II. EL METABOLISMO, O LAS TRANSFORMACIONES DE LAS MOLÉCULAS EN LAS CÉLULAS
EL METABOLISMO no es otra cosa que la enorme serie de cambios
que sufren las moléculas para convertirse unas en otras y en otras y en otras,
de manera complicada, al parecer interminable y que, desde luego, estamos muy
lejos de conocer en su totalidad. Son tantas las sustancias que componen a un
organismo que una gran proporción de ellas se desconoce, como sucede aun en
el caso de los organismos unicelulares. Pero a pesar de que el proceso total
es tan complicado, es posible definir algunos de sus componentes, y aun quienes
no tenemos un conocimiento profundo de la bioquímica y la fisiología podemos
tener una idea aproximada de cómo es y para qué sirve.
EL METABOLISMO PROPORCIONA MATERIALES Y ENERGÍA
Todas las transformaciones de las moléculas tienen dos funciones principales: la primera, proporcionar a la células, tejidos, órganos, etc., materiales que requieran para sus distintas funciones, siendo la más importante la renovación constante de sus propias moléculas; la segunda, obtener diferentes formas de energía para mantener las funciones vitales. Así, plantas y algas reciben como "alimento" materiales muy sencillos, como sales minerales, CO2 y H2O, pero su energía la obtienen del Sol, y con ella satisfacen sus necesidades para funcionar y fabricar sus materiales. Pero las plantas también proporcionan al hombre materiales y la energía que contienen sus enlaces, con lo cual lo ayudan a sobrevivir.
El metabolismo celular puede considerarse como una serie de caminos de ida y vuelta, formados por una gran cantidad de moléculas que se transforman constantemente. Estos caminos reciben a las que llegan al organismo o a la célula del exterior, pero además tienen sus propias moléculas.
CÓMO TOMAMOS LOS MATERIALES DEL EXTERIOR
Ya mencionamos que las plantas y la mayoría de las algas toman materiales muy sencillos del exterior y que sólo requieren de sistemas para transportarlos al interior de las células.
Los animales, el hombre incluido, recibimos pocas moléculas sencillas y una
gran cantidad de macromoléculas, como almidones, proteínas o grasas. Éstas
son sometidas al proceso de la digestión para hidrolizarlas o partirlas en sus
componentes (ver capítulo I), antes de ser absorbidas en el intestino y de entrar
propiamente al organismo. Así, lo que ingresa a la sangre para ser tomado por
las células son las moléculas simples: los aminoácidos, los ácidos grasos, el
glicerol (glicerina) y la glucosa. A partir de estas moléculas podemos analizar
las transformaciones de los diferentes tipos de sustancias que reciben las células.
Cuando toman sustancias del exterior, la mayoría de los organismos unicelulares
cuentan con enzimas que degradan las moléculas más grandes y toman luego los
productos de ese proceso. Algunos organismos unicelulares, o células animales
especializadas, como las amibas o los leucocitos, pueden tomar del exterior
moléculas grandes, o inclusive partículas y otros materiales, pero los digieren
antes de utilizarlos.
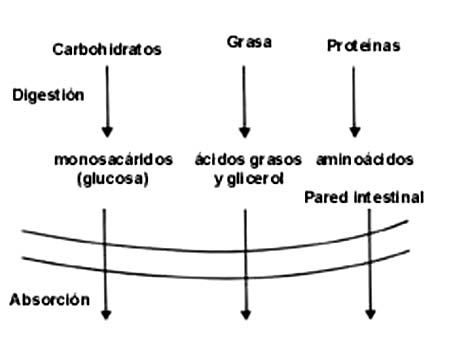
Figura II.1. Digestión y absorción.
ANABOLISMO Y CATABOLISMO (CONSTRUCCIÓN Y DESTRUCCIÓN DE MOLÉCULAS)
Una vez que las células y los organismos reciben esas moléculas sencillas, uno de los caminos que éstas pueden seguir es la síntesis de las llamadas macromoléculas (de macros, grande), como el almidón en las plantas o el glucógeno en los animales, que se forman de la unión de miles de moléculas de glucosa y en donde se almacenan azúcares. Por este proceso de síntesis se forman también las grasas y otros tipos de lípidos, como los fosfolípidos, que constituyen las membranas celulares, o las grasas neutras (mantecas o aceites), que en los animales se acumulan en el tejido adiposo, y en las plantas en algunas semillas.
En algunos animales, incluyendo al hombre, las grasas pueden ser un enorme almacén de reserva alimenticia. También por una síntesis se forman las moléculas de las diferentes proteínas, pero ni éstas ni los aminoácidos se pueden almacenar. Sin embargo, hay una perenne renovación en las células, en donde constantemente se necesitan aminoácidos para producir moléculas nuevas y degradar las existentes.
La figura II.2, además de mostrar un esquema de la posibilidad de que las moléculas
unitarias sean utilizadas para producir otras más grandes, señala también que
todas ellas se pueden degradar hasta producir CO2, H2O
y, en el caso de los aminoácidos, amoniaco (NH3). Distinguimos así
con claridad dos procesos. En uno de ellos, el anabolismo, a partir de moléculas
pequeñas se obtienen otras más grandes; en el otro, el catabolismo a
partir de moléculas grandes se obtienen otras más pequeñas.
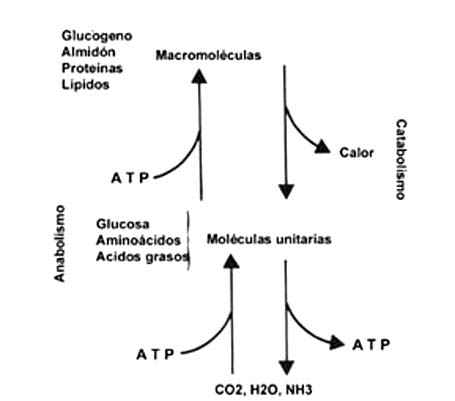
Figura II.2. Esquema del metabolismo y sus componentes, el anabolismo y
el catabolismo. Hay unidades para las grasas, los azúcares y las proteínas,
que pueden formar macromoléculas, o degradarse hasta CO2, H2O
y NH3, principalmente. Además, durante el catabolismo una parte de
la energía de los enlaces de las moléculas se transforma en calor, y otra en
diversas manifestaciones, como los enlaces del ATP (ver el capítulo III), o
los aceptores de hidrógenos.
Otra característica importante del proceso metabólico es que en el catabolismo de las moléculas pequeñas, como la glucosa, los ácidos grasos o los aminoácidos, se logra transformar la energía de sus enlaces químicos en la energía de los enlaces del ATP y otras sustancias, que proporcionan en forma directa la energía que requieren las células para todas sus funciones. Además, los procesos de síntesis, tanto de moléculas sencillas como de macromoléculas, requieren energía, la cual proviene del ATP y del llamado poder reductor que tienen las moléculas llamadas NADH y NADPH, entre otras. Es interesante señalar que la degradación que sufren las macromoléculas para producir las unidades que las componen hace que se transforme en calor toda la energía de sus enlaces.
Pero también en la degradación de las moléculas para producir otras más sencillas, o para sintetizar ATP, hay una liberación de calor.
LAS VÍAS METABÓLICAS Y LAS ENZIMAS
Otra característica del metabolismo es que cada una de sus transformaciones está movida casi invariablemente por una enzima diferente. Las enzimas son proteínas, las moléculas más complicadas de la célula, que se encargan de catalizar (es decir, de acelerar) las reacciones individuales del metabolismo. Aunque las reacciones químicas de muchos pasos metabólicos pueden ocurrir en forma espontánea, prácticamente todas ellas transcurrirían con una enorme lentitud si no existieran las enzimas. Éstas aceleran mucho (habitualmente mucho más de un millón de veces) las reacciones individuales del metabolismo. Casi cada reacción requiere de una enzima diferente para moverse con suficiente velocidad. Pero así como las enzimas son capaces de acelerar las reacciones químicas de los seres vivos, muchas pueden ser reguladas por muy diversas sustancias. Muchas ocasiones son las responsables de que, en un paso del metabolismo, toda una vía se mueva con mayor o menor velocidad.
La figura II.3 es un esquema del funcionamiento de una enzima, que es una proteína
grande, la cual promueve la transformación de una o más moléculas, que reciben
el nombre de sustrato, en otra u otras, llamadas productos. En
ocasiones también participan moléculas con funciones definidas, llamadas coenzimas.
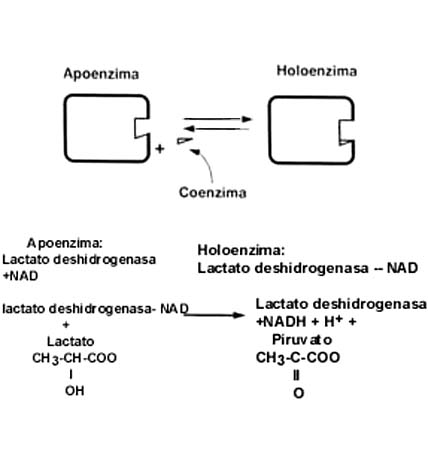
Figura II.3. Los componentes de un sistema enzimático: la enzima,
el o los sustratos
y el o los productos.
LAS COENZIMAS, LAS VITAMINAS Y EL METABOLISMO
En algunas reacciones del metabolismo, además de las enzimas toman parte, en pequeña cantidad, otras moléculas llamadas coenzimas, en cuya composición, en muchos casos intervienen las vitaminas. A continuación señalaremos algunas de ellas:
Nicotinamida. La componen dos coenzimas, el nicotín adenín dinucleótido (NAD) y el nicotín adenín dinucleótido fosfato (NADP); su estructura y funciones se describen en el capítulo III. Es la que porta los hidrógenos y sus electrones en muchas reacciones de oxidación, y se utiliza para la síntesis de algunas moléculas o en las transformaciones de energía en ATP. La deficiencia de esta vitamina produce la enfermedad llamada pelagra.
Riboflavina. Esta vitamina es componente de dos coenzimas relacionadas también con el transporte de los electrones en la cadena respiratoria: el flavín adenín mononucleótido y el flavín adenín dinucleótido. También participa en la cadena que transporta hidrógenos y electrones.
Ácido pantoténico. Es parte de la llamada coenzima A, que se muestra más adelante. Participa en el metabolismo de los ácidos grasos, pero muy especialmente en el de los fragmentos de dos átomos de carbono, que constituye la acetil coenzima A.
Tiamina. Esta vitamina participa en reacciones en las que algunos ácidos pierden su carboxilo (grupo-COOH). Las enzimas encargadas del proceso se llaman descarboxilasas.
Piridoxina. Participa en las reacciones de transferencia de grupos amínicos de los aminoácidos, como coenzima de diversas transaminasas, y en otras reacciones.
Éstos son sólo algunos ejemplos de la acción de las vitaminas en el metabolismo, como parte de las enzimas, en las llamadas coenzimas. Estas sustancias son en realidad una especie de segundo sustrato, o componente del sistema enzimático, que participa en la reacción. En las reacciones de deshidrogenación, por ejemplo, las coenzimas reciben electrones de un sustrato y los transfieren a otro, con la participación de otra enzima. En muchos de estos casos, las coenzimas son otro sustrato que también se modifica durante la reacción, pero tienen la característica de ser utilizadas en muchas reacciones diferentes. Al NAD, por ejemplo, lo encontraremos en la glucólisis, en el ciclo de Krebs y en reacciones de deshidrogenación de distintos tipos.
LAS PRINCIPALES TRANSFORMACIONES DE LOS AZÚCARES
Como ya se describió antes, los organismos tienen diferentes tipos de carbohidratos: monosacáridos, disacáridos, moléculas que se llaman oligosacáridos y polisacáridos, cuyo grado de agregación o polimerización depende de los diferentes monosacáridos. Sin duda, el eje del metabolismo celular es la glucosa; este monosacárido está distribuido en todos los organismos, ya sea en forma libre, como parte de otras moléculas, como los disacáridos sacarosa y lactosa, o en polisacáridos de reserva, como el almidón, el glucógeno o la celulosa, o en productos derivados, algunos de los cuales se mencionaron en el capítulo I.
Los almacenes de azúcar. Cuando la glucosa entra a una célula puede
tomar el camino hacia la síntesis de polímeros. La figura II.4 muestra cómo
en las células animales este azúcar, a través de varios pasos, se puede incorporar
a una molécula ya existente de glucógeno. En las células vegetales, uno de los
destinos de la molécula de glucosa es convertirse, por un mecanismo semejante,
en almidón. Tanto el glucógeno como el almidón y otros polímeros de los azúcares
forman sistemas de reserva en los organismos. Por lo tanto, también hay vías
para la degradación de estas sustancias. La misma figura II.4 muestra también
que la conversión de la glucosa en glucógeno es un camino reversible. De hecho,
cuando comemos se sintetiza el glucógeno para guardar los azúcares que hemos
ingerido. Luego, este polímero se va degradando lentamente entre una comida
y otra y mantiene el nivel de glucosa de nuestra sangre, pues muchos tejidos
requieren azúcar en forma constante. En los animales el glucógeno sirve para
almacenar azúcar, la cual se gasta entre una comida y otra; en las plantas el
almidón se almacena en las estructuras que aseguran su reproducción, como las
semillas de los cereales, o en los tubérculos, como las papas.
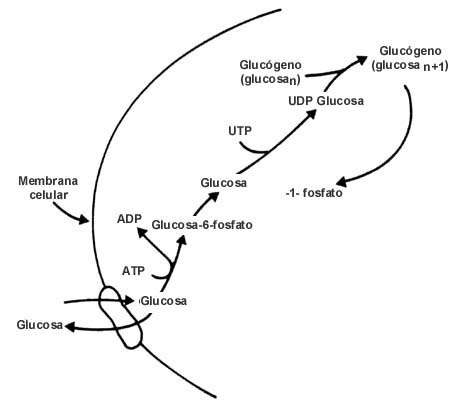
Figura II.4. El almacenamiento de la glucosa. En los animales se polimeriza
para fabricar glucógeno, que es la principal manera de almacenamiento; en los
vegetales crea almidón. Ambas formas pueden luego degradarse para dar de nuevo
glucosa, la cual aprovechan los seres vivos.
El otro camino que puede seguir la glucosa es su degradación. Para ilustrar
este caso pensemos en una célula animal, la muscular, por ejemplo. La figura
II.5 muestra las transformaciones que este azúcar sufre al convertirse en piruvato
(la sal del ácido pirúvico), o el lactato (la sal del ácido láctico). Las fórmulas
químicas están sólo como referencia.

Figura II.5. Esquema general de la glucólisis.
El esquema completo se presenta para mostrar la complejidad del sistema, aunque no está representado con todo detalle. A continuación destaco algunos puntos importantes que se dan cuando una célula degrada una molécula de glucosa:
A) Al degradar la glucosa para convertirla en lactato o piruvato sólo implica partir a la molécula, de seis átomos de carbono, en dos de tres. No obstante lo complicado que pueda parecer la vía metabólica, la transformación final es relativamente simple.
| C6H12O6 | ® | 2CH2 | — | CH | — | COOH |
| ½ | ||||||
| OH |
B) En este proceso, aunque al principio se invierten dos moléculas de ATP, después entran dos de fosfato (P) y cuatro más de ADP, para dar cuatro de ATP. El resultado neto es que al partir una molécula de glucosa en dos de lactato, se generan en forma neta dos moléculas de ATP a partir de dos de fosfato y dos de ADP.
C) Aunque la cantidad de ATP que resulta de cada molécula de glucosa es muy pequeña, esta vía puede ser extremadamente veloz. Durante el ejercicio muscular intenso, de un atleta por ejemplo, proporciona casi toda la energía que se requiere, compensando el rendimiento con la velocidad.
D) Otra cuestión interesante es que la degradación de la glucosa en esta vía se puede continuar por la transformación del piruvato en acetil coenzima A, hacia el llamado ciclo de los ácidos tricarboxílicos, o ciclo de Krebs; en él se degrada totalmente hasta dar CO2 y agua. En este ciclo, por otra parte, al conectarse con la fosforilación oxidativa, que se mencionará al hablar de las mitocondrias, se produce la mayor parte del ATP sintetizado. Por esta última razón la glucólisis es considerada también la vía de entrada de la glucosa y los azúcares en general a una de las vías catabólicas principales.
E) Finalmente, esta vía es en gran parte reversible; es decir, a partir del piruvato se obtiene glucosa, e incluso el mismo glucógeno y otros polisacáridos. Esto es importante, porque así es como se transforman en azúcares los aminoácidos, que provienen de las proteínas.
Como ya se vio en el capítulo I, las grasas neutras están compuestas de glicerol y ácidos grasos. Para degradarse, primero se separan los ácidos grasos de las moléculas. La parte más importante de la degradación de los lípidos consiste en la transformación, paso a paso, de las cadenas largas, de 16 a 18 átomos de carbono las más comunes, en fragmentos de dos átomos de carbono, como acetil CoA.
La coenzima A (figura II.6) es una molécula complicada que sirve como una especie de mango o asa que las enzimas requieren para el manejo de fragmentos de dos átomos de carbono o mayores, para formar la acetil (de acetilo) coenzima A, o el caso que sea. Sin embargo, la única parte que sufre cambios es la porción del ácido graso o el acetilo unido a la coenzima A. En la figura II.7 se presenta el proceso de degradación de un ácido graso con fórmulas químicas como referencia a su complejidad.
Aquí vemos cómo una molécula de un ácido graso, unida a la coenzima A, en varios pasos termina por perder, como acetil CoA, un fragmento de dos átomos de carbono. El proceso tiene las siguientes características:
A. Es un ciclo que se repite; partiendo de un ácido graso de, por ejemplo, 18 átomos de carbono, en la primera vuelta termina en uno de 16, pero en otra da uno de 14 y así sucesivamente, hasta que todo el ácido graso se convierte en fragmentos de acetil CoA, de dos átomos de carbono.
B. El proceso completo se lleva a cabo en la mitocondria.
C. En cada vuelta de este ciclo la célula recoge en dos moléculas especiales, el FAD y el NAD, que se convierten en FADH2 y NADH, dos pares de hidrógenos, que representan una forma de energía que se utiliza en la cadena respiratoria para sintetizar ATP.
D. Como el proceso degrada todo el ácido graso hasta fragmentos de acetil CoA,
con ésta se alimenta el ciclo de los ácidos tricarboxílicos, igual que con los
derivados del piruvato y de la glucosa, y se oxidan hasta CO2 y agua;
de aquí se obtiene energía en cantidades muy grandes.
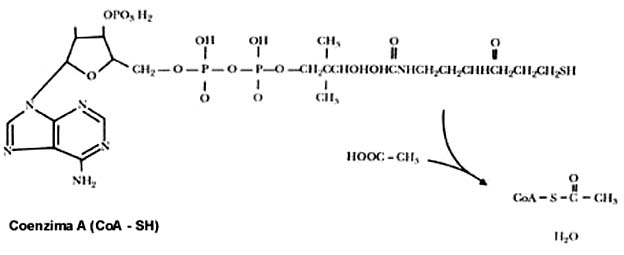
Figura II.6. La coenzima A y la acetil CoA.
E. Como en el caso de la vía anterior, la degradación de las grasas es en cierta
forma reversible. Aunque no intervienen las mismas enzimas ni cofactores, el
proceso de síntesis de un ácido graso es parecido, pero inverso a la degradación,
y en cada vuelta de un ciclo se añade a la molécula original un fragmento de
dos átomos de carbono. Esta nueva vía de síntesis es también útil para sintetizar
grasas a partir de azúcares o proteínas (figura II.5).
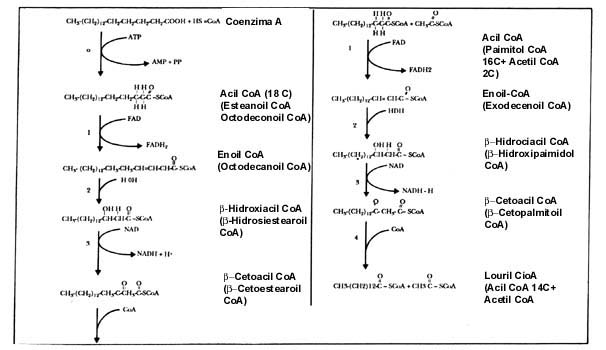
Figura II.7. La beta oxidación.
LAS TRANSFORMACIONES DE LAS PROTEÍNAS
Dado que en el capítulo IV ya se describieron los mecanismos de síntesis de las proteínas, en esta sección sólo mencionaremos los mecanismos de conversión de unos aminoácidos y su degradación. Las proteínas se degradan para dar aminoácidos, y viceversa, pero las transformaciones más interesantes son las de los aminoácidos.
En las células hay cetoácidos y aminoácidos, y mientras los primeros tienen
un grupo ceto (-C=O) en el carbono número 2, los segundos tienen un amino (-NH2).
Como se muestra en la figura II.8, un cetoácido (piruvato) se puede convertir
en un aminoácido (alanina), tomando el grupo amínico de un aminoácido (glutámico),
que en el proceso se convierte a su vez en cetoácido (cetoglutárico). Este proceso,
que recibe el nombre de transaminación, ocurre en muchos otros pares de cetoácidos
y aminoácidos y permite el intercambio de unos aminoácidos en otros.
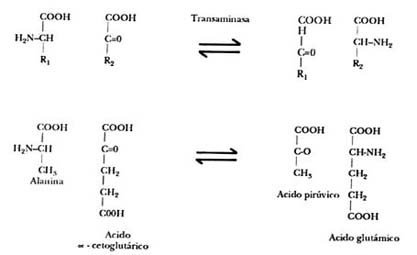
Figura II.8. La transaminación. Un cetoácido recibe el grupo amínico de un
aminoácido, y éste a su vez se convierte en cetoácido.
Por otra parte, los aminoácidos pueden perder su grupo amínico por otros procesos,
reacción en la cual liberan amoniaco (NH3) y dan lugar a un cetoácido.
Este proceso permite obtener cetoácidos diferentes, como el piruvato, el oxalocetato
o el cetoglutarato. El primero es el resultado final de la glucólisis y los
otros dos son pasos intermedios del ciclo de Krebs. De aquí resulta que, cuando
los aminoácidos pierden su grupo amínico, el residuo se puede incorporar a diferentes
caminos metabólicos para su degradación (figura II.9).
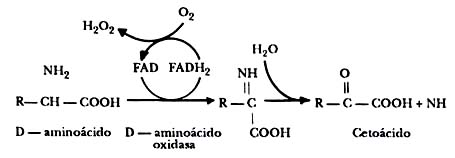
Figura II.9. La desaminación. Los aminoácidos pueden también perder su grupo
amínico y convertirse en cetoácidos.
Otro punto importante en el metabolismo de las proteínas es el destino del amonio que pierden al desaminarse. Dependiendo de los organismos, éste se puede eliminar como tal o como diferentes compuestos. En los humanos, una gran parte del amoniaco, que les resulta tóxico, se elimina después de unirlo con CO2, dando lugar a una molécula inerte, la urea:
| NH2 | ||||
| ½ | ||||
| C02 | + | 2NH3 | ® | C=O |
| ½ | ||||
| NH2 |
Finalmente, también hay enzimas capaces de sintetizar aminoácidos a partir
de cetoácidos y amoniaco, como la deshidrogenasa glutámica, que puede aminar
al cetoglutarato, o sea, ponerle un grupo amínico en lugar de cetónico, como
se muestra en la figura II.10.
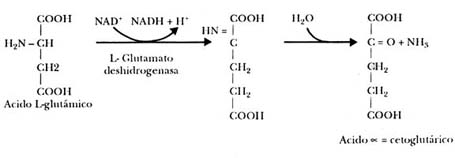
Figura II.10. Conversión del cetoglutarato en glutamato por la deshidrogenasa
glutámica.
EL CICLO DE LOS ÁCIDOS TRICARBOXÍLICOS
Ya hemos mencionado que de los principales componentes que obtenemos de los alimentos —azúcares y grasas principalmente, pero también de las proteínas— se pueden obtener moléculas como el piruvato, y a partir de éste la acetil CoA . Ésta, a su vez, puede provenir directamente de las grasas.
En la figura II.11 se muestra otro camino metabólico cíclico, el ciclo de los ácidos tricarboxílicos; pueden alimentarse con acetil CoA y tiene algunas caraterísticas importantes, que señalamos a continuación:
a) Se encuentra localizado en la mitocondria.
b) Termina en el mismo producto en que se puede considerar que se inicia, el ácido oxalacético u oxalacetato, que puede unirse otra vez con una molécula de acetato de la acetil CoA.
c) En el proceso, los dos átomos de carbono del acetato salen como CO2.
d) Entre los productos del ciclo se forman cuatro pares de hidrógenos, que pueden ser tomados por la cadena respiratoria para llevarlos al oxígeno y sintetizar ATP.
e) Muchos de los intermediarios del ciclo se forman de otras sustancias,
como por ejemplo, el cetoglutarato del glutamato, o el oxaloacetato del aspartato.
De esta forma se constituye en el mecanismo de conexión de diferentes vías metabólicas.
Tal vez la principal función sea la de servir de fuente muy eficiente de pares
de hidrógenos, para que, al entregar éstos a la cadena respiratoria se sintetice
ATP. De cada par de hidrógenos, la mitocondria puede obtener entre dos y tres
moléculas de ATP, de manera que es parte fundamental en los mecanismos de conservación
de la energía, que se verán más adelante al hablar de las mitocondrias y los
cloroplastos.
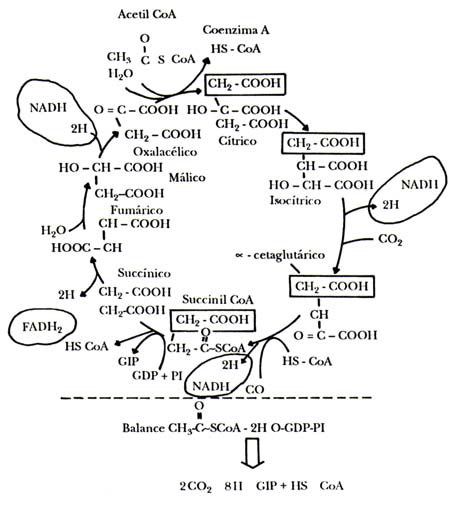
Figura II.11. El ciclo de los ácidos tricarboxílicos. Esta especie de molino
metabólico produce hidrógenos, una importante fuente de energía para la síntesis
del ATP, que van a la cadena de transporte de electrones de las células, y CO2,
a partir de muchas moléculas que provienen de diferentes partes del metabolismo.
UNAS VÍAS ESTÁN CONECTADAS CON OTRAS
De manera muy sencilla hemos descrito las transformaciones que sufren algunas moléculas, como la glucosa, los ácidos grasos y algunos aminoácidos. Tal pareciera que todas estas sustancias tienen caminos metabólicos definidos que las llevan a tal o cual producto, pasando por tal o cual sustancia intermedia. Pero no es así. Cada molécula o grupo de ellas puede participar en una vía y no necesariamente seguir el camino que hasta ahora hemos señalado. Incluso, en un mismo individuo o especie, al cambiar las condiciones de su vida o alimentación pueden producirse cambios también en el manejo de sus vías metabólicas. Así es como surgen las conexiones entre las vías metabólicas, y a manera de ejemplo señalaremos algunas de ellas.
Un caso sería la conexión entre el metabolismo de las grasas y el de los azúcares. El alimento de una persona se compone en buena parte de carbohidratos (pan, pastas, cereales, leguminosas, etc). En alguien que come lo justo, los azúcares, que le proporcionan la mayor parte de la energía para realizar sus funciones, entran en su mayor parte a las vías que ya señalamos, la glucólisis y el ciclo de los ácidos tricarboxílicos, y se degradan totalmente hasta CO2 y H2O, para producir ATP. También los alimentos se consumen o se gastan en función del gasto de ATP. De no ser el caso, disminuye la velocidad con que se utilizan, y se acumulan algunos de los productos, y entonces, aunque hay sistemas reguladores muy especiales, uno de los caminos que quedan a las moléculas sobrantes o acumuladas es la síntesis de ácidos grasos y grasas. Al menos en algunos animales, como los cerdos, e incluso en algunos humanos, la capacidad de acumulación de grasa es enorme, y se realiza por la transformación de azúcares en ellos. Otro ejemplo es la acumulación de las grasas (aceites) en algunas semillas; estos aceites se forman en las plantas de los azúcares sintetizados originalmente en la fotosíntesis.
Otro caso que podemos imaginar es el de un individuo que no consume carbohidratos, como pudiera darse en ciertas dietas; pero, existe un grupo étnico que prácticamente vive de una dieta de proteínas y carbohidratos: los esquimales. En estos individuos, las grasas se oxidan para obtener energía, pero muchos tejidos requieren de glucosa para funcionar. Su organismo resuelve el problema intensificando algunos caminos metabólicos que permiten la transformación de los aminoácidos en glucosa y otros azúcares.
Dado que muchas de las vías, como la glucólisis, tienen como componente al ADP, algunas de sus reacciones no pueden proceder sin éste, o al menos lo hacen más lentamente si se encuentra en cantidades limitadas. Este ese el caso, por ejemplo, de un individuo que no hace ejercicio físico; con poco ADP, la glucólisis avanza con lentitud y además, los mecanismos de regulación permiten que la glucosa se convierta en piruvato y acetil CoA, que va finalmente a la síntesis de grasas, único grupo de sustancias que se pueden acumular.
Pero hay también caminos metabólicos que en algunos de sus pasos tienen mecanismos muy variados de regulación. En muchas ocasiones, las enzimas, encargadas de alguno de los pasos del metabolismo, son susceptibles de modificar su función, debido a moléculas que en ocasiones están cerca en los esquemas metabólicos; pero otras veces sucede que pueden ser inhibidas o estimuladas por moléculas lejanas o ajenas a la propia vía regulada.
Hay también otros factores que modifican la velocidad de funcionamiento de una vía metabólica; los organismos cuentan también con moléculas diseñadas específicamente para regular muy diversas funciones celulares por diferentes mecanismos. Estas sustancias, producidas por tejidos u órganos específicos, llamados glándulas, son capaces de modificar la función de otras células a distancia que se llaman hormonas.
Las hormonas pueden modificar el funcionamiento de otras células, pero, más específicamente, su metabolismo. Hay algunas, como la insulina, producida por el páncreas de los animales superiores, que tiene la propiedad de estimular el paso de la glucosa a través de las membranas de muchas de las células. De esta manera, la hormona puede regular la glucólisis y demás vías que le siguen, por lo asequible de la primera sustancia que alimenta la vía. Otras hormonas tienen efectos más complicados; pueden actuar directamente, como ciertos esteroides, uniéndose a las moléculas que forman los cromosomas y permitiendo la expresión de ciertas regiones del DNA y con ello la síntesis de determinadas proteínas. Otras cuentan con receptores específicos en las membranas de las células y mecanismos de transmisión de señales resultantes de su presencia y complicadas acciones subsecuentes en el interior de la célula. La adrenalina, por ejemplo, sin penetrar a la célula, a través de su receptor puede producir en el hígado, por una molécula que actúa como su mensajero (llamado segundo mensajero) la estimulación de la degradación del glucógeno almacenado y la liberación de glucosa a la circulación.
Estos casos que mencionamos son ejemplos de los mecanismos de regulación del metabolismo por las hormonas.
Hemos descrito de modo superficial una forma de degradación de la glucosa que parte a la molécula en dos fragmentos iguales, dos moléculas de piruvato o de lactato. Hemos mencionado que esto sucede en el ejercicio muscular intenso, y que permite obtener con rapidez grandes cantidades de ATP. Pero hay diversas variantes de esta vía; tal vez la más importante, por su repercusión en la vida diaria, sea la fermentación. Esta es simplemente una variación de la glucólisis, que dio lugar al estudio del metabolismo hacia finales del siglo pasado, cuando los Büchner señalaron que las células rotas de levadura eran capaces de transformar la glucosa en alcohol etílico.
La diferencia entre la glucólisis y la fermentación estriba en que, en esta
última, prácticamente no existe el paso que convierte al piruvato en lactato,
y en su lugar hay otra enzima que convierte al piruvato en acetaldehido, y otra
a éste en alcohol, como se muestra en la figura II.12.
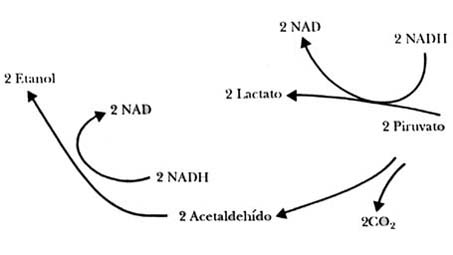
Figura II.2. La fermentación como una variante de la glucólisis.
La importancia de la fermentación estriba en que, en primer lugar, es la vía metabólica responsable de la producción de los cientos de bebidas alcohólicas que existen en el mundo. Independientemente de la importancia económica de las bebidas alcohólicas, el alcohol en sí es un solvente industrial de gran importancia, y se le ha utilizado como combustible sustituto de la gasolina.
Otra característica importante es la obtención de CO2 por la transformación del piruvato en acetaldehido. Al mezclar levadura con masa de harina se producen en ésta burbujas muy pequeñas que durante la cocción se dilatan y permiten fabricar el pan. También es posible modificar la fermentación, interrumpiéndola a medio camino, y lograr la producción de glicerol a partir de la glucosa; este método se utilizó en la primera Guerra Mundial como fuente de glicerol para la producción de la nitroglicerina. Es así que, desde tiempos bíblicos, con el pan y el vino, se ha venido utilizando una variante de una vía metabólica para el beneficio del hombre.
Estos ejemplos de variaciones de una vía metabólica para la producción de materiales útiles para los humanos no son sino una muestra de la enorme diversidad de procedimientos que aprovechan las diferencias naturales o provocadas de las vías metabólicas y que constituyen la base de una gran parte de la moderna biotecnología.