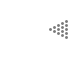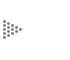III. QUÍTATE TÚ PARA PONERME YO. LA COMPETENCIA ENTRE ESPECIES
EL PUEBLO de Malinalco, en el Estado de México, es muy conocido por su espectacular templo del dios del viento. Malinalco también tiene un convento agustino del siglo XVI, en cuyo claustro algún anónimo artista indígena pintó unos interesantísimos murales. El tema de los murales no es bíblico ni religioso, sino profano y, a más abundamiento, naturalista. A primera vista, es difícil distinguir algún orden en las paredes blanquinegras y repletas de figuras y líneas tan entrecruzadas que pudieran sugerir un ejercicio de caligrafía arábiga. Sin embargo, al acercarse, el orden subyacente se manifiesta. Vemos entonces un bosque formado por una gran variedad de plantas, estilizadas pero reconocibles, habitado por garzas, colibríes, serpientes, coatíes, conejos, etc. Los animales se encuentran casi todos enfrascados en diferentes formas de lo que Darwin llamó "la lucha por la existencia": hay garzas devorando peces, serpientes a punto de atacar pajarillos, colibríes y palomas visitando flores, y un par de coatíes arrebatándose la comida.
¿Cuál habrá sido el propósito religioso o didáctico del mural? No sé si exista una respuesta a esta pregunta. Sería muy interesante averiguar los motivos que tuvieron los frailes agustinos del siglo XVI para encargar estos frescos. Y el artista (o artistas) indígena que los ejecutó, ¿por qué razón habrá escogido los ejemplos particulares?
Cualesquiera que hayan sido las razones religiosas o artísticas que expliquen el tema de los murales de Malinalco, desde mi punto de vista resultan particularmente interesantes porque en ellos están representadas las tres principales clases de interacciones entre dos especies: la competencia, la depredación y el mutualismo (donde ambas especies se benefician). Habiendo analizado ya los tipos de interacciones que ocurren más comúnmente entre los miembros de una misma especie, iniciaremos ahora la descripción de lo que ocurre entre diferentes especies.
Los casos más simples, mejor entendidos teóricamente y para los que se cuenta con una mayor cantidad de datos, son aquellos en los que intervienen preponderantemente sólo dos especies. Ahora bien, en la naturaleza todas las interacciones son multiespecíficas, hablando estrictamente. Sin embargo, como ya dijimos, está justificado metodológicamente tratar de entender primero los fenómenos más simples y menos reales o comunes para pasar después a los más complejos y usuales. Dicho metafóricamente, trataríamos de interpretar todo lo que se pueda del mural en términos de sus detalles. Lo global en términos de lo local. Personalmente creo que este ejercicio reduccionista no puede llevarse a un extremo: las propiedades de una comunidad biológica en toda su complejidad no pueden deducirse de la mera suma de las interacciones entre pares de componentes. Sin embargo, es indispensable entender estas interacciones para el planteamiento general del problema de la ecología de las interacciones multiespecíficas. Por otro lado, los casos para los que contamos con más datos cuantitativos y una mejor comprensión de la historia natural descriptiva, son los de interacciones biespecíficas.
En forma similar a la gráfica que en el capítulo anterior representaba los tipos de interacciones entre individuos de una misma especie, se puede integrar otra que represente los resultados de las interacciones entre dos especies, tomando como criterio los resultados demográficos de la interacción. Simbolizando con un + un aumento, con un - un decremento, y con 0 la ausencia de cambio en las tasas de crecimiento, tenemos: 1) la interacción resulta perjudicial para ambas especies (--); 2) beneficiosa para ambas (++); 3) beneficiosa para una y perjudicial para la otra (+-); 4) beneficiosa para una e indiferente para la otra (+0) y, por último, 5) perjudicial para una e indiferente para la otra (-0). Los tres primeros tipos de interacciones son aparentemente los más importantes y han sido bastante bien estudiados. Se denominan, por orden: competencia, mutualismo y relación depredador-presa (lato sensu). Los dos últimos tipos (+0 y -0) incluyen algunas interacciones fascinantes desde el punto de vista de la historia natural (por ejemplo, las foresis, o relaciones de transporte, las cuales se ejemplifican en el libro de la doctora Ana Hofmann, de esta misma colección). Sin embargo, su importancia en el desarrollo de la teoría ecológica ha sido bastante menor, por lo que no se mencionarán en lo sucesivo.
La primera clase de interacciones que analizaremos en este libro es la - -, es decir, las llamadas interacciones competitivas.
La interacción entre una especie A y otra B se llama competitiva cuando un aumento en los números de A causa una caída en los números de B y, de manera recíproca, un incremento en B redunda en una caída en los de A. Estrictamente, esta disminución en las tasas de crecimiento debe ser a causa de que la oferta de algún recurso común (espacio, comida, luz, etc.) sea limitada, por lo que, a altas densidades de población, una cierta proporción de individuos de ambas especies queda siempre sin alcanzar una cantidad óptima del recurso.
Es posible que la competencia por el recurso sea directa o incluso violenta; por ejemplo, la que se da entre ciertas abejas tropicales y los colibríes que explotan las mismas flores. Las abejas persiguen activamente a los colibríes y les impiden aprovechar el néctar. A este tipo de interacción se le denomina "competencia por interferencia". Por otra parte, la competencia puede darse sin que se establezca nunca un contacto físico directo. Pensemos otra vez en la competencia por el néctar de las flores. La sola presencia de una especie explotadora, al disminuir la cantidad de néctar, puede afectar a otra especie de explotador que visite las flores algo más tarde que la primera. Se da la interacción sin que los competidores se encuentren nunca juntos y se le llama "competencia por explotación".
La teoría de las interacciones competitivas fue de las primeras en desarrollarse en la ecología moderna y tuvo una gran importancia en la conformación actual de esta ciencia. En la década de los veinte, un matemático italiano, Vito Volterra, y un biofísico norteamericano, Alfred Lotka, propusieron independientemente las ecuaciones conocidas en la actualidad como modelos de Lotka-Volterra. No obstante ser esquemáticamente simples, dichas ecuaciones catalizaron de manera notable la actividad de muchos ecólogos. Algunos se lanzaron, casi febrilmente, a demostrar en el laboratorio o en el campo que las ecuaciones eran "correctas , mientras que otros más, claramente irritados por la brusca intromisión de matemáticos en los dominios de la historia natural, intentaron con la misma dedicación demostrar que las ecuaciones eran "incorrectas". Un tercer grupo se concentró en desarrollar teóricamente las implicaciones de las ecuaciones o en buscarles alternativas. Poco a poco, fue asentándose la idea de que la importancia de tales modelos simplificados no radicaba en su "veracidad" o "falsedad"; que su papel no era predecir o describir con exactitud los fenómenos ecológicos, sino más bien dar un contexto conceptual y sugerir las hipótesis de trabajo que guiaran la investigación empírica. Durante mucho tiempo, la investigación en ecología de poblaciones y comunidades (sobre todo en los países anglosajones) estuvo permeada por la noción de que la competencia era la interacción de mayor importancia en la estructura de las comunidades. Estudios posteriores han hecho hincapié menor en la competencia, aunque ésta sigue siendo la interacción preferida de una influyente escuela de ecólogos de comunidades.
A continuación presentaremos los fundamentos de la teoría de la competencia y posteriormente la ilustraremos con datos de laboratorio y de campo.
En el capítulo anterior se facilitaba el planteamiento de las ecuaciones de
crecimiento poblacional suponiendo un crecimiento en generaciones discretas
o separadas. Sin embargo, a partir del presente capítulo resulta más simple
suponer que las especies crecen continuamente: esto es, que los nacimientos
y muertes ocurren a lo largo de todo el año, sincrónicamente. Si aceptamos esto,
así como algunas otras premisas que se analizarán después, se pueden plantear
las siguientes relaciones:
| Tasa de crecimiento del competidor A |
|
Tasa de crecimiento del competidor A en ausencia del competidor B |
|
Tasa de mortalidad producida por el competidor B |
| Tasa de crecimiento del competidor B |
|
Tasa de crecimiento del competidor B en ausencia del competidor A |
|
Tasa de mortalidad producida por el competidor A |
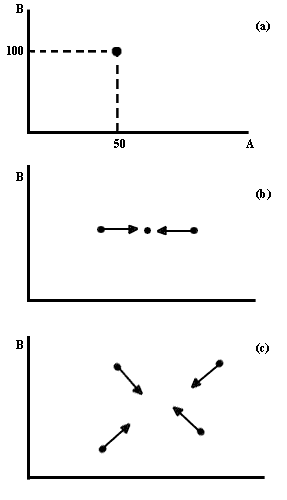
Figura III.1. Representación de los cambios poblacionales mediante el espacio
de fase.
Al llegar a este punto deberemos abrir un paréntesis para describir la herramienta de análisis que aplicaremos al estudio de estas relaciones, y las correspondientes a los capítulos posteriores. En esencia, se trata de la representación geométrica del sistema mediante lo que se conoce como espacio de fase. La idea es simple y elegante: en un plano representamos a la densidad poblacional de la especie A como distancias medidas en el sentido horizontal, y a la especie B como distancias medidas en un sentido vertical (Figura III.1a). Cualquier combinación concebible de especies corresponde a algún punto en el plano. Así, el origen corresponde a la ausencia de A y B; el eje de las X a cualquier densidad de A en ausencia de B, etcétera.
Supongamos que para una combinación dada de A y B, por ejemplo 50 y 100 (véase la figura III.la), calculamos el tamaño de la tasa de crecimiento de la especie A (usando una ecuación como la planteada arriba). Esta tasa de crecimiento puede representarse como una flecha sohre la línea horizontal que pasa por el punto 50, 100. En caso de que la tasa de crecimiento sea positiva (la población tienda a crecer), convendremos en apuntar la flecha hacia la derecha; si la tasa es negativa, la flecha apuntará hacia la izquierda; y si la tasa es de cero (esto representa una población en equilibrio) entonces no hay flecha (véase la figura III.lb). Lo propio se puede hacer para la especie B, con la diferencia de que las flechas se encuentran ahora sobre líneas verticales. Tenemos entonces representadas en un plano geométrico no sólo todas las posibles combinaciones de las dos especies, sino también la tendencia demográfica de cada una. Podemos resumir la información usando una sola flecha para cada punto, en lugar de dos, si aceptamos que la longitud de cada flecha sea proporcional al tamaño de la tasa de cambio en ese punto. La "suma vectorial" de ambas flechas, llamada la resultante, puede tener cualquier dirección (ya no sólo vertical y horizontal) y representa la dirección hacia la que tendería a moverse el punto que representa a una combinación dada (Figura III. lc). Nuestra representación geométrica adquiere entonces una dimensión dinámica que antes no tenía: si conocemos la "topografía" de las flechitas, podremos visualizar hacia dónde se moverá cualquier punto que pudiera ser de interés.
Por razones matemáticas que tienen que ver, entre otras suposiciones, con la de que el cambio en las poblaciones es continuo y no se da por generaciones separadas, "la topografía" del espacio de las flechitas resulta simple y regular. El plano se encuentra dividido en regiones dentro de las cuales la dirección general de las flechas es la misma. Esto es, dentro de una misma región no es posible encontrar flechas que apunten, por ejemplo, en direcciones opuestas. Hay que cambiar de región para encontrar cambios cualitativos en la dirección de las flechitas. Los tipos de regiones son pocos: 1) regiones donde ambas poblaciones crecen; 2) donde ambas decrecen; 3) donde una crece y la otra decrece, y 4) donde una o ambas se encuentran estacionarias. Las primeras tres son, en general, áreas, mientras que en la cuarta categoría encontramos las líneas o puntos que sirven de fronteras a las regiones de los primeros tres tipos. La colocación, tamaño y forma de cada posible región están determinados por los detalles biológicos de la interacción que se está modelando. En particular, a los tres tipos básicos de interacción entre dos especies (++, +- y - -) corresponden también tres tipos básicos, cualitativamente distintos, de disposición de las regiones mencionadas. El lector que haya seguido la argumentación hasta el final estará en condiciones de comprender la teoría básica de las interacciones entre dos especies.
Una vez cerrado el paréntesis regresemos a la competencia interespecífica.
En general, las ecuaciones que nos sirven para representar la competencia entre
dos especies diferentes corresponden a un espacio de fase del estilo del que
aparece en la figura III.2a. Las líneas (llamadas "isoclinas cero") corresponden
a las combinaciones de competidores que permitirían que cada especie se mantuviera
con una densidad estacionaria. Dicho en otras palabras, a lo largo de la línea
correspondiente a la especie A, ésta se encuentra en equilibrio, aunque B no
necesariamente se halle igual y viceversa. Si existe un punto en el que las
dos isoclinas se cruzan, éste corresponde a la combinación de densidades donde
ambas especies estarían en equilibrio al mismo tiempo. El caso más simple es
aquel en el cual no hay intersecciones. En la figura III.2b se indica la topografía
de las regiones de flechitas.
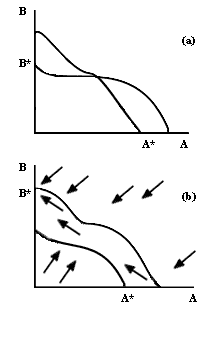
Figura III.2 Isoclinas cero para un sistema competitivo (a), y espacio de
fase de un sistema competitivo con exclusión de la especie A (b).
Resulta claro que desde cualquier punto (esto es, combinación inicial de especies) en que se empiece, la comunidad se desplazará poco a poco hacia un punto sobre el eje de la especie B que corresponde a la desaparición de la especie A, a lo cual se le llama exclusión competitiva. Si la isoclina de A es la que se encuentra por arriba, la excluida resultaría la especie B. La topografía detallada del espacio de flechas se obtiene a partir de la forma específica de las ecuaciones correspondientes a cada sistema. Aunque el procedimiento no es complicado, no se presentará aquí. El texto de Begon, Harper y Townsend es una excelente obra de consulta para obtener más detalles al respecto.
Ahora bien, en el caso de que las isoclinas se intersecten en un punto, pueden
darse dos posibilidades, ilustradas en la figura III.3: a) Se excluye
una u otra de las especies, dependiendo de la combinacion inicial, o b) ambas
especies de competidores coexisten indefinidamente. Tenemos entonces tres casos
donde hay exclusión y uno donde se puede dar la coexistencia. Biológicamente
hablando, para que ocurra la coexistencia es necesario que la tasa de crecimiento
de la especie A sea positiva cuando A es muy poco abundante y B está en su tamaño
máximo en ausencia de competidores, y viceversa. En términos gráficos, se requiere
que en la cercanía de los puntos A* y B* de la figura III.3b, tanto la flechita
de B como la de A, respectivamente, se dirijan hacia adentro del plano, y ninguna
hacia afuera.
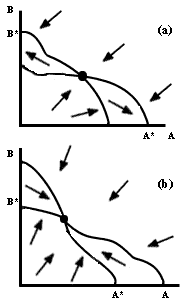
Figura III.3. Espacios de fase que corresponden a exclusión dependiente
de la población inicial (a), y coexistencia. (b).
La teoría de la competencia, delineada aquí, sugiere que en un sistema con dos competidores hay una tendencia a la exclusión de uno de ellos, y con ella se indican las condiciones precisas para que se dé la coexistencia. Ahora bien, no es posible, tanto por razones prácticas como fundamentales, obtener las ecuaciones "exactas" de la competencia entre dos especies antes de realizar el experimento. En efecto, no existen unas ecuaciones exactas de la competencia, pues como ha sido demostrado en estudios de laboratorio muy detallados, el modo mismo (los parámetros o incluso la forma de las ecuaciones) de competir va cambiando con cada generación, de tal suerte que no es posible predecir con exactitud el resultado de una interacción competitiva con base en las ecuaciones de Lotka-Volterra. Por lo tanto, la predicción resultante de la teoría de la competencia parecería reducirse, como Hutchinson ha hecho notar, a la frase vacía "dos especies que compiten, o coexisten o no lo hacen". La teoría de la competencia resultaría ser, más que nada, una clasificación de los posibles resultados de la interacción.
Sin embargo, la imagen anterior es injusta. Contar con una teoría permitió a los ecólogos encuadrar sus observaciones dentro de un marco conceptual. La búsqueda de "confirmaciones" o "refutaciones" de la teoría se convirtió pronto en mucho más que una mera lista de ejemplos y contraejemplos: se clasificaron grandes grupos de organismos por su tendencia a competir usualmente o no; se inició el estudio de los mecanismos biológicos que promueven la coexistencia; se desarrolló la hipótesis de que la competencia era la principal determinante del número de especies que podían vivir juntas en una comunidad; la necesidad de realizar las precisas observaciones requeridas por la teoría llevó a afinar y desarrollar la metodología experimental en la ecología de campo, etc. Teoría y observaciones se estimularon mutuamente, en un juego que incluía al mismo tiempo refutaciones y confirmaciones, estímulo a la imaginación y su desaliento, todo dentro de una ortodoxia en la que se consideraba un poco herético suponer que la competencia no fuera la interacción biológica por excelencia.
Esta fructífera influencia recíproca entre teoría y práctica se inició cuando en la década de los treinta, inspirado por las ecuaciones de Lotka-Volterra, el biólogo ruso G. F. Gause realizó experimentos de laboratorio con dos especies de protozoarios, en las que observó la exclusión en algunos casos y la coexistencia en otros. A los experimentos de Gause les sucedieron otros en los que se utilizaban gorgojos (insectos que se alimentan de granos) y en los cuales, dependiendo de las condiciones de temperatura, se obtenía la exclusión de una u otra especie. Estos experimentos (junto con las ecuaciones de Lotka-Volterra) se utilizaron para dar apoyo a lo que después se denominó "principio de exclusión competitiva", que durante un corto tiempo gozó del dudoso privilegio de ser considerado el dogma de la ecología de poblaciones: "dos especies suficientemente parecidas no pueden coexistir en la naturaleza". Por "suficientemente parecidas" se entiende que utilizan los mismos recursos en forma similar. Los naturalistas, sin embargo, conocían un buen número de contraejemplos al principio de exclusión competitiva. El debate subsecuente parecía separar por un lado a teóricos y experimentalistas y por otro a naturalistas, pero muy pronto un grupo de naturalistas especializados en ecología de aves y roedores comenzó a tomar partido a favor de la validez del principio de exclusión. Para la década de los cincuenta parecía haberse llegado a un impasse, con los ecólogos de poblaciones divididos en dos grandes escuelas: la que consideraba que los fenómenos de regulación dependientes de la densidad eran fundamentales en el funcionamiento de las poblaciones, y los que pensaban que las poblaciones no estaban reguladas y que el principio de exclusión competitiva era un concepto que carecía de contenido empírico.
A mediados de los cincuenta, George Evelyn Hutchinson y su alumno de doctorado, Robert MacArthur, presentaron un notable avance teórico, seguido por un extraordinario estudio de campo, lo que prestó nuevos ímpetus a los estudios sobre la competencia. Dicho avance consistió en la formalización de la teoría del nicho ecológico. Hutchinson presentó en términos matemáticos la vieja idea de que los organismos, al ocupar un determinado espacio físico, al estar activos a horas definidas del día, al utilizar ciertos recursos con preferencia a otros, etc., definen un cierto "nicho", propio de cada especie o población; la competencia se da dentro de las partes en común de los nichos de diferentes especies. Posteriormente, MacArthur reformuló los coeficientes de las ecuaciones de Lotka-Volterra en términos de esta idea.
El trabajo de campo, la tesis doctoral de MacArthur, consistió en aplicar a un sistema natural las ideas anteriores. El autor estudió uno compuesto por varias especies de warblers (cerrojillos o reinitas), pajarillos residentes en un bosque de Nueva Inglaterra. Las cinco especies de reinitas son sumamente parecidas y es necesaria cierta experiencia en la observación de aves para distinguirlas en el campo. Más aún, era bien sabido que las cinco especies se alimentaban de insectos, y como los bosques de abetos en Maine y Vermont son bastante homogéneos, el caso de las reinitas podría considerarse como un magnífico ejemplo de especies cercanas que no se excluían, esto es, constituían una contradicción al "dogma" de la exclusión competitiva.
MacArthur invirtió mucho tiempo en la observación y en el estudio de las actividades de las reinitas, así como de la zona de los árboles donde transcurrían éstas. Uno de los más importantes resultados de este estudio fue la demostración de que cada una de las cinco especies pasa la mayor parte de su tiempo en diferentes zonas del árbol (véase la figura III.4). Dicho en otras palabras: MacArthur logró demostrar que, aunque aparentemente las reinitas en Nueva Inglaterra ocupaban el mismo nicho (silvícolas, diurnas, insectívoras), en realidad un estudio más fino revelaba importantes diferencias en su forma de explotar los insectos del bosque, y por ende los nichos de las especies resultaban diferentes. La consecuencia es que, al no ser suficientemente parecidas desde un punto de vista ecológico las diferentes especies de reinitas no constituyen verdaderos competidores entre sí.
La idea de que para que dos especies sean competidoras deben tener nichos sobrepuestos,
junto con la demostración de que las diferencias en los nichos pueden ser muy
sutiles y difíciles de observar, estimularon una sucesión de estudios de campo
en donde se pretendía explicar la coexistencia de especies cercanas mediante
el descubrimiento de inevitables diferencias en sus nichos.
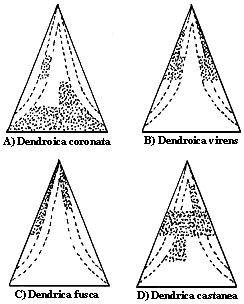
Figura III.4. Segregación del uso de hábitat en las "reinitas" estudiadas
por R. H. MacArthur.
Así, diferencias en los tamaños de los picos de aves granívoras, en las profundidades a las que los peces depredadores buscan a sus presas, en los periodos de floración de las plantas, etc., se propusieron como pruebas de la acción de la competencia, recurriendo al argumento: "estas especies son diferentes porque si no lo fueran ya alguna habría sido excluida". Sin embargo, este argumento no es susceptible de falsificación, es decir, no se puede diseñar, ni en principio, una observación que llegue a refutar el argumento anterior porque: 1) no es posible especificar ni medir el grado de diferencia que se requeriría para que tuviera que presentarse la exclusión, y 2) el ecólogo no tiene acceso a información cuantitativa sobre la historia pasada de las especies que actualmente coexisten, ni sobre la de aquellas que ya no lo hacen. Siendo infalsificable, dicho argumento es sugestivo y motivador, pero de dudosa utilidad directa.
Conforme más ecólogos se percataban de lo anterior, a las observaciones de gran detalle se empezaron a añadir experimentos de campo. Básicamente se intentaba, mediante manipulación artificial de las densidades de las especies en supuesta competencia, demostrar que los signos de las tasas de cambio eran ambos negativos. Por ejemplo, en un experimento realizado con lagartijas del género Anolis, se dividió el área en varias parcelas. En algunas se retiró a la especie A de Anolis, en otras a la especie B, y en otras se mantuvieron las dos, para tener un control experimental. Si se demostraba (como en efecto ocurrió) que la ausencia de A tenía un efecto positivo en la tasa de crecimiento de B, y viceversa, se presentaría un fuerte argumento en favor de la existencia de una interacción competitiva y entonces sería posible interpretar con mucho mayor rigor las observaciones de historia natural que documentaban diferencias de nicho.
Aunque los experimentos de campo en esta área no son ninguna novedad (Darwin realizó algunos clásicos), sí es cierto que empezaron a hacerse comunes a partir de la década de los sesenta, con un predominio de los realizados con plantas, no sólo porque muchas plantas son sujetos experimentales más sencillos que los animales, sino porque se contaba con la experiencia de los agrónomos, quienes por largo tiempo habían realizado experimentos semejantes a aquellos que los ecólogos comenzaban a intentar.
Los experimentos de campo del ecólogo moderno son, en su mayoría, muy difíciles de realizar. Se requieren diseños experimentales con un fundamento estadístico a veces muy complicado, y la puesta en práctica del experimento suele implicar muchos días de trabajo duro y rutinario, marcando, contando, rastreando y midiendo organismos que nunca cooperan con el experimentador. Además, como el control que el ecólogo tiene sobre una serie de variables importantes (por ejemplo, el clima), es nulo o muy limitado, los resultados de los experimentos rara vez admiten una explicación inequívoca. Sin embargo, junto con métodos cuantitativos en la observación de la historia natural de los organismos, los experimentos le han dado a la ecología de poblaciones de campo un alto grado de rigor.
El ya largo periodo (más de sesenta años) de desarrollo teórico-experimental de la noción de la competencia interespecífica, ha tenido consecuencias de gran importancia para la ciencia de la ecología. Hemos marcado la relevancia de algunas de las consecuencias de tipo metodológico: una más correcta apreciación del papel de los modelos teóricos, no tanto como herramientas productivas sino heurísticas, así como el desarrollo de un grado mucho mayor de refinamiento en el trabajo empírico, tanto experimental como observacional. Mencionaremos ahora algunos de los principales resultados generales que podemos aceptar como válidos, dada la situación actual de desarrollo de la ecología.
En primer lugar, parece claro que hay grandes grupos de organismos para los cuales la competencia interespecífica rara vez resulta ser una interacción importante. Un ejemplo de lo anterior son los insectos herbívoros, sobre todo en climas templados o con estaciones muy marcadas. Para ilustrar esto podemos referirnos al estudio de las comunidades de insectos asociadas al helecho Pteridium aquilinum. Esta asociación ha sido estudiada por John Lawton en Inglaterra y Hawai y Colorado EUA, y ha sido imposible demostrar la presencia de competencia interespecífica, ni directa ni indirectamente. Existen ejemplos similares en otras comunidades de insectos.
Por otra parte, en muchas poblaciones de vertebrados, como en aves, pequeños roedores, lagartijas, etc., la competencia interespecífica se ha demostrado en forma experimental. Similarmente, no hay duda de que entre muchas especies de plantas se presenta una a veces muy aguda competencia por recursos tales como la luz, el agua, o micrositios adecuados para germinar. Joseph Connell, en 1982, analizó 527 experimentos de campo reportados en la bibliografía por diferentes autores. Para casi la mitad de las especies estudiadas, y en un 40% de los experimentos realizados, se pudo demostrar la presencia de interacciones competitivas. De modo que es indudable que existe la competencia, aunque su importancia relativa aún se cuestiona.
¿Cuáles son las consecuencias de la competencia interespecífica para aquellas especies o tipos de especies en las que se presenta? En primer lugar, la competencia, al ser una presión selectiva, tiene consecuencias adaptativas directas para aquellas especies que han evolucionado bajo su influencia. Las diferencias ("segregación") en los nichos de especies competidoras son a menudo atribuidas a los efectos de la competencia. Por ejemplo, cuando dos especies de ciertos ratones habitan el mismo sitio, sus tamaños promedio no se superponen. Esto es, una especie es algo menor en tamaño que la otra. En áreas diferentes, donde las mismas especies no coexisten, los tamaños promedio son aproximadamente iguales. Se explica la observación anterior como un resultado de la competencia que ha promovido la especialización en semillas más pequeñas por una especie y más grandes por la otra, para evitar esta interacción competitiva en aquellas regiones en que ambas especies viven juntas.
De manera similar, la competencia afecta las áreas de distribución de los organismos competidores, de tal forma que especies cercanas o de ecología muy parecida deben de coexistir geográficamente mucho menos a menudo que especies muy distintas. David Lack ha demostrado que tal es el caso para un buen número de grupos de aves de todo el mundo. Sin embargo, deberían encontrarse excepciones a la regla anterior en medios ambientes complejos, en donde se presentan posibilidades de subdividir el hábitat en diferentes zonas, o los recursos en diferentes partes, segregando los nichos, como se mencionó anteriormente.
En segundo lugar, la competencia puede inducir importantes similitudes en grupos completos de organismos en diferentes partes del mundo. Por ejemplo, James Brown ha estudiado diversas comunidades de roedores en el desierto sonorense. En sitios similares climática y topográficamente, las comunidades convergen hasta un grado sorprendente. Tanto el número de especies, como sus tamaños y la forma de utilizar los recursos son muy parecidos de un sitio a otro, pese a que las especies son diferentes en los distintos sitios. Algo muy semejante ha sido reportado por Martin Cody en las comunidades de aves de los chaparrales californianos y los matorrales chilenos: pese a la enorme distancia y la composición totalmente diferente de especies, la estructura de las comunidades (el número de especies, sus tamaños, la sobreposición de nichos, etc.), en ambas localidades es muy parecida.
En resumen, el estudio de las interacciones competitivas en la ecología de poblaciones ha tenido una gran importancia teórica, y aunque en la actualidad resulta muy claro que la competencia no es un fenómeno tan extendido, ni sus consecuencias son tan importantes como se pensaba hace pocos años, tampoco hay duda de que en ciertos sistemas y grupos biológicos constituye una interacción de gran relevancia y capaz de proporcionar cierta estructura a las comunidades.