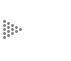IV. TRANSPORTE Y ENERGÍA
EL COMPORTAMIENTO DE LAS MOLÉCULAS EN SOLUCIÓN
TODOS conocemos el hecho aparentemente sencillo y trivial de que si colocamos una gota de tinta en un vaso de agua, las partículas de colorante se mueven o difunden lentamente hasta que en un tiempo, más o menos largo, ocupan todo el espacio de que disponen y la solución se vuelve completamente uniforme. Esto, que se puede ver con los colorantes, es el fenómeno llamado difusión; sucede con todas las sustancias por igual, y se debe a una razón muy sencilla: las moléculas de todas las sustancias tienen energía que las mantiene en constante movimiento, y que hace que tiendan a desplazarse de los lugares de mayor a menor concentración, llegando a dar una concentración uniforme en todas las partes de una solución al final de cierto tiempo.
Esta energía que todas las moléculas tienen, se puede manifestar en diferentes
formas. Tal vez las más claras evidencias se observen al comprimir un gas. Este
procedimiento consiste en forzar una sustancia gaseosa a ocupar un espacio menor
del que tenían antes; y es tal el estado energizado, que podemos realizar diferentes
tipos de trabajo con la energía del gas comprimido; hay herramientas muy variadas
que se impulsan con aire comprimido. Lo mismo sucede con el vapor de agua cuando,
en lugar de dejarlo libre al hervir agua, lo encerramos en un espacio; la presión
que alcanza puede ser tan alta que también con él podemos realizar un trabajo
utilizando una máquina de vapor. Exactamente de la misma forma, las moléculas
de una solución ejercen una verdadera presión sobre las paredes del recipiente
que la contiene, por sus constantes choques; no hay diferencia en la manera
como las moléculas de un gas la ejercen sobre el recipiente que lo contiene.
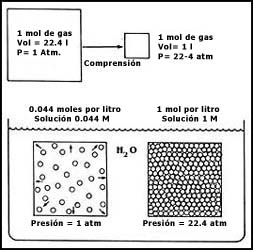
Figura 21. Equivalencia entre la presión de un gas y la de una solución. La presión osmótica se pone fácilmente de manifiesto cuando las soluciones se colocan en un medio que permite el paso del agua, pero no el de la sustancia disuelta.
Es entonces necesario no perder de vista este concepto: todas las moléculas de una solución, como las de un gas, tienen una cierta energía, y la cantidad de ella depende del volumen que ocupa una cantidad de moléculas o iones; es obvio que mientras tengamos un mayor número de ellas en un cierto volumen, será mayor la cantidad de energía. En otras palabras, la energía de las moléculas de una solución depende de la concentración (cantidad por volumen) de ella. Esta energía hace que las moléculas tiendan a difundirse de los lugares de mayor a los de menor concentración, igual que el aire comprimido tiende a escapar del recipiente en que se encuentra almacenado, perdiendo así energía en el proceso. También, aunque pueda parecer difícil imaginarlo, en el proceso mediante el cual una solución se diluye, hay una liberación de energía que en lugar de liberarse podría ser aprovechada para realizar un trabajo o cambiarla en otra forma. Lo contrario también es válido; para que un grupo de moléculas pase de un lugar de menor a uno de mayor concentración es necesario aplicar energía al sistema, como si se tratara de comprimir un gas.
Hemos utilizado hasta ahora el término transporte para designar simplemente al hecho de que, si la mayor parte de las membranas impide el paso de la mayor parte de las sustancias, para atravesarlas, éstas deben ser ayudadas o "transportadas" de un lado al otro; sin embargo, además del impedimento físico mismo que una membrana representa, el proceso de transporte también tiene implicaciones energéticas.
TRANSPORTE "ACTIVO" Y "PASIVO"
Durante muchos años se habló del transporte en estos términos; se decía que era "activo" cuando dependía del metabolismo celular y, en general, cuando:
—Una sustancia en general, metabolizable por las células, como la glucosa, necesitaba de una fuente de energía.
—Se inhibía con los llamados venenos metabólicos o por circunstancias que inhibían el metabolismo, como la disminución de la temperatura.
Esto apunta hacia un hecho: en algunos casos, el transporte depende del funcionamiento
del metabolismo. Pero éste sirve, entre otras cosas, para producir energía,
y el transporte celular requiere de ésta para funcionar. Esto ya indicaba que
la relación entre el metabolismo celular y el transporte pudiera ser la energía
producida en uno y necesaria para mover al otro de los procesos.

Hay otro tipo de transporte "pasivo", que se distingue del activo porque no es sensible a los factores que alteran a éste. Es el caso, por ejemplo, del transporte de azúcar en los eritrocitos o en algunos otros sistemas biológicos. Para que éste tenga lugar, lo único que se requiere es que la concentración de azúcar en el exterior sea mayor que la del interior. Lógicamente, si la concentración en el exterior es inferior a la interior, el azúcar sale en lugar de entrar. Hay así muchos sistemas de transporte en los cuales las sustancias se mueven sin necesidad de utilizar energía, simplemente siguiendo la tendencia natural de la difusión, que va del sitio de mayor al de menor concentración.
Son numerosos los sistemas de transporte que tienen una relación de dependencia
absoluta con el metabolismo. En la actualidad sabemos que esa dependencia se
debe al requerimiento de energía para el transporte. En su mayoría, estos sistemas
de transporte son aquellos en que se trata de lograr una mayor concentración
de una sustancia en un lado de una membrana que del otro. Realmente, la relación
entre el metabolismo y el transporte no es otra cosa que la energía, y el metabolismo
representa la fuente directa de ésta en las células, que se requiere para impulsar
el transporte cuando tiene lugar en contra de la tendencia natural de difusión
de una sustancia (Figura 23).
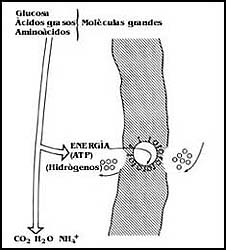
Figura 23. Energía, transporte y metabolismo. Al degradarse las moléculas
que intervienen en el metabolismo, se produce energía. Ésta puede ser utilizada
por los sistemas de transporte para introducir ( o sacar) sustancias de la célula.
Aunque hay un número enorme de caminos metabólicos, es decir, de series de transformaciones de unas sustancias en otras en las células, hay algunas de estas vías que tienen más directamente a su cargo la generación de formas de energía seguramente aprovechables por las células. Unos de ellos, que se encuentra en muchos sistemas biológicos es la cadena respiratoria, que se realiza, por ejemplo, en las mitocondrias de todas las células y en las membranas de las bacterias. El otro está representado, en su forma más sencilla, por un camino que degrada la glucosa para convertirla en ácido láctico o en alcohol, a través de una serie de pasos que dan como resultado una molécula que es fácilmente aprovechable por algunos sistemas de transporte de las células, y cuyas características veremos más adelante.
La cadena respiratoria recibe ese nombre porque consiste en un complicado sistema de moléculas que toman átomos de hidrógeno y electrones de diferentes sustancias que las células obtienen de la degradación de los materiales con los que se nutren. A través de los componentes de la cadena respiratoria, estos hidrógenos y electrones viajan hacia el oxígeno, con el cual se combinan al final. En cierta forma, este proceso puede verse como la manera en la que las células llevan a cabo la combinación del oxígeno con el hidrógeno para formar agua, y es realmente lo que constituye la respiración celular. Pero en el proceso se puede obtener una cantidad muy grande de energía derivada, en términos muy sencillos, de la gran tendencia que tiene el hidrógeno para unirse con el oxígeno, que todos conocemos.
La cadena respiratoria se realiza precisamente en las membranas de las mitocondrias
de las células, o en la membrana externa de las bacterias. Como puede verse
en la Figura 24, cuando los átomos de hidrógeno o en algunas partes de la cadena,
los electrones que provienen de ellos viajan hacia el oxígeno, liberan una cantidad
de energía, que en lugar de disiparse, se utiliza para mover hidrogeniones (H+)
de un lado al otro de las membranas.
Figura 24. La cadena respiratoria. Las moléculas provenientes del metabolismo
ceden pares de átomos de hidrógeno que liberan energía al combinarse con el
oxígeno. Esta energía no se desperdicia, sino que se utiliza para mover
protones (hidrogeniones) hacia un lado de la membrana.
Como se indica en la figura, este transporte de H+ (hidrogeniones) representa en gran medida la forma de "atrapar" o convertir la energía derivada de la tendencia natural de los electrones a llegar al oxígeno, en otra forma de energía, la diferencia de concentración de los H+ en ambos lados de la membrana. La tendencia natural de los hidrogeniones que se han concentrado de un lado de la membrana representa una nueva forma de energía, que luego puede ser aprovechada para mover a otras sustancias, como se verá más adelante.
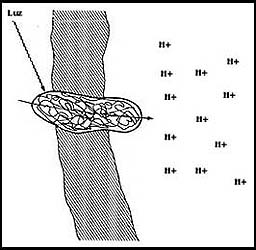
Dentro del grupo de acarreadores directamente ligados a las fuentes de energía,
hay uno que tiene un interés muy especial: la bacteriorrodopsina. Ésta es una
molécula que existe en algunas bacterias y es capaz de generar el movimiento
de H+ de un lado a otro de la membrana, creando también una diferencia de concentración
de esta especie química, pero para ello basta que se le ilumine; es decir, puede
aprovechar directamente la energía luminosa para mover y producir diferencias
de las concentraciones de los H+ (Figura 25).
El ATP (adenosín trifosfato) es una sustancia que existe en todos
los seres vivos y tiene una gran importancia porque funciona como fuente directa
de energía para muchos procesos.
Hay motores que funcionan con gasolina y otros que lo hacen con diesel, o hay máquinas de vapor que utilizan carbón o leña como combustible. A todos nos queda claro que no es posible hacer funcionar un motor de gasolina con diesel ni al revés. La razón es muy simple: el diseño de las máquinas es tal que los combustibles no son intercambiables. De la misma forma, las células no pueden utilizar directamente hidrógeno ni glucosa para mover, por ejemplo, sus sistemas de transporte. Para muchos de ellos, y para otros, las células requieren de un combustible especial y sólo éste les puede proporcionar directamente la energía que requieren para moverse. Este combustible se llama ATP, abreviatura química que significa adenosín trifosfato.
El ATP es una molécula que tiene en su composición tres moléculas
de fosfato unidas entre sí, pero que en su enlace tienen una gran cantidad de
energía, utilizable por diversos sistemas. Cuando el ATP empleado
por enzimas u otros sistemas adecuados se rompe, se produce ADP (adenosín
difosfato) y una molécula de fosfato, como se muestra en la Figura 26, liberándose
en ese proceso la energía contenida en el enlace. Hay muchos sistemas biológicos
que pueden descomponer el ATP, que se llaman ATPasas,
pero todos tienen en común la capacidad de utilizar la energía del enlace del
último fosfato de la molécula para diferentes fines.
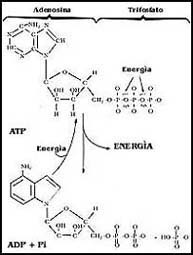
Figura 26. La estructura del ATP (adenosín trifosfato). La energía se produce
al romperse para dar ADP (adenosín difosfato) y fosfato inorgánico (Pi).
En la Figura 27 se muestra un tipo diferente de transporte primario utilizando
como fuente de energía el ATP. Las células cuentan con moléculas
o sistemas de ellas capaces de mover iones a través de las membranas, empleando
esta energía de la ruptura de los fosfatos del ATP (figura 27).
Hay toda una familia de proteínas que se encuentran en muy diferentes sistemas
biológicos que reciben el nombre genérico de ATPasas, es decir,
enzimas que rompen el ATP; muchas de ellas tienen esta propiedad,
usando la energía de su ruptura (hidrólisis), para transportar iones de un lado
a otro de las membranas.
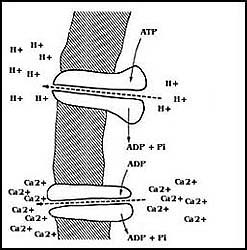
Figura 27. Las ATPasas o "bombas" del transporte. Estas son enzimas que pueden mover iones a través de las membranas biológicas utilizando la energía de la hidrólisis (ruptura) del ATP. Por este mecanismo pueden crear diferencias enormes de concentración y de carga eléctrica a los lados de la membrana.
Como ejemplo tenemos los siguientes:
|
ATPasa de H+
|
Para mover protones (H+)
|
|
ATPasa de Na — K
|
Para mover sodio y potasio.
|
|
ATPasa de Ca2+
|
De esta forma, mediante varios mecanismos las células se valen de componentes enclavados en sus membranas, y pueden realizar el movimiento de diferentes sustancias, generalmente iones, y ello les permite establecer una liga efectiva entre sus procesos metabólicos y el transporte. En esencia, los mecanismos de transporte que se han señalado hasta ahora no significan otra cosa que la conversión de la energía metabólica o hasta la luminosa en otro tipo: diferencias de concentración a través de las membranas, que luego pueden ser aprovechadas para mover a otras sustancias o con otros fines. Nuevamente es necesario no olvidar que las diferencias de concentración de las sustancias a ambos lados de una membrana representan una forma eficaz de energía, que puede ser utilizada para muchas otras funciones.
Hemos considerado la posibilidad de que se movilicen sustancias, y hemos hablado de la energía que esos movimientos implican, porque éstas tienen una concentración en un lado de la membrana y otra en el opuesto. Sin embargo, como en los casos que acabamos de revisar, además de existir una diferencia de concentración de una sustancia cualquiera, resulta que ésta tiene una carga eléctrica; de inmediato es claro que no es lo mismo tener una diferente concentración de glucosa a ambos lados de una membrana, que tener la misma diferencia de concentración de Na+. Aunque en ambos casos esta diferencia de concentración significa también una diferencia de energía en ambos lados, en el caso del ion sodio hay además una diferencia en la cantidad de cargas y esto significa un componente adicional en la energía del sistema: la energía eléctrica. De esta forma, al hablar del movimiento de sustancias con carga eléctrica, hay que considerar dos elementos desde el punto de vista de lo que eso significa: la diferencia de concentración y la diferencia de la cantidad de cargas a ambos lados de la membrana. Para que se desplace una partícula cargada, ésta debe estar influida por la diferencia de cargas que casi siempre existe en ambos lados de las membranas y viceversa, al moverse modifica el estado de equilibrio que existía.
Es así como se entiende la relación entre la energía y el transporte. Las diferencias de concentración de las sustancias representan estados energizados, de la misma forma que hay energía en el aire de una llanta que se encuentra a mucha mayor presión que la del medio ambiente, o en el agua de un tanque que se encuentra en el techo de un edificio. Para llegar a obtener las diferencias de concentración se requiere de energía, y estas diferencias alcanzadas representan otra forma de energía en la que la primera se ha convertido, compuesta una parte por las diferencias de concentración, y la otra por las diferencias en la cantidad de cargas eléctricas. Para todas las sustancias que existen a un lado y al otro de una membrana, esta suma es lo que se llama el potencial electroquímico.
LOS TRANSLOCADORES SECUNDARIOS
A diferencia de los translocadores primarios, que están directamente conectados con las fuentes de energía de las células y son capaces de mover iones utilizándola directamente, hay otra especie, la de los translocadores secundarios, que no tienen, por así decirlo, acceso directo a las fuentes de energía, y desplazan sustancias a través de las membranas por otros medios; el caso más sencillo de éstos es tal vez el de un acarreador que simplemente puede reconocer a una molécula y permitirle el paso de un lado al otro de la membrana, de acuerdo con la diferencia de su concentración en ambos lados. En realidad, éste es casi un caso de difusión, pero como no ocurre de manera libre a través de la membrana, se le llama de difusión facilitada. Así ocurre el transporte de glucosa en algunas levaduras.
En estas células, la glucosa se transporta sencillamente de un lado al otro de la membrana, facilitada por la presencia del acarreador; según su diferencia de concentración la glucosa penetra si está más concentrada en el exterior que en su interior, pero puede darse el caso de que la concentración en el interior sea superior a la de afuera; en estas condiciones, el azúcar sale de la célula. Existe un sistema de transporte semejante para la glucosa en las levaduras, que habitualmente se utilizan para fermentar medios, como el mosto en la producción del vino, que contienen altas concentraciones de azúcar. Este es un caso de transporte de acuerdo con el gradiente o diferencia de concentración, y, por su semejanza con la difusión, se le llama difusión facilitada; es decir, la sustancia ve favorecida su entrada (o salida) por la presencia del acarreador en la membrana, pero se desplaza en realidad por las condiciones energéticas propias, que derivan de sus diferencias de concentración en ambos lados de la membrana.
Hay otro caso que no es tan sencillo, pero semejante al anterior, y que se
refiere al movimiento de una sustancia, aparentemente en contra de su tendencia
natural, con menor concentración de un lado de la membrana, y con una mayor
concentración del otro. Este tipo de sistemas se encuentra en muchos seres vivos,
y como ejemplo se observa en las mitocondrias de algunas células: el Ca2+
puede penetrar a las mitocondrias, aun cuando en el interior se
encuentre más concentrado, de acuerdo con el mecanismo que se representa en
la Figura 28. La mitocondria tiene un sistema para bombear protones o hidrogeniones
(H+) al exterior, y ello resulta de la generación
de un potencial eléctrico, negativo en el interior. El Ca2+,
un catión o ion con carga positiva, se ve atraído al interior de la mitocondria,
de tal manera que con la sola presencia de un sistema de transporte que facilite
su entrada, puede traspasar la membrana y acumularse con una mayor concentración
que afuera. Este es un sistema de transporte que se parece muchísimo a la valinomicina,
sólo que utiliza otro ion. También se representa en la Figura 28 el caso semejante
del sistema para acumular potasio en la levadura. Estos dos sistemas descritos
son lo que se conoce como uniportadores, es decir, sustancias que trasladan
una sola sustancia o ion de un lado al otro de la membrana a la vez.
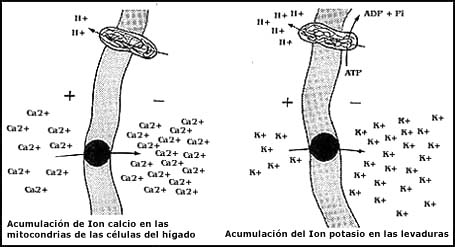
Figura 28. Uniportadores que tienen la capacidad de acumular iones en un lado de una membrana, aprovechando una diferencia de potencial eléctrico. Los iones se acumulan impulsados por la diferencia de carga que una ATPasa o "bomba", ha generado previamente. El fenómeno es semejante a la difusión facilitada, pero en él influye además una carga del ion y la diferencia de potencial a los lados de la memebrana.
Hay otras sustancias que se pueden acumular en las células gracias a la existencia
de otros sistemas de transportadores secundarios: los simportadores, o cotransportadores,
como el que se representa en la Figura 29. Se muestra como ejemplo el caso de
la acumulación de glucosa por las células intestinales, aun en contra de su
diferencia de concentración, pero aprovechando que en el intestino hay una concentración
mayor de Na+ que dentro de las células. Esta
diferencia de concentración proviene del funcionamiento de un translocador primario
que expulsa al sodio al exterior. Habiendo así una mayor concentración de sodio
en el exterior, éste muestra una tendencia natural a entrar; hay entonces un
sistema de transporte que la aprovecha, pero obligándolo a entrar sólo si se
acompaña de una molécula de glucosa; de esta forma, se logra que la molécula
no sólo penetre, sino que se pueda aprovechar y acumular, aun cuando las concentraciones
exteriores sean muy bajas, o al menos menores que las del interior de las células.
Hay un sistema semejante de transporte que aprovecha en la levadura y en la
mitocondria, por ejemplo, la diferencia de concentración de H+,
para introducir un fosfato. También aquí se aprovecha la tendencia natural del
hidrogenión (H+) a entrar, para obligarlo a
que, en cierta forma, empuje con él a un fosfato.
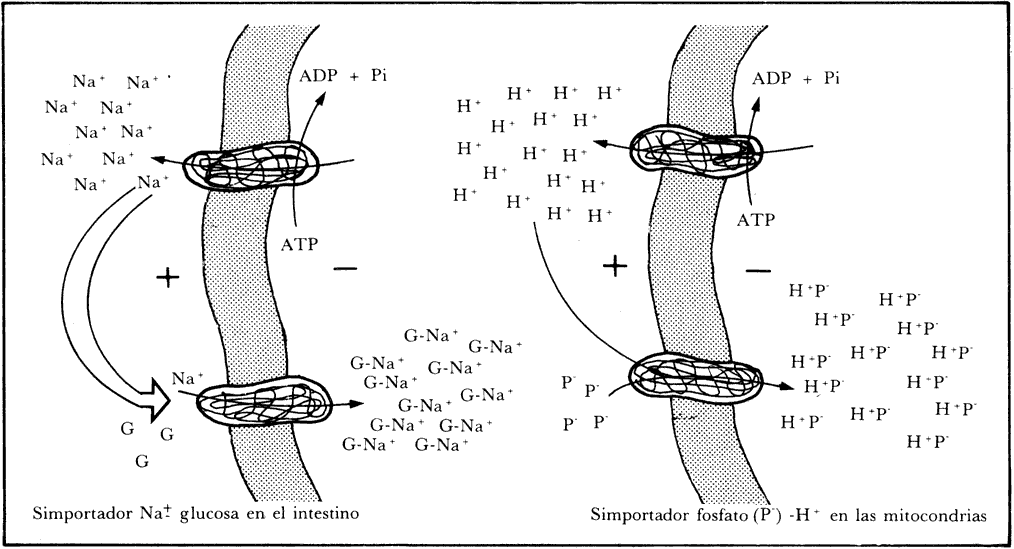
Figura 29. Simportadores. Son sistemas de transporte que permiten a las
células concentrar algunos materiales, aprovechando las diferencias de concentración
que hay de otros.
Hay todavía otro tipo de transportadores secundarios llamados antiportadores o intercambiadores; en el caso de éstos, una sustancia también se puede acumular dentro de una célula o una partícula subcelular. El fosfato puede acumularse en una mitocondria por el mecanismo que ya señalamos en el párrafo anterior; pero ahora, la mitocondria tiene la necesidad de acumular otra sustancia para su metabolismo llamada malato; para ello hay un sistema de transporte capaz de intercambiar un fosfato de adentro por un malato de afuera, y de esta forma satisfacer la necesidad del segundo. En las mitocondrias de algunas células en especial ha sido posible describir lo que se llama una "cascada" de intercambios, en la cual se acumula primero fosfato, que luego se puede intercambiar por malato, y después éste se puede intercambiar por citrato.
Los sistemas de transporte así organizados se vuelven extraordinariamente versátiles. No es necesario que las células tengan un translocador primario conectado a las fuentes de energía en forma directa para acumular a cada uno de ellos. Basta con uno de estos translocadores primarios para mover a muchas sustancias e inclusive acumularlas.
Por otra parte, la acumulación de algunos iones, de los cuales el más importante
es el potasio, sirve a las células como una forma de energía aprovechable especialmente
para el transporte. Las células animales, por ejemplo, tienen la llamada bomba
de sodio-potasio, que funciona como se señala en la Figura 30. Utilizando la
energía del ATP, puede expulsar tres átomos de sodio y acumular
dos de potasio. De esta forma, por un lado cuentan con un gradiente de concentración
de sodio y otro de potasio, y además un potencial negativo en el interior. Todos
estos componentes que resultan de la operación de un solo translocador primario
tienen luego importancia extraordinaria, no sólo en los fenómenos de transporte
como tales, sino en un buen número de funciones celulares, algunas de ellas
muy especializadas en algunos casos (véase más adelante).
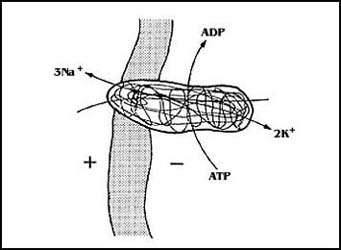
Figura 30. La bomba de Na+ y K+.
Esta enzima, aparentemente tan sencilla, es de gran importancia para las células.
Se le ha encontrado en todos los tejidos animales y es responsable de numerosas
capacidades adicionales de transporte, además de que mantiene la elevada concentración
de K+ que las células requieren en su interior
para funcionar adecuadamente.
Este sistema de transporte es esencial para el buen funcionamiento de órganos tan importantes como el sistema nervioso central y el corazón. En este último órgano se encontró hace muchísimo tiempo que los extractos de algunas plantas del género Digitalis mejoraban el estado de los pacientes con insuficiencia cardiaca. Al estudiar esta enzima hace algunos decenios, se encontró que las sustancias contenidas en estos extractos ejercen su efecto curativo inhibiendo a la enzima. Parece que esta inhibición altera las condiciones de las células dañadas en tal forma que les permite recuperar la capacidad de contracción.