreadores; el término móvil implica la necesidad de que la molécula se mueva dentro de la membrana, como en el caso del antibiótico. Sin embargo, no se conoce gran cosa respecto del mecanismo de los acarreadores móviles que hay, por ejemplo, en las células animales.
LA ESPECIFICIDAD DE LOS SISTEMAS DE TRANSPORTE
Si bien es cierto que los sistemas de transporte son esenciales para que las células intercambien materiales con el exterior, también lo es que uno o varios mecanismos indiscriminados de este tipo podrían matarlas en poco tiempo. Un ejemplo real puede ser el de los antibióticos ya mencionados, la nistatina y la filipina, que precisamente matan a las células porque forman poros por los cuales pueden salir todas las moléculas pequeñas, iones, etcétera.
Es pues necesario que los sistemas de transporte del tipo que sean, canales o acarreadores móviles, tengan una alta selectividad, es decir que de entre las moléculas presentes en las soluciones que bañan a las células sean capaces de reconocer a las que necesitan, sea en forma individual y altamente selectiva o por grupos.
Los sistemas que se han estudiado, con variaciones dentro de ciertos límites, son específicos; es decir, guardan un alto grado de exigencia con respecto a las sustancias que transportan. Por ejemplo, los sistemas de transporte para el fosfato, la forma más común de aprovechamiento del fósforo por células y organismos, son capaces de discriminar y no transportan ni cloruros, ni sulfatos, ni otros aniones de estructuras más o menos lejanas. Sin embargo, y esto es desafortunado, el sistema puede ser engañado, por así decirlo, ofreciéndole arsenato, sustancia mucho más parecida al fosfato, pero por otro lado, altamente tóxica, pues es la forma más común del arsénico que se puede ofrecer a los seres vivos. El caso del sistema de absorción o transporte de los azúcares en el intestino delgado es de menor exigencia; aunque no todos los azúcares simples, llamados monosacáridos, son transportados con la misma velocidad, son muchos los que se pueden mover, aparentemente por el mismo sistema; sólo se requiere que ofrezcan una estructura básica más o menos común.
Hay también diferencias entre los sistemas de transporte en cuanto a la abundancia
o concentración que requieren de las sustancias que transportan en el medio
del cual las toman. El asunto tiene importancia en el caso, por ejemplo, de
bacterias u otros microorganismos que pueden sobrevivir en medios extremadamente
pobres. Para ello necesitan de sistemas de transporte capaces de tomar con gran
eficiencia las moléculas de materiales nutritivos que requieren para vivir y
que se encuentran en escasa cantidad en el medio. Pero hay también sistemas
de transporte que no necesitan de tal capacidad para capturar unas cuantas moléculas
que existen en el medio ambiente. Esto es lo que sucede con muchos sistemas
de transporte de las células animales; en cierto modo se puede pensar que no
se requieren sistemas de alta afinidad para muchas de las sustancias, puesto
que viven en un medio interno que está regulado en cuanto a las concentraciones
de sustancias que contiene, y, en muchos de los casos, éstas son relativamente
elevadas.
Figura 19. La selectividad y la afinidad de un sistema de transporte, que debe distinguir, entre muchísimas otras, a unas cuantas moléculas dispersas, que son las que debe mover a través de la membrana.
Entre los microorganismos hay casos muy interesantes, pues algunas bacterias pueden tomar sustancias del medio ambiente que se encuentran en concentraciones extremadamente pequeñas. Hay incluso casos, por ejemplo, de microbios que son capaces de contar con varios sistemas de transporte para la misma sustancia. Así sucede con la levadura, que, cuando crece en un medio rico en potasio, desarrolla un sistema de transporte de una afinidad relativamente baja. Sin embargo, cuando crece en un medio pobre en este ion, el sistema de transporte que utiliza para tomarlo del medio tiene una afinidad 1 000 veces mayor, aproximadamente.
Otra de las características importantes de los sistemas de transporte es la velocidad con la que funcionan, como en el caso de las enzimas del metabolismo celular, donde hay sistemas más veloces que otros. Con frecuencia se encuentra que los sistemas con mayor afinidad muestran velocidades menores y viceversa.
Pero además, existe siempre una relación entre la abundancia o concentración de una sustancia y la velocidad con la que es transportada por una célula. Es obvio que si el fenómeno de transporte depende de la interacción de la sustancia a transportar con su acarreador, a medida que aumenta la concentración, la velocidad del transporte es mayor. Pero esto tiene límites; cada interacción individual del sistema de transporte tiene una duración que no se puede acortar indefinidamente, y debe llegar el momento en que el sistema de transporte sature su capacidad, es decir, a su velocidad máxima de funcionamiento.
Descritas en estos términos tan sencillos, las moléculas de los acarreadores son, sin embargo, sistemas de una extraordinaria eficiencia, que tal vez fueron los primeros componentes de las membranas en aparecer. Es necesario pensar que los primeros seres vivos sobre la faz de la tierra fueron los microorganismos de vida libre, y la abundancia de muchos de los materiales en el medio ambiente no era grande. Aparentemente de este hecho es de donde resultó una gran especialización de los sistemas que tenían que encargarse de transportar esos materiales tan escasos, para poder asegurar la supervivencia de los organismos.
En muchos de los microorganismos conocidos, además, no es raro encontrar que
cuentan con varios sistemas de transporte para una sola sustancia. Ya se mencionó
el caso del transporte de K+ en la levadura que aparece en dos formas: una que
le permite aprovechar este catión cuando se encuentra en concentraciones más
o menos grandes, y otro para poder utilizarlo cuando las concentraciones son
extremadamente pequeñas en el medio. Pero lo interesante no es sólo eso; el
hecho es que la célula cuenta con un sistema de control que se muestra en forma
esquemática en la Figura 20, según el cual sólo produce el sistema de transporte
de mayor afinidad (que requiere para captar el ion cuando las concentraciones
son bajas), si previamente se ha cultivado en un medio pobre en potasio. Éste
es sólo uno de los casos que hay en la naturaleza; son numerosos los sistemas
de transporte que, por así decirlo, sólo aparecen en las células cuando existen
los materiales a transportar.
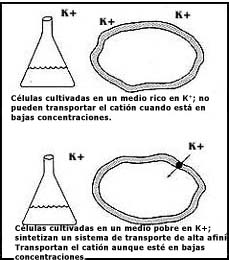
Figura 20. Esquema que muestra el mecanismo para regular la aparición de un sistema de transporte de alta afinidad en algunos microorganismos.
Ya en la realidad, la forma como funciona este mecanismo de control es regulando la síntesis misma del acarreador de alta afinidad; para no desperdiciar energía, la célula sólo lo produce cuando se encuentra en un medio pobre de este catión.
El grado de especialización de los sistemas de transporte de las células ha llegado a ser tan grande en algunos casos, que durante su evolución se le pudo adaptar para funciones de gran importancia como la transmisión nerviosa. En otros casos, como se describe en el capítulo siguiente, la clara relación entre la energía y el transporte llevó, por un lado, a la existencia de sistemas de transporte capaces de aprovechar la energía metabólica para tomar del medio sustancias que se encuentran en concentraciones bajas, pero también fue a través de los sistemas de transporte modificados que se pudieron diseñar sistemas para la conversión de la energía en formas directamente aprovechables por las células.
También sucede que muchos receptores en las células pueden verse como sistemas de transporte capaces de reconocer a ciertas moléculas, algunas con gran afinidad y especificidad. De esta forma, puede pensarse que las células y los organismos modificaron sus sistemas originales de transporte para dar lugar a los receptores y contar con la posibilidad de comunicarse entre sí y de ahí diseñar sistemas de control.
El sistema nervioso mismo reconoce como base un complicadísimo sistema de intercomunicación de las neuronas, pero que en último análisis tiene fundamentalmente la participación de un enorme número y tipo de sistemas de transporte y receptores.
Hay inclusive un tipo especial de receptores que son capaces de detectar la presencia de moléculas disueltas o volátiles y que se encuentran en células especializadas. Éstas son las células receptoras de los órganos de los sentidos; el olfato y el gusto también pueden considerarse resultado de la evolución de los sistemas de transporte, en los que la modificación de los antiguos acarreadores permitió desarrollar estas capacidades que en los animales han llegado a ser tan importantes.