V. SI TÚ ME RASCAS LA ESPALDA, YO TE LA RASCO A TI. LAS INTERACCIONES MUTUALISTAS
| He llegado a las ramas de los árboles enflorados: cual colibrí enjoyado gozo aspirando (su olor, con el cual deleitó suavemente, gratamente, mis labios. Poesía náhuatl |
EL ÚLTIMO tipo de interacción biespecífica que trataremos es aquel en el cual ambas especies participantes se benefician, por lo que se puede simbolizar por ++. Algunas de estas interacciones (llamadas técnicamente mutualismos) son bien conocidas por todo el mundo (por ejemplo, las de los polinizadores y sus plantas), y siempre han interesado a los naturalistas. Sin embargo, sólo hasta tiempos relativamente recientes los ecólogos de poblaciones han empezado a considerar las interacciones ++ como importantes e interesantes. Hasta la década de los setenta los mutualismos se catalogaban como parte de una historia natural fascinante, pero sin mayor importancia poblacional.
Obviamente, hay una larga lista de precursores que desde el siglo pasado se preocuparon por las interacciones ++, pero siempre al margen de la corriente principal de la ecología teórica. Es interesante preguntarse por qué en los países anglosajones los fenómenos de la competencia y la depredación fueron considerados interacciones sobre las cuales valía la pena teorizar, mientras que el mutualismo fue tradicionalmente del interés de biólogos con antecedentes anarquistas (Kropotkin), revolucionarios (Kostitzin) o cuáqueros (Allee). Fuera cual fuese la razón, el hecho es que actualmente ya ningún ecólogo duda de la importancia de las interacciones ++. Incluso se ha propuesto que los eucariontes (los organismos cuyas células poseen un núcleo verdadero; o sea la totalidad de los organismos, con excepción de las bacterias y algunos otros grupos menores) se originaron a partir de una interacción mutualista.
A semejanza de las interacciones -+, hay una gran variedad de mecanismos biológicos que producen el balance demográfico ++. Para tratar de ordenar esta diversidad, comenzaremos mencionando las principales "monedas" con las que se "paga" el mutualismo. Existen tres tipos principales de beneficios:
En primer lugar, los beneficios tróficos o alimentarios. Una especie puede ofrecer alimentos en forma directa a sus mutualistas, como el néctar y/o el polen que proporcionan las flores a sus polinizadores; los parásitos y desechos que los organismos limpiadores reciben de sus clientes, etc. El beneficio alimentario también se puede dar en forma indirecta, como la flora y fauna intestinales de los animales, que ayudan a digerir compuestos que el animal en cuestión sería incapaz de asimilar por sí solo (por ejemplo, la celulosa de las plantas); o como las bacterias nitrificantes de las leguminosas, que convierten en amonio el nitrógeno atmosférico; o las micorrizas de los bosques, que aceleran grandemente los procesos de degradación y absorción de la materia orgánica por las raíces de los árboles.
En segundo lugar, hay beneficios derivados de la transportación de gametos, propágulos o individuos adultos. Las plantas, al ser en general incapaces de moverse de un sitio a otro, requieren de ayuda para dispersar su polen y sus semillas. Existe además una gran variedad de especies de animales que necesitan obligadamente de dispersores. Por ejemplo, como señala la doctora Ana Hoffmann en otro volumen de esta misma serie, muchos ácaros e insectos pequeños son foréticos, esto es, se transportan en las patas de escarabajos o mariposas, en los picos de las aves, etcétera.
Una tercera gran categoría de beneficios es la protección. Ésta puede ser muy directa, como en el caso de las anémonas, cuyos tentáculos venenosos dan refugio a los peces payaso en los arrecifes tropicales; o las hormigas, que protegen a las acacias y los guarumos (Cecropia) tropicales del ataque de herbívoros y parásitos; u otras especies de hormigas, que protegen muy eficientemente a las larvas de ciertas mariposas, o a los pulgones, de la presencia de los parasitoides. También puede existir protección en un sentido más amplio, como el ambiente relativamente estable y seguro que los endosimbiontes (la flora y la fauna intestinales de muchos animales; las micorrizas de las plantas, etc.) encuentran dentro de sus hospederos. Asimismo, se considera protección el servicio de limpieza que muchos organismos realizan sobre otros, para remover parásitos o partículas inutilizadas de comida, como hacen ciertas aves con los búfalos, los rinocerontes o los cocodrilos, y los peces limpiadores que incluso ocupan territorios fijos a los que acuden los "clientes" en demanda de "servicios".
Una especie mutualista puede estar necesariamente ligada a su "socio", en cuyo caso la desaparición de éste conduce a la desaparición de la población del mutualista obligado. Los ejemplos del primer caso abundan entre los mutualismos simbióticos, es decir, aquellos en los que la asociación es íntima, como las bacterias digestivas, los hongos y las algas en los líquenes, etc. También se encuentran algunos mutualismos obligados no simbióticos, por ejemplo, ciertas orquídeas del género Catasetum que son obligatoriamente polinizadas por abejas Euglossidas. Es posible que también éste sea el caso del árbol Calvaria major de la isla Mauricio: se ha argumentado que las poblaciones de esta especie no han producido ni una sola plántula en los últimos 300 años, debido a que estaban asociadas obligadarnente a los dodos. Éstos eran unas aves grandes, torpes y mansas que hasta finales del siglo XVII habitaban la isla Mauricio. Los marineros europeos que llegaban ahí las mataban, en parte por su carne y en parte por gusto. En efecto, eran muy dóciles y mansas y no podían volar, por lo que había quien consideraba que matarlas a palos, nada más porque sí, resultaba divertido. Como consecuencia de este "deporte", antes de que finalizara el siglo XVIII los dodos ya se habían extinguido. Ahora bien, se sabe que las semillas de muchas especies de plantas necesitan pasar por el tracto digestivo de las aves para poder germinar, y entonces se propuso la hipótesis de que las semillas de Calvaria major requerían de ser tragadas y luego excretadas por los dodos para su germinación. Temple, en 1977, reportó el resultado de un experimento que consistió en dar semillas de Calvaria a guajolotes (lo más cercano a los dodos que Temple encontró): un 17% de las semillas que estos animales excretaron germinó, lo cual apoya la hipótesis de que, en la naturaleza, Calvaria dependía obligatoriamente del dodo para su reproducción.
Posiblemente la mayoría de los mutualismos sean facultativos (no obligados). Hay grupos (llamados gremios) de especies que realizan tareas similares dentro de una comunidad particular. Por ejemplo, hay varios gremios de polinizadores, constituidos por especies con características similares, que se reparten las visitas a los distintos gremios de plantas. Así tenemos a los colibríes y abejorros de proboscis larga que visitan a las plantas del gremio de flores rojas con corola estrecha y alargada. La remoción de una sola especie del gremio posiblemente no afecte de modo severo a cualquier otra del gremio asociado, puesto que existen especies alternativas que pueden realizar las funciones de la desaparecida.
Para explorar un poco las principales diferencias en la dinámica poblacional de los mutualismos facultativos y los obligados, plantearemos ahora algunos modelos de espacio de fase semejantes a los de los capítulos anteriores.
En general, debemos esperar que las isoclinas de especies mutualistas tengan
pendiente positiva, puesto que a mayor número de individuos de una de las dos
especies, se requerirá de un mayor número de mutualistas para compensar los
efectos negativos de la competencia intraespecífica. Por otra parte, el beneficio
demográfico derivado de la presencia del mutualista no puede aumentar indefinidamente.
Por ejemplo, por muchos polinizadores que existan, no es posible fecundar más
óvulos que los que cada planta tenga, y por mucho néctar y polen que las plantas
provean, las abejas no podrán aumentar su población más allá de lo que otros
factores limitantes determinen. Por lo tanto, las isoclinas deben de curvarse
para reflejar este efecto de saturación, de tal forma que cada pequeño aumento
en la densidad de una especie requerirá cada vez más mutualistas para compensar
los efectos negativos. Algunas isoclinas típicas de esta situación se muestran
en la figura V.l.
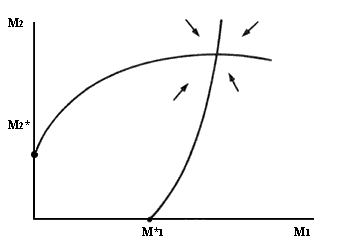
Figura V.1. Espacios de fase de una interacción entre dos especies de mutualistas
facultativos.
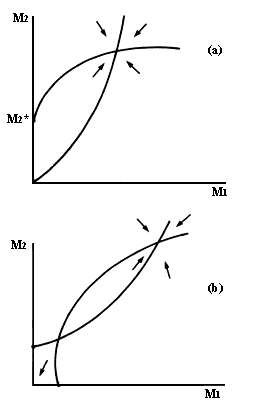
Figura V.2. Espacio de la fase de una interacción entre un mutualista facultativo
y otro obligado (a), y de dos mutualistas obligados, ambos con un umbral de
densidad de la especie mutualista (b).
La topografía del espacio de flechas indica que se alcanzará un equilibrio estable en el cual la densidad de cada especie es superior a la que se obtendría en ausencia del mutualista. Como en el caso de la competencia, es posible demostrar que no se presentarán ciclos en este tipo de espacio de fase. Hay que notar que se supone que cada especie puede sobrevivir en ausencia de su mutualista; esta suposición corresponde al caso de mutualistas facultativos. Tendremos entonces una interacción sumamente sólida, en la que las únicas perturbaciones capaces de simplificar la asociación son aquellas que excluyan completamente a una de las dos especies.
¿Qué pasaría si una de las dos especies, digamos la A, fuera facultativa? Esto se representa corriendo la isoclina de la especie A hacia la izquierda, para que su equilibrio se haga cero o negativo. Un equilibrio negativo se interpreta biológicamente diciendo que la población A necesita una densidad mínima B* de su socio para poder establecerse. En la figura V.2a tenemos ejemplos de ambos casos. Nótese que se sigue obteniendo un solo punto de equilibrio sobre el plano y que éste es estable. Si en cambio ahora hacemos que ambas poblaciones dependan obligadamente de los socios, entonces las isoclinas pueden cruzarse en dos puntos de equilibrio. Uno estable y otro inestable (Figura V.2b). Ahora el sistema entero puede desaparecer si alguna causa externa a la interacción mueve las poblaciones a la zona de atracción del origen. Por lo tanto, esperaríamos que los mutualismos obligados fueran menos comunes que los facultativos, pues los primeros resultarían más sensibles a las fluctuaciones medioambientales.
Hasta donde sabemos ahora, éste es el caso. Es casi seguro que ejemplos de asociaciones consideradas tradicionalmente obligadas, como las yucas y sus polinizadores, las de algunas micorrizas de ciertas especies de árboles, o la microfauna del tracto digestivo de las termitas (la cual digiere la celulosa que las termitas comen) en realidad lo sean, pero no se han realizado los experimentos de remoción de uno de los dos mutualistas que permitirían determinar el grado de obligatoriedad de la relación. En cambio, abundan los ejemplos de mutualismos difusos, en donde los "socios" mutualistas son gremios de especies que actúan similarmente. Por ejemplo, los gremios de dispersores de semillas (varias especies de aves) y las especies de árboles cuyas semillas dispersan. En este mutualismo, los dispersores obtienen las sustancias nutritivas de los frutos, y las plantas reciben transporte a sitios alejados de donde está la planta materna, lo cual puede ser benéfico si en la vecindad de la planta madre la competencia (por luz, por ejemplo) resultara más intensa. En esta área todavía hace falta realizar mucho trabajo experimental, de manipulación de densidades de especies, para entender bien el funcionamiento de las interacciones ++.
Sin embargo, con la información disponible ya se puede afirmar que los mutualismos son tan importantes como las otras interacciones en sus efectos a nivel de la comunidad. Pensemos solamente en la importancia de las micorrizas como mutualismos que permiten la existencia de los bosques y selvas. Es muy probable que si se removieran las micorrizas de los bosques, éstos sufrirían una profunda transformación e incluso muchas especies de árboles desaparecerían. De manera similar, sin dispersores de semillas, las selvas tropicales adquirirían una fisonomía muy diferente, pues su diversidad se debe en mucho a que las semillas son transportadas por una variedad de organismos que las mezclan y remezclan en extensiones muy amplias. Sin los dispersores, lo más probable es que muy pocas especies de árboles dominarían la composición de la selva, en lugar de la escasa dominancia específica que se observa. También en este caso queda por realizar casi toda la labor experimental que permitiría rechazar las hipótesis arriba mencionadas. El estudio ecológico de las interacciones ++ está aún "en pañales".
Sin embargo, aunque en muchos casos falta evidencia experimental, las adaptaciones que observamos en los participantes en las asociaciones ++ sugieren claramente que, por lo menos en el pasado, se produjeron los cambios demográficos necesarios para que actuara la selección natural. Por ejemplo, la morfología y la pauta de coloración en muchas flores promueve la visita eficiente de los polinizadores. Las rayas, manchas alargadas o hileras de puntos que muchas especies de flores poseen pueden servir para guiar a los polinizadores hacia las recompensas (néctar o polen) y aumentar así la eficiencia del polinizador. Esta eficiencia se mide por el número de visitas "exitosas" que el polinizador realiza por unidad de tiempo. La importancia de estas guías se ha demostrado experimentalmente por medio de tres técnicas principales: la primera consiste en utilizar flores artificiales en el laboratorio, algunas con guías y otras sin ellas, y medir la cantidad de visitas que los polinizadores realizan en un tiempo dado; la segunda, en pintar a las flores naturales en el campo; la última radica en utilizar especies en las cuales se presentan naturalmente flores con y sin guías. En los tres tipos de experimentos, las flores con guías reciben más visitas en el mismo tiempo porque los visitantes van directamente a la parte de la flor que contiene el néctar, mientras que en las flores sin guías se pierde mucho tiempo vagando sin encontrar esta recompensa.
En muchas especies de plantas, el aspecto de las flores, su coloración, el tipo de néctar y la hora de apertura se conjugan en los llamados "síndromes de polinización". Estos son conjuntos de características florales que las adaptan a la polinización por grupos restringidos de visitantes. Por ejemplo, el "síndrome de murciélago", o quiropterofilia, se manifiesta en flores grandes, de colores pálidos, con gran cantidad de néctar y de apertura principalmente nocturna. Este síndrome favorece las visitas regulares por parte de animales nocturnos, como los murciélagos, con altos requerimientos energéticos y capaces de detectar a las flores en la oscuridad. Otro síndrome fácilmente reconocible es el del colibrí. Corresponde a flores rojas o anaranjadas, de corola larga y estrecha, y de néctares ricos en aminoácidos. Los visitantes típicos serían animales de pico (o trompa) larga, buena vista y muy dependientes del néctar para su alimentación.
Aunque los síndromes tienen muchas excepciones en ambos sentidos (polinizadores atípicos visitan las flores del síndrome, y los visitantes típicos visitan flores que no lo presentan), son una valiosa guía que, "a ojo de buen cubero", puede indicar qué tipo de polinizador se puede esperar.
En este punto conviene subrayar algo que ya se mencionó en el primer capítulo. La selección natural no actúa (hasta donde nos consta) para beneficio de la especie y mucho menos de otras especies. En las interacciones ++ una especie beneficia a otra únicamente porque recibe algo a cambio. Más aún, estrictamente hablando, lo que ocurre es que el material genético responsable de la conducta mutualista produce más copia de sí mismo que aquellos genes que no propicien el efecto mutualista. Por lo tanto, por triste que pudiera resultar para algunos enamorados de la imagen de una naturaleza amable y altruista, el mutualismo en los seres vivos es únicamente un "egoísmo genético" disfrazado, y a la menor oportunidad se pueden seleccionar conductas que aporten a la especie el beneficio del mutualismo sin tener que pagar su costo.
En las relaciones entre polinizadores y plantas, este tipo de "trampa" ha aparecido un gran número de veces. Existen plantas cuyas flores se reparten el costo de producir el néctar, el cual es muy caro en términos de energía. La papaya, por ejemplo, tiene flores femeninas sin néctar, pero que producen aromas parecidos a los de las flores masculinas con néctar. Las flores femeninas son polinizadas por insectos que, debido a experiencias anteriores con flores masculinas, asocian el aroma con la existencia de una recompensa en forma de néctar. La "deshonestidad" de las plantas puede alcanzar extremos casi escandalosos, como en el caso de muchas orquídeas. Por ejemplo, en el género Ophirys, existen especies cuyas flores presentan una notable similitud con las hembras de ciertas avispas. El macho de la avispa se deja engañar por la orquídea y trata de copular con ella, creyéndola una hembra de su especie. Cuando por fin abandona la flor (sin ninguna recompensa, sea en néctar o de otro tipo), ya lleva colocada en el tórax la masa de polen pegajoso que la orquídea le depositó. Una ulterior visita a otra falsa avispa hembra culminará en la transferencia de la masa de polen a la zona receptiva de otra orquídea. La avispa realiza la transferencia del polen de manera notablemente precisa y sin ningún costo energético aparente para la planta.
Pero también existen los "ladrones de néctar", que le hacen trampa a las plantas perforando un pequeño agujerito en la base de la corola y extrayendo el néctar sin impregnarse de polen.
Otra interacción en la que se han producido adaptaciones verdaderamente maravillosas es la de los pequeños pececillos "limpiadores" de los arrecifes tropicales. Más de cuarenta especies de peces y por lo menos seis de camarones se dedican al "oficio" de limpiar diferentes partes del cuerpo de otros organismos, retirando partículas de comida, pequeños parásitos, etc. Cuando un individuo de alguna de las especies de "clientes" requiere limpieza, se dirige al sitio ocupado por un organismo limpiador. El acercamiento puede acompañarse por cambios en la coloración y la conducta del "cliente". Esta coloración sirve de señal para advertir al limpiador de que el cliente se aproxima como tal, y no como depredador. El limpiador corresponde con una pauta de nado muy precisa, que es seguida por la conducta de limpiado. Durante la limpieza, el encargado de ésta puede penetrar a la boca abierta del cliente y remover las partículas de alimento incrustadas en los dientes. Se ha demostrado experimentalmente, removiendo de una zona a todos los limpiadores, que sin ellos los peces clientes se enferman de la piel y sus poblaciones disminuyen. Tenemos, por lo tanto, un ejemplo demostrado de asociación ++.
Muchas veces las asociaciones mutualistas se ven invadidas por organismos llamados "aprovechados", es decir, que toman ventaja (se aprovechan) de la existencia del mutualismo pero sin dar nada a cambio, o hasta en perjuicio de uno o ambos participantes en el mutualismo. Un ejemplo de esto ocurre entre los limpiadores y sus clientes. El pez limpiador Labroides dimidiatus tiene un mímico, Aspidontus teniatus, el cual es extremadamente parecido al primero. Cuando un cliente en busca de limpieza se aproxima a un individuo de Aspidontus, éste imita incluso la "danza" de bienvenida del limpiador. El cliente se pone entonces a disposición del falso limpiador, quien en lugar de remover las basuras o los parásitos se aprovecha de la confianza del visitante para arrancar un buen bocado de agalla o de "cachete".
Otro ejemplo de aprovechado es el de la campamocha africana Hymenopus coronatus, que se aprovecha del mutualismo entre los polinizadores y sus plantas. El parecido de este depredador de insectos a un grupo de flores es realmente asombroso, de manera que los polinizadores, engañados, en lugar del néctar o polen encuentran su fin.
Como se ha visto, las interacciones mutualistas deberían de facilitar la existencia de las especies involucradas, tal vez incluso manteniendo las especies dominantes de una comunidad (micorrizas de los árboles, por ejemplo), por lo que su papel en la estructura de las comunidades puede ser central. Las presiones selectivas sobre los participantes en las asociaciones ++ deben de actuar en el sentido de estrechar cada vez más la asociación, a diferencia de lo que ocurre con la depredación, donde uno de los participantes (la presa) está sujeto a presiones selectivas que promueven la separación o disociación con el depredador; o la competencia, en donde ambos participantes deben de evolucionar en el sentido de separarse ecológicamente de los competidores.