II. RECEPTORES: LOS OÍDOS DE LAS CÉLULAS:
EN EL CAPíTULO ANTERIOR se menciona que las hormonas viajan para interactuar con sus células blanco. Esto implica que sólo algunas y, por lo tanto, no todas las células del organismo son receptoras del mensaje. Por otro lado, el mensaje se concentra en estas células receptoras. Se han hecho experimentos en los que se administra una hormona marcada con radiactividad a un animal de experimentación y se observa que dicha radiactividad se concentra en las células blanco. ¿A qué se debe todo esto? Una de las razones más importantes es la presencia de receptores. Pero, ¿qué es un receptor? Básicamente, es una estructura química (proteína) capaz de recibir al mensajero y de transmitir el mensaje para que se produzca la respuesta de la célula. Es decir, el receptor como tal tiene dos características fundamentales: 1) reconocer al mensajero para interactuar con él y 2) activar la secuencia de eventos que conducen a la respuesta celular.
En 1906 Langley estaba estudiando los efectos de dos sustancias, la nicotina y el curare, en una preparación experimental con células de músculo y nervio. Este investigador observó que dichas sustancias competían entre sí, lo que lo llevó a concluir que "el mutuo antagonismo del curare y la nicotina sobre el músculo sólo se puede explicar satisfactoriamente si se supone que ambos se combinan con una misma sustancia receptora, la cual recibe el estímulo, y al transmitirlo causa la contracción del músculo". Hacia 1913 Paul Ehrlich formula su postulado clásico corpora non agunt nisi fixata: las sustancias no actúan a menos de que se fijen. Estos conceptos sientan las bases de mucho de lo que se sabe hoy día sobre la acción hormonal.
La idea de "receptor" permaneció como tal, es decir, como un concepto abstracto sin pruebas experimentales directas, por muchos años, pero recientemente se ha materializado. En la actualidad ya se conoce la naturaleza química de muchos receptores, se han purificado varios de ellos, se han reconstituido no pocos (esto es, incorporado a sistemas artificiales donde se puede reproducir alguna de sus funciones en la célula), y un buen número ya ha sido sintetizado en el laboratorio con métodos semiartificiales, lo cual ha obligado a aquellas células que normalmente no sintetizan a algún receptor a hacerlo. Así por ejemplo, se ha podido incorporar la información genética para que algunas células de ratón sean capaces de expresar receptores humanos para algunas hormonas. ¡Increíble!, ¿no les parece? ya es posible purificar e incluso sintetizar a estos "oídos" de la célula y, seguramente, lo mejor está aún por venir.
Pero volvamos al concepto de receptor. Los receptores son proteínas grandes, de peso molecular elevado. Como todas las proteínas, la información para su síntesis se encuentra almacenada en el material genético de cada célula (ADN). De tal suerte, que en la célula que nos dio origen ya estaba almacenada la información para la síntesis de los receptores para todos los mensajeros con los que se comunican nuestras células. Por supuesto, éstas al irse diferenciando, es decir, convirtiendo en células del cerebro, hígado u otro órgano, van expresando los receptores que necesitan, en el momento y en las cantidades que se requieren.
Vale la pena hacer algunas breves consideraciones. Nosotros somos el resultado de un complejísimo proceso evolutivo. Podríamos pensar que hace millones de años surgió la necesidad de comunicarse entre las primeras células de nuestro planeta. Allí surgieron los primeros mensajeros y los primeros receptores. A través de modificaciones en el material genético (mutaciones), nuestras proteínas en general y entre ellas nuestros receptores, han ido evolucionando durante millones de años, y continuarán haciéndolo. Así, algunas células multiplicaron la información genética para estos receptores y dicha información se modificó, ahora tenemos receptores discretamente diferentes; con ello se adquirió, poco a poco, la capacidad de responder a diferentes mensajeros. Al irse diferenciando se formaron familias de receptores, y de estas familias nuevas familias y nuevas familias hasta constituir un enorme árbol familiar para los receptores. En estos momentos, y gracias al avance de la bioquímica y la biología molecular, empezamos a conocer partes de ese árbol y las relaciones que existen entre sus componentes.
Volvamos a los receptores. Éstos, como todas la proteínas, tienen estructura tridimensional, es decir, una forma determinada en el espacio. Existen receptores con muy diversas formas, se pueden imaginar como estructuras más o menos semejantes a un balón, o bien, alargadas como una salchicha; pensemos, además, que su superficie no es tersa sino llena de irregularidades, donde en alguna parte, está precisamente el sitio de reconocimiento al que se une el mensajero. Las superficies del mensajero y del receptor se adaptan perfectamente entre sí. Podría decirse que es como el acoplamiento entre dos naves espaciales; como la interacción entre una llave y su chapa, o bien, como un guante y una mano. Esta perfecta adaptación de superficies es la base de la alta selectividad de los receptores para una hormona específica. Imaginemos al receptor no como una estructura rígida, sino como una estructura con cierta flexibilidad, capaz de sufrir ciertos cambios en su forma. El sitio de reconocimiento también tiene cierta flexibilidad, lo cual nos lleva al concepto de afinidad.
La afinidad puede definirse como una medida de la facilidad de interacción entre dos sustancias, en este caso entre el receptor y el mensajero. Esto es similar a lo que ocurre entre los seres humanos; dos individuos que tienen que interactuar para la realización de un trabajo lo harán con facilidad si existe afinidad entre ellos y lo efectuarán con extrema dificultad si no son afines.
Véase el siguiente experimento: en una preparación se tienen 100 receptores; para ocupar la mitad deben agregarse 50 unidades de hormona A; para hacer lo mismo, pero ahora con la hormona B, tienen que agregarse 50 000 unidades de esta sustancia; si ahora se hace con la hormona C, tendrán que agregarse 50 millones de ella. Esto quiere decir que tanto A como B o C pueden interactuar con el receptor; pero este receptor prefiere 1 000 veces a A que a B y un millón de veces a A que a C. El receptor nos está mostrando su capacidad para seleccionar a un mensajero, pero si se fuerzan las condiciones se puede lograr que mensajeros con baja afinidad por un receptor interactúen con él. Es como si se metiera por la fuerza una mano grande en un guante pequeño; desde luego que se puede hacer, pero cuesta más trabajo.
Esto tiene importancia por varias razones. En algunas enfermedades las concentraciones de una hormona pueden aumentar tanto, que llegan a observarse efectos que ésta ejerce interactuando con receptores para otras hormonas discretamente diferentes en su estructura. Además, nos ilustra sobre la necesidad de que, cuando los médicos receten compuestos con efectos hormonales, ajusten las dosis con una gran precisión.
La mayoría de las sustancias que se recetan y que actúan sobre los sistemas
de comunicación celular han sido diseñadas para tener una altísima selectividad
por un solo receptor; muchas veces mayor que la del mensajero natural. Sin embargo,
hay algunos compuestos que pueden interaccionar, y de hecho lo hacen, en concentraciones
similares con varios tipos de receptores; a estos agentes los llamamos "promiscuos"
y casi siempre tienen más acciones indeseables que de utilidad. Existe una caricatura
en la cual el médico le dice al paciente:
—tómese esta píldora cada ocho horas; ahora bien, para contrarrestar su
efecto sobre el pulmón, tome este jarabe con cada comida; si le da diarrea,
tome estas pastillas, y si.... Poca selectividad, ¿no creen?
Permítanme contarles la no tan triste historia del honorable doctor don Farmacón Forte. Este sabio investigador dedicó los primeros 20 años de su vida profesional a estudiar la química de un mensajero. Una vez alcanzado su objetivo, se enfrascó los siguientes 30 años en diseñar y sintetizar en su laboratorio un compuesto que se uniera con el receptor para ese mensajero, con 1 000 veces mayor afinidad que el mensajero natural; es decir, del que se requiriera administrar 1 000 veces menos moléculas que del mensajero natural para obtener la misma respuesta de la célula. ¡Realmente había que sintetizar a este mensajero prodigioso! Una vez lograda la síntesis, y después de penosas y explosivas experiencias (entiéndase, después de algunas explosiones), decidió hacer el experimento crucial y administrar una pequeña cantidad de su compuesto a unas células. Sorprendentemente nada pasó... "Quizá fue demasiado poco, no es tan potente como yo creía" —pensó don Farmacón— y agregó una buena dosis complementaria (digamos medio frasco, por aquello de no quedarse corto). Pero, para su gran desaliento y frustración, nada sucedió. Las células seguían como si nada hubiera ocurrido. Don Farmacón se sintió desolado; colgó su bata y se retiró derrotado a casa. Por su mente pasaban todos los minutos de esos 50 años invertidos en el supermensajero. A pesar de todo, comió (pues no hay que exagerar) y durmió. Durante el sueño recordó sus cursos de "receptorología avanzada", y el postulado sapientísimo "lo que no estimula, inhibe" resonaba en su cabeza. Despertó iluminado, ¡todo estaba aclarado! Regresó al laboratorio donde repitió el experimento y observó que, efectivamente, el compuesto que había sintetizado no tenía efecto por sí mismo, pero bloqueaba la acción del mensajero natural, y a concentraciones bajísimas. Preparó una comunicación para la "tres veces H" sociedad científica, "Royal and Democratic Tingüindinus Society", donde hacía ver, con toda veracidad cómo desde hacía 50 años estaba tratando de sintetizar un antagonista del mensajero natural y cómo, finalmente, su preclaro pensamiento y su notable esfuerzo lo habían llevado a alcanzar el tan deseado éxito.
El relato anterior ilustra dos conceptos importantes de la comunicación celular: la afinidad y la actividad. La afinidad es una medida de la facilidad de acoplamiento entre el mensajero y el receptor. La actividad es la capacidad del mensajero para producir el efecto. A un compuesto que es capaz de unirse con el receptor y producir una respuesta en la célula, o sea, un efecto, se le llama agonista. Un antagonista (o anti-agonista) es aquella sustancia que, por sí misma, no produce efecto en la célula (no tiene actividad), pero que es capaz de interactuar con el receptor, ya que sí tiene una buena afinidad por él. Al asociarse con el receptor ocupa el sitio que pudiera ocupar el mensajero natural u otro agonista; esto es, inhibe o antagoniza el acoplamiento mensajero-receptor y así bloquea el efecto. A muchos de nosotros, cuando tenemos alguna reacción alérgica, el médico nos receta un antihistamínico. En muchas reacciones alérgicas participa el mensajero histamina y lo que nos recetan es un antagonista; es decir una sustancia que por sí misma no active a los receptores para la histamina, pero que pueda interactuar con estos receptores y bloquear el efecto de la histamina que se está produciendo en nuestro organismo.
Se desconoce qué hace que, de dos sustancias capaces de unirse con un mismo receptor, una sea agonista y la otra antagonista. Sin embargo, alguna información se desprende del campo experimental. Una primera conclusión es que el simple acoplamiento con el receptor es insuficiente para desencadenar la respuesta de la célula; lo cual significa que el agonista "activa" al receptor, mientras que el antagonista no lo hace. Pero, ¿qué significa "activar" al receptor? Posiblemente inducir un cambio tal en su estructura, que permita que el receptor, ya activado, interactúe ahora con el siguiente elemento en la transmisión del mensaje. Múltiples estudios se han realizado para determinar qué partes de las estructuras químicas de los diferentes mensajeros (hormonas, neurotransmisores, etc.) son importantes para determinar su afinidad y cuáles para definir su actividad. Esta es la principal tarea de investigación de buena parte de la industria farmacéutica y de muchos investigadores interesados en la relación estructura-actividad. Por otro lado también se están localizando, dentro de la estructura química de algunos receptores, los sitios específicos donde interactúan tanto el mensajero natural como los agentes sintéticos agonistas o antagonistas.
Recientemente se ha aclarado que muchos receptores, aun sin ser estimulados por agonistas, tienen cierta actividad basal. Es decir, existe un cierto "ruido de fondo". Algunos agentes, considerados anteriormente como simples antagonistas, tienen la capacidad no sólo de evitar que el receptor se active por los agonistas, sino de disminuir la actividad basal de los receptores. A estos agentes se les ha dado el nombre de agonistas inversos o superantagonistas.
D) ¿DÓNDE INTERACTÚAN LOS MENSAJEROS CON LOS RECEPTORES?
Las hormonas constituyen un conjunto bastante heterogéneo de sustancias que pueden dividirse en muchos grupos químicos. Afortunadamente, desde el punto de vista de su acoplamiento con los receptores la situación es más sencilla; a las hormonas las dividimos en dos grandes grupos: aquéllas en las cuales el acoplamiento con el receptor se lleva a cabo en el interior de la célula, y aquéllas en las que se efectúa en el exterior de la misma. Se pudiera decir entonces que existen dos tipos básicos de "oídos celulares": los internos y los externos.
Se hablará primero de los receptores externos. Pediré al lector que imagine que se va reduciendo de tamaño en forma progresiva, o que cuenta con un microscopio potentísimo que le permite ver lo minúsculo como enorme. Así logrará imaginar a los glóbulos rojos como grandes botes de hule inflable, con sus bordes abultados y sus centros hundidos; verá cómo estas células enormes se golpean al circular por los vasos sanguíneos y cómo sus paredes son sumamente elásticas, como si fueran globos. Ahora nos haremos mucho más pequeños aún, o cambiaremos las lentes de nuestro microscopio por otras más potentes. ¡Qué diferencia! Entramos como en otro mundo. El glóbulo rojo es ahora tan grande como si fuese el mar; vemos dónde comienza pero no dónde termina. Su superficie, que antes nos parecía brillante y tersa, ya no es tan brillante y es mucho menos tersa. Si observamos con cuidado veremos muchas estructuras en su superficie, es decir, en esta ahora inmensa membrana. Enormes pelotas, como montañas que deambulan a semejanza de los icebergs, se encuentran como suspendidas en la membrana de la célula. Algunas de estas montañas son los receptores que se asoman en la membrana externa o plasmática de la célula. La membrana está constituida por la unión de dos capas delgadísimas de lípidos que contienen en su seno múltiples proteínas. Imaginemos, pues, esta "piel" de la célula como ese mar con múltiples icebergs inmersos y otras estructuras flotando en su superficie: las proteínas de la membrana. La membrana no es una entidad rígida; todo lo contrario, está en continuo movimiento y cambio, como el mar. Las proteínas de la membrana pueden atravesarla totalmente, desde su parte externa hasta el interior de la célula (de hecho, la pueden atravesar en múltiples ocasiones, entrando y saliendo de ella), o bien permanecer orientadas hacia fuera o hacia dentro.
Los receptores externos tienen una cara hacia el exterior de la célula, una parte de su estructura embebida en la membrana plasmática y otra porción o cara mirando hacia el interior de la célula. La cara exterior contiene el sitio de reconocimiento para el mensajero; el resto del receptor sirve para procesar y transmitir la información a la célula. La presencia del receptor en la cara externa de la membrana plasmática hace innecesario que la hormona o neurotransmisor tenga que penetrar al interior de la célula para producir el efecto, ya que como hemos visto puede hacerlo desde fuera.
Pero, ¿cómo sabemos que esto sucede realmente así?, ¿cuáles son las evidencias para afirmarlo? Hay una gran cantidad de experimentos que apoyan estas afirmaciones. Se señalarán algunos de ellos. Con el uso de técnicas relativamente simples en el laboratorio se pueden romper las células y separar sus componentes: el líquido intracelular o citosol, los núcleos, las mitocondrias, e incluso la membrana plasmática. Utilizando una hormona, marcada con radiactividad, por ejemplo, se puede determinar cuanta de ésta se acopla a su receptor en las diferentes fracciones de las células y así darnos una idea de dónde se localiza la mayor concentración de radiactividad y, por ende, de receptores. Por medio de estas técnicas se ha podido determinar que la membrana plasmática está enriquecida de receptores; esto es, hay receptores en otros tipos de membranas (ya se verá más adelante por qué están allí), pero que en general los receptores se localizan preferentemente en la membrana plasmática.
El experimento anterior nos habla de la localización de los receptores en la membrana plasmática, pero no nos dice nada acerca de que estos receptores sean los que ejerzan la acción. Se puede hacer otro experimento para probar esto último; pensemos en una sustancia que destruya las proteínas (incluyendo a los receptores) que se encuentran en la superficie exterior de las membranas, pero que no penetre al interior de las células. Algunas proteasas (enzimas que rompen proteínas) pueden ser utilizadas con este fin. Son como leñadores que cortan todo lo que sobresalga de la superficie de la membrana; la dejan "rasurada".
Supóngase ahora que "se rasura" una célula y luego se la pone en contacto con el mensajero. Si la célula no responde (como de hecho sucede), indica que algo que se ha quitado de su superficie provoca que no "oiga" el mensaje. Por lo tanto, el mensajero interactúa con "algo" que se encuentra en el exterior de la membrana. Este "algo" son los receptores.
Otra forma ingeniosa de demostrar que la interacción entre la hormona y el receptor ocurre en el exterior de la célula es la siguiente: unir el mensajero a una partícula de tamaño tan grande, que le sea imposible penetrar a la célula; si ésta, aun así, responde al mensaje, querrá decir, sin lugar a dudas, que la interacción hormona-receptor es externa. Todos estos experimentos se han realizado con algunas hormonas e indican: a) la localización externa del receptor, y b) que la interacción adecuada para que se produzca el efecto es, precisamente, la que se realiza con el receptor localizado en la cara externa de la membrana plasmática. Ahora bien, ¿se pueden emplear estas pruebas con los receptores internos? Cuando los receptores son intracelulares todos los trucos anteriores no dan resultado. En estos casos, el mensaje debe atravesar la membrana plasmática para ejercer su efecto.
En los fenómenos de comunicación celular, un parámetro de gran importancia es el tiempo. Hay efectos que son casi inmediatos, mientras que otros tardan en producirse horas o días. Entre los primeros se pueden mencionar, como ejemplos, el aumento en la frecuencia cardiaca ante un peligro o emoción intensa; la sudoración o la sensación de sequedad de la boca ante un examen u otra calamidad semejante; y miles menos perceptibles. Y, como acciones a largo plazo, las relacionadas con el crecimiento y el desarrollo. En general, las acciones hormonales que ocurren en forma casi inmediata (segundos o minutos) involucran a receptores externos; las acciones a largo plazo pueden involucrar tanto a receptores externos como internos, pero casi siempre están asociadas a la síntesis de nuevas proteínas. Aclararé esto último más adelante.
E) ¿HAY UN RECEPTOR PARA CADA HORMONA?
Ya se mencionó que la información para la síntesis de todos los receptores hormonales de nuestras células se encuentra almacenada en el ADN de la célula que nos dio origen: conforme éstas se van diferenciando, van expresando (sintetizando) los receptores para las hormonas que regularán su funcionamiento, es decir, de las cuales van a ser blanco. Como resulta obvio, las células no expresan la totalidad de los receptores para los cuales tienen información, sino que van haciendo una expresión selectiva de aquellos receptores que necesitarán para realizar sus funciones, de acuerdo a su programa de diferenciación.
Ahora bien, ¿hay un receptor para cada mensajero, o varias hormonas comparten un mismo receptor? La evidencia que se tiene hasta el momento nos indica que hay gran especificidad en los receptores y que existe, por lo menos, un receptor específico para cada mensajero. ¿Cómo que por lo menos? Sí, por lo menos. Hay muchos casos en que existen varios tipos de receptores para un solo mensajero. Esto parece ser especialmente claro en el caso de los receptores para hormonas y neurotransmisores cuya estructura química es más simple: acetilcolina, adrenalina, histamina, etc. Vale la pena mencionar que, para la adrenalina, cada uno de nosotros cuenta con por lo menos nueve tipos de receptores (tres subtipos de receptores de cada una de tres familias de receptores adrenérgicos); para la acetilcolina hay dos familias, una de ellas con cinco subtipos, para la histamina hay tres subtipos, y.... seguimos contando..., pues el número total de receptores aún no ha sido definido. Ciertamente hay mucho que hacer. Esto pudiera parecer una excesiva complicación, pero en realidad no es así, ya que brinda a las células una mayor capacidad para regular sus funciones, además de que tiene una gran importancia, tanto en el funcionamiento normal del organismo, como para el tratamiento de diversas alteraciones.
F)¿PUEDE UNA CÉLULA TENER VARIOS TIPOS DE RECEPTORES PARA LA MISMA
HORMONA?
A través de los años se ha reconocido que algunos tejidos tienen grandes cantidades de un tipo de receptor en particular; de hecho, aun se llegó a decir que un mensajero es "selectivo" o exclusivo para un órgano, con base en su alta densidad de un tipo de receptores. Conforme avanzan los años, las técnicas se perfeccionan y lo que no se veía anteriormente ahora se empieza a distinguir. Estudios más específicos han demostrado que ciertamente hay varios tipos de receptores para una misma hormona en un mismo tejido y que no existe tal absoluta especificidad de tejido para una hormona.
Evidencias recientes indican que en un solo tipo de célula pueden expresarse varios de los subtipos que existen para un receptor y que estos receptores pueden llevar un mensaje diferente a la célula. ¡Qué barbaridad! ¡como si fueran pocos los mensajeros, ahora además resulta que cada mensajero puede tener diferentes tipos de receptores y que cada subtipo de receptores puede llevar una información distinta a la célula! En realidad esto es lo más atractivo de la regulación hormonal; con cada descubrimiento se vislumbran nuevos mecanismos de adaptación celular. Una de las características de este tipo de conocimientos es que permite definir formas en que, potencialmente, se puede influir en las respuestas celulares. Esto tiene implicaciones significativas para el tratamiento de algunas enfermedades.
G) ¿VARÍAN EL NÚMERO Y EL TIPO DE RECEPTORES EN LAS CÉLULAS EN EL
CURSO DE LA VIDA?
El número y el tipo de receptores, que expresa una célula, puede cambiar de acuerdo a las circunstancias. A continuación se presentan dos ejemplos de este fenómeno.
Las hormonas producidas por la glándula tiroides regulan la respuesta del corazón a la adrenalina. Así, se sabe que en personas normales la administración de una pequeña cantidad de adrenalina puede aumentar la frecuencia cardiaca en digamos los latidos por minuto; en personas que tengan una secreción anormalmente elevada de hormonas tiroideas (hipertiroideas), la misma dosis de adrenalina produce una respuesta cinco o seis veces mayor. Por el contrario, en personas con una producción deficiente de ellas (hipotiroideas), se produce una muy escasa o nula respuesta. Con el uso de modelos experimentales se ha logrado aclarar, en gran parte, a qué se debe este fenómeno. El número de un subtipo de receptores para la adrenalina disminuye en animales hipotiroideos y aumenta en hipertiroideos. En otras palabras, el incremento en la respuesta a la adrenalina que se observa en las células cardiacas de pacientes hipertiroideos se debe, en gran parte, a un aumento en el número de un tipo de receptor para esta hormona en el corazón, y en el caso de los hipotiroideos a una disminución en el número de este tipo de receptores. Así, en estas alteraciones tenemos una célula muscular cardiaca que escucha demasiado (hipertiroidismo) y otra casi sorda (hipotiroidismo) para la acción de la adrenalina, respectivamente. Es interesante mencionar, además, que el número de receptores para la adrenalina en el corazón vuelve a la normalidad al corregirse el problema tiroideo; lo que nos habla de la enorme plasticidad del sistema.
Estudios recientes realizados en mi laboratorio, y en otros, han demostrado otro caso muy interesante. El tipo de receptores para una hormona en un tejido dado puede variar conforme a la edad del animal. Vayamos al ejemplo: la respuesta a la adrenalina en el hígado de la rata en la etapa fetal y en la recién nacida es mediada por un tipo de receptor: el B2-adrenérgico. Conforme la rata crece y madura, el receptor expresado cambia de tal forma que, en la rata joven, la célula hepática expresa tanto receptores B2 como receptores a1B-adrenérgicos y la acción de la hormona está mediada por ambos. En la rata adulta, la acción de la adrenalina está mediada sólo por receptores a1B-adrenérgicos. Es decir, durante el desarrollo, la célula hepática va cambiando la expresión del tipo de receptor que utiliza para escuchar al mensajero adrenalina, pasando de B2 a 1B.
Desafortunadamente, no se cuenta todavía con un elixir rejuvenecedor. Ante este infortunado hecho, se tuvo que realizar un rejuvenecimiento parcial (¿?). El hígado de la rata es un órgano notabilísimo, tanto por sus casi innumerables funciones, como por algunas de sus particularidades celulares. Una de ellas es su enorme capacidad de regeneración. Si se extirpa el 80% del hígado a una rata, tres días después de la operación el 20% que se dejó se ha multiplicado, observándose que el nuevo hígado pesa aproximadamente ¡entre 80 y 90% del original! ¡Qué maravilla!, ¿no les parece? Las células del hígado, que normalmente se reproducen muy poco, ante un estímulo como la extirpación quirúrgica se reproducen activamente hasta regenerar el órgano.
Bueno, como se explicó anteriormente, dado que no se puede rejuvenecer a toda una rata, se le rejuvenece el hígado: se realizó la operación a ratas adultas lográndose, como se esperaba, que el 20% del hígado original restante nos formara un nuevo órgano. Estudiamos entonces el tipo de receptor que mediaba las acciones de la adrenalina en este nuevo hígado y... ¿qué creen ustedes?, tres días después de la operación el tipo de receptores era B2. ¡Se había logrado rejuvenecer la respuesta adrenérgica hepática! Sin embargo, siete días después de la operación, la respuesta a la adrenalina del hígado neoformado volvía a ser mediada por receptores a1B-adrenérgicos. Es decir, las células "envejecieron" rápidamente; el proceso de maduración de la respuesta hepática, que toma muchos meses en la rata, se había reproducido en sólo siete días. Nótese la gran plasticidad de las células y obsérvese cómo el funcionamiento del sistema no es tan sencillo como lo imaginamos en un principio: se secreta un mensajero y se produce una respuesta. Nuestras células cambian en su capacidad para responder a las hormonas y en el tipo de sistemas o receptores que son activados por éstas. Todo ello ocurre sin que nos percatemos en lo más mínimo.
Las células están en una continua adaptación a las condiciones cambiantes que las rodean; ya sean células aisladas como los microorganismos o células de un tejido en un ser complejo. Anteriormente hablábamos de que esta adaptabilidad es precisamente la que les permite continuar viviendo. Parte importante de ella es las modulación de la respuesta celular a estos estímulos extracelulares que llamamos mensajeros. Se ha visto, en uno de los ejemplos anteriores, cómo algunas hormonas modulan la respuesta de las células a otros mensajeros. Así pues, las interacciones son múltiples y muy complejas: un mensajero no sólo produce una respuesta específica, sino que, al mismo tiempo, puede regular la respuesta a otros agentes.
H) ¿PERMANECEN ESTÁTICOS LOS RECEPTORES? O "VEINTE MIL LEGUAS DE VIAJE
CELULAR"
Los receptores tienen una movilidad extraordinaria dentro de la membrana celular; efectúan movimientos en varios sentidos: horizontal y vertical. Se agrupan; pasan a otras membranas en el interior de la célula; se reciclan, es decir, regresan a la membrana plasmática, etcétera.
A continuación se explicará algo de lo que se sabe acerca de este movimiento de los receptores y de sus "misteriosos viajes" a través de la célula; pero antes se aclarará algo sobre la membrana.
Unas páginas atrás se mencionó que la membrana plasmática es una estructura sumamente dinámica (con amplio movimiento) sujeta a un permanente cambio de sus elementos. Este cambio no sólo implica una nueva localización física de dichos elementos sino un recambio constante, es decir, que unos componentes son sustituidos por otros nuevos. Esta gran movilidad no debe sorprendernos. Hace muchos años se pensaba que el organismo humano tenía una fase de crecimiento en la que se formaban sus partes, y que luego, en la etapa adulta, estos elementos ya formados permanecían estables por mucho tiempo hasta que empezaban a destruirse durante el envejecimiento. Esto, que parecería ser verdad e incluso evidente (pues creemos verlo), no es cierto. Fueron dos investigadores del noreste de Estados Unidos quienes al realizar experimentos con ratas adultas, supuestamente estables, observaron que todos los componentes químicos de estos animales se sintetizan y degradan rápidamente. Y cuando se dice todos se hace referencia a todas. De tal forma se estableció que todo nuestro organismo está en un continuo recambio: síntesis y degradación. ¿Entonces, preguntarán ustedes, por qué en algunas etapas de la vida no se aprecian cambios? Lo que sucede en estas etapas de aparente estabilidad y calma es que existe un equilibrio entre la síntesis y la degradación; es decir, se fabrica lo mismo que se destruye. De tal modo que, en el balance final, todo queda como si nada estuviera sucediendo. En los periodos de crecimiento, la síntesis supera a la degradación y, por lo tanto, el balance es positivo, o sea de ganancia para la célula o el organismo. Por el contrario, en otras etapas menos favorables (desnutrición, envejecimiento, enfermedad prolongada, etc.), la degradación supera a la síntesis, dando como resultado un balance negativo para el individuo.
Aclarado lo anterior, regresemos a la membrana. Decíamos que ésta tiene un gran recambio de sus componentes, los cuales se sintetizan y degradan con una velocidad muy alta. En el caso de los lípidos que la forman, este recambio parece tener una importancia capital en las señales hormonales. Pero esto se dejará para un capítulo posterior, y por ahora nos centraremos en los receptores que se localizan en la membrana plasmática de la célula. Estos receptores se encuentran nadando en ese mar de aceite, la bicapa lipídica, que es la membrana. Se sabe que algunos la atraviesan de un lado a otro. Dentro de ese mar, estos receptores tienen movimientos en sentido horizontal; es decir, pueden trasladarse de un sitio a otro de la superficie de la célula. Se ha observado que, bajo la acción de agentes o mensajeros que ocupan y activan a los receptores, éstos, en algunos casos, se agrupan en un punto de la célula y posteriormente se internalizan. Analicemos este fenómeno más detenidamente.
Bajo la acción de agonistas (que ocupan y "activan" a los receptores), pero no de antagonistas (que los ocupan pero no los activan), algunos receptores migran, se mudan de la membrana plasmática a membranas que se localizan en el interior de la célula. Un fenómeno muy común es que los receptores, después de ser activados y concluida (por lo menos en parte) su misión de inducir alguna o algunas respuestas en las células, se desplacen en la membrana hasta agruparse, formando zonas de alta densidad de receptores. Esto ha podido visualizarse en células vivas con ayuda de técnicas fluorescentes y microscopía de luz. También la microscopía electrónica nos ha permitido obtener una gran evidencia al respecto. Sin embargo, como se dijo antes, el fenómeno es común, pero no general; hay células para las que no se ha podido obtener evidencia de aglutinación de receptores. Pero, ya sea que se aglutinen o no, el siguiente paso parece ser la formación de una invaginación de la membrana; dicho de otro modo, la membrana se proyecta hacia el interior de la célula en una zona específica (véase la figura 3). Al aumentar de tamaño la invaginación, la membrana plasmática tiende a juntarse hasta fusionarse, formándose así una vesícula intracelular rica en receptores. Dichos receptores ya no se encuentran en la superficie de la célula, sino que han penetrado a su interior; se han internalizado (véase la figura 3). ¿Qué ocurre ahora con estos receptores? Algunas de estas vesículas ricas en receptores se fusionan con lisosomas (el aparato digestivo de las células), lo cual puede conducir a la degradación de dichos receptores, o a que sean reciclados nuevamente a la membrana plasmática por medio de un fenómeno inverso al de la internalización, y que queden listos para seguir capturando mensajeros. En el caso del receptor de la insulina se ha calculado que, en algunos tipos celulares, los receptores pueden ser internalizados y reciclados a la membrana plasmática hasta 300 veces antes de degradarse. Los receptores que se degradan, es decir, que son destruidos por las mismas células mediante sus lisosomas, se sustituyen por receptores nuevos, recién fabricados. En el caso de algunas hormonas se ha observado que la vesícula endocitótica se encuentra llena de receptores, los cuales se internalizan con todo y la hormona a la cual permanecen unidos; después de que la vesícula se ha fusionado con el lisosoma, se produce un cambio de pH (se vuelve ácido el interior de la vesícula), lo cual hace que la hormona y el receptor se separen. La hormona es usualmente destruida, mientras que el receptor puede ser reciclado.
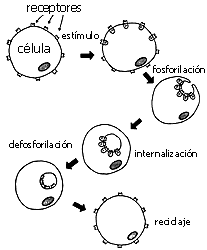
FIGURA 3. Fosforilación de receptores, internalización y reciclaje.
¿Cuál es la señal que indica a la célula que debe internalizar algún receptor? No hay aún una respuesta precisa a esta pregunta. Parece que una de estas señales podría ser la fosforilación del mismo receptor, es decir, la incorporación de una o más moléculas de fosfato por medio de enzimas específicas llamadas cinasas, de las que platicaremos más adelante. El caso más elegantemente estudiado hasta el momento es el del aceptor de la trasferina. En este sistema, se ha demostrado que si se induce la fosforilación del receptor, éste se internaliza, y que al desfosforilarse sale nuevamente a la superficie de la membrana plasmática (véase la figura 4). Si la fosforilación de receptores es una señal para la internalización del receptor, y la desfosforilación para su salida, entonces se podrán generalizar acciones en el futuro al saber cuáles agentes activan a las cinasas y cuáles a las fosfatasas (enzimas que ponen y quitan fosfatos, respectivamente), que actúan sobre determinados receptores. Sin embargo, es altamente probable que otras señales químicas determinen si el receptor se internaliza o no.
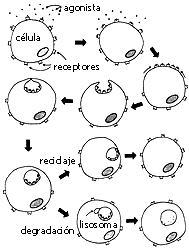
Figura 4. Internalización y procesamiento intracelular (degradación o
reciclaje) de receptores estimulados por agonistas.
Es interesante mencionar aquí un experimento reciente, realizado por el grupo de Paul Insel en California. Se sabe desde hace algún tiempo que, durante la isquemia (falta de circulación y oxigenación) del corazón, que ocurre en los infartos, aumenta la respuesta a la adrenalina por el mismo corazón; sin embargo, aún se desconocía la razón por la cual se presentaba este fenómeno. Insel y sus colaboradores demostraron que al disminuir la circulación en el corazón, los receptores para adrenalina (de tipo B1), que están en vesículas intracelulares, pasan rápidamente a la superficie de las células musculares cardiacas. Este aumento en el número de receptores, en la membrana plasmática de los miocitos cardiacos, posiblemente sea responsable de la hipersensibilidad a la adrenalina durante la falta de oxigenación. Ahora bien, ¿cuáles son los mecanismos por medio de los que se lleva a cabo? ¿Desfosforilación inducida por la falta de oxigenación? Aún no se sabe. Lo que sí se conoce es que dicha hipersensibilidad a la adrenalina está involucrada en algunas de las complicaciones agudas más graves de las crisis coronarias y de los infartos del miocardio. Evidentemente, cuando se conozca cuáles son los mecanismos que operan para que la externalización de estos receptores ocurra, se podrán diseñar tratamientos más racionales para los padecimientos mencionados.
I) RECEPTORES INTRACELULARES: "VIAJE AL CENTRO DE LA CÉLULA"
Como su nombre lo indica, estos receptores se localizan en el interior de la célula. Contrariamente a lo que pudiera pensarse, no se encuentran unidos a las membranas, sino libres, en la parte soluble del citoplasma de la célula: el citosol. Es aquí donde la interacción con la hormona tiene lugar. El receptor intracelular, al igual que el de membrana plasmática, está sujeto a modificaciones químicas por diversas enzimas. En los últimos años se han descrito fosforilaciones y otras modificaciones covalentes. Aún no se sabe con precisión para qué ocurren dichos cambios, pero es posible pensar que estas reacciones, de alguna forma, sean señales para el procesamiento de la información que lleva el mensajero.
Los receptores citoplasmáticos tampoco permanecen estáticos; al igual que los de membrana, cambian su localización en la célula. Actualmente se sabe que viajan al núcleo, su movilización es parte fundamental del mecanismo de transmisión del mensajero. Esto se verá con más detalle más adelante. Sin embargo, es importante hacer notar aquí que su función básica es la misma: reconocer al mensajero y continuar el proceso de activación celular.
Como mencionamos arriba, podemos dividir a los receptores por su localización en la célula en dos grandes familias: los que se localizan en la membrana plasmática y los intracelulares.
Los receptores que se encuentran en la membrana plasmática tienen diferentes características tanto estructurales como funcionales, lo que ha permitido dividirlos en varios grupos. Mencionaremos ahora los grupos principales para ir explicando cada uno de ellos, poco a poco, en los siguientes capítulos. Estos son: 1) Los receptores acoplados a proteínas G, 2) los receptores con actividad enzimática, 3) Los receptores que carecen do actividad enzimática pero que se acoplan a enzimas itinerantes, y 4) los receptores canal.