II. LA GENÉTICA
EL HOMBRE, a través de la historia, ha realizado experimentos prácticos utilizando, sin conocerlos, los principios relacionados con la herencia de los caracteres. Por ejemplo, los babilonios desde hace 8 000 años dejaron en sus piedras labradas figuras que representan árboles genealógicos de caballos y esquemas de polinización artificial de dátiles. Por su parte, los chinos mejoraron las semillas de arroz escogiendo las plantas con las características deseadas y haciendo cruzas controladas entre ellas. En América el maíz fue mejorado por selección y los griegos propusieron en sus relatos híbridos raros: por ejemplo, de la cruza entre el dromedario y el leopardo resultaba la jirafa, el camello se originaba del dromedario y el jabalí y el plátano de la acacia y la palma.
La genética como ciencia se inicia formalmente con los principios y reglas analíticas propuestas en 1865 por el monje agustino Gregorio Mendel, quien presentó en la Sociedad de Historia Natural de Brno [actual República Checa] el resultado de sus experimentos acerca de la hibridación en plantas, y postuló las leyes de transmisión de los caracteres hereditarios discretos. Fue una sesión en la que no hubo preguntas, pues sus monumentales contribuciones no fueron comprendidas por sus contemporáneos, a pesar de que Mendel propuso una teoría científica unificadora sobre los mecanismos que rigen los patrones hereditarios de los organismos.
Hoy día la herencia es un fenómeno natural para todos. Sin embargo, se trata de una ciencia joven; el cuerpo de conocimientos que integran a la ciencia que estudia a los genes y su variación, la genética, se han acumulado en este siglo. Las unidades funcionales de la herencia, los genes, constituyen el centro de estudio de la genética, al igual que su transmisión de generación en generación, sus propiedades básicas, las fuentes de variación naturales o artificiales que existen y de cómo se reflejan éstas en los individuos y en las poblaciones.
Estos conocimientos han unificado a la biología moderna, de manera que hoy día no es posible pensar en ningún área de la biología en la que no incida la genética: la unidad de la vida está dada por la estructura química básica del gene y sus funciones, que son muy semejantes en todos los seres vivos. La diversidad biológica puede explicarse al menos en parte por los arreglos peculiares del material genético en cada especie, que son el resultado de la herencia y la variación, y la continuidad de la vida misma se establece al transmitirse el material genético de generación en generación.
Las características biológicas que se observan en los seres vivos, tales como forma, color, talla, capacidad metabólica, funciones bioquímicas, etc., están bajo control genético. Por lo tanto, los genes controlan miles de reacciones químicas que se presentan en los organismos.
Los genes interactúan con el medio para producir un carácter o rasgo, llamado fenotipo. Así, dos individuos con genotipo idéntico (como los gemelos monocigóticos) se desarrollan de manera distinta en dos ambientes diferentes, de la misma manera que dos individuos con genotipo diferente (como los hermanos) se desarrollan de forma distinta en el mismo ambiente.
El genotipo permanece constante durante la vida del organismo. Sin embargo, el fenotipo cambia continuamente en función del tiempo y de la secuencia peculiar de medios interno y externo a los que está expuesto el individuo en su historia particular.
Los seres vivos se clasifican en procariontes y eucariontes, de acuerdo con su organización celular. Los procariontes son organismos unicelulares en los cuales los genes se encuentran localizados en una estructura generalmente circular, desnuda y libre en el citoplasma, llamada cromosoma o genóforo. En cambio, los eucariontes son aquellos seres vivos que presentan en sus células al menos un núcleo rodeado por una membrana nuclear, en el cual queda albergado el material genético que siempre está asociado a proteínas específicas (histonas), formando un complejo que se denomina cromatina.
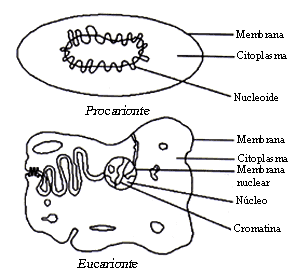
Figura 10. Célula procarionte y célula eucarionte
Los procariontes poseen un solo cromosoma circular o nucleoide y se reproducen mediante una división celular después de que el cromosoma se ha duplicado. Entre los eucariontes, el cromosoma tiene forma de varilla y se presentan más de dos cromosomas en número característico para cada especie. En estos últimos se presentan los fenómenos de dominancia y recesividad debido a que los cromosomas se presentan por pares (diploidia). Los procariontes son haploides, es decir, el genoma es único. En la figura 10 se muestra de manera esquemática una célula procarionte y otra eucarionte.
Los eucariontes pueden estar formados por una o por muchas células. Las formas de división celular pueden ser asexuales, es decir, que un solo progenitor se divide en dos, generándose dos células iguales, o bien a partir de estacas, como ocurre entre las plantas frutales. La forma de reproducción sexual implica la unión de dos gametos o células sexuales provenientes de dos progenitores distintos.
Los eucariontes que presentan reproducción sexual poseen en su organismo células que conforman al cuerpo, o somáticas, en las que se mantiene el número cromosómico característico de la especie, ya que antes de dividirse duplican sus cromosomas, los que se distribuyen en forma igual a las células hijas. Este proceso se llama mitosis, y es distinto a la meiosis, en la cual se forman las células sexuales o gametos. Una célula se divide dos veces, en tanto que el material genético se duplica una sola vez, de manera que a partir de una célula diploide se obtienen cuatro células haploides. Es decir, el número cromosómico característico de la especie queda reducido a la mitad.
La fusión de los gametos masculino y femenino restituye el número diploide de la especie, a partir del huevo o cigoto. Este va dividiéndose por mitosis sucesivas hasta que se produce el nuevo organismo, el cual estará formado por células somáticas diploides y por gametos haploides. Cuando vuelven a unirse los gametos por fecundación se restituye el complemento diploide.
La mitosis es esencialmente un proceso conservativo, ya que a partir de una célula diploide se forman dos células hijas iguales con idéntico complemento cromosómico. La meiosis es el proceso que genera variabilidad entre los organismos de una misma especie; en ella los cromosomas intercambian información genética. Este fenómeno, llamado recombinación, da origen a productos en combinaciones distintas de las parentales; el número y el sitio en el que se producen los intercambios de material genético en los gametos nunca es igual de una meiosis a otra. Este mecanismo permite explicar la razón por la cual no hay dos individuos exactamente iguales, a menos de que se trate, como ya vimos, de gemelos homocigóticos.
LOS ÁCIDOS NUCLEICOS: MACROMOLÉCULAS ESENCIALES.
La vida está formada por varios elementos químicos, entre los que se destacan sobre todo el carbono, el hidrógeno, el oxígeno, el nitrógeno, el fósforo y el azufre. En las células estos átomos forman moléculas simples como el agua y los fosfatos, o bien grandes moléculas constituidas por miles de átomos, como los ácidos nucleicos y las proteínas. En las moléculas, los átomos se mantienen unidos por varios tipos de enlaces, como por ejemplo el iónico, que resulta de la interacción de dos elementos con carga eléctrica opuesta, y el covalente, en el que los átomos comparten electrones.
Los ácidos nucleicos son las macromoléculas en las cuales se almacena y procesa toda la información genética de los sistemas biológicos. En las células de los organismos existen dos tipos de ácidos nucleicos: el ácido desoxirribonucleico (ADN) y el ácido ribonucleico (ARN). Todos los organismos actuales utilizan al ADN como macromolécula informativa, aunque existen muchos virus, como el de la polio y el de la gripa, que usan ARN.
Las moléculas del ácido desoxirribonucleico son, como se muestra en la figura 11, cadenas de nucleótidos formadas por una base nitrogenada, un azúcar, la desoxirribosa y un fosfato.
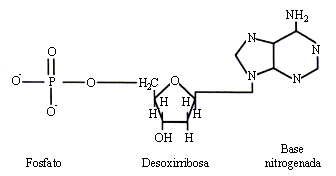
Figura 11. Nucleótido del ADN.
Las bases nitrogenadas pueden ser púricas, como la adenina y la guanina, y pirimídicas, como la citosina y la timina (Figura 12).
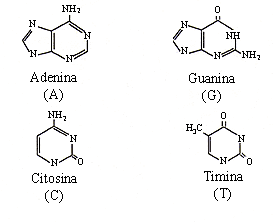
Figura 12. Bases nitrogenadas del ADN.
Aunque la composición básica del ADN se conoció durante algunas décadas, los genetistas de los años cuarenta no podían imaginar cómo este grupo tan limitado de moléculas podría guardar tanta y tan variada información de los seres vivos. Los primeros científicos que propusieron una estructura química coherente con las funciones que realiza el ácido desoxirribonucleico fueron James D. Watson y Francis H. Crick en 1953.
El modelo de la doble hélice describe la conformación de las moléculas del ADN en el espacio. Cada hélice es una cadena de nucleótidos que se mantiene unida a través de puentes fosfodiester, de tal manera que el grupo fosfato forma un puente entre dos grupos OH de dos azúcares adyacentes (Figura 13).
Figura 13. Una hélice o cadena del ADN.
Las dos hebras de la doble hélice son antiparalelas, es decir, están orientadas en dirección opuesta. Las dos hélices se mantienen unidas entre sí por puentes de hidrógeno que se establecen entre las bases nitrogenadas, las cuales se aparean de forma complementaria, tal como ocurre con la llave que abre una cerradura. Es decir, debido a la estructura química de las moléculas, la adenina sólo puede aparearse con la timina mediante dos puentes de hidrógeno, y la citosina con la guanina con tres puentes de hidrógeno (Figura 14). A diferencia de los enlaces covalentes, los puentes de hidrógeno son relativamente débiles, hecho que es muy importante para las funciones que realiza esta macromolécula esencial.

Fig. 14. La dobre hélice y apareamiento entre las bases nitrogenadas por puentes de hidrógeno.
El esclarecimiento de la base química de la herencia sugirió la forma en que la molécula de ADN se replica, ya que cada base especifica a su complementaria (que sólo es una), debido a los puentes de hidrógeno que pueden formarse entre ellas.
La estructura de doble hélice también sugirió que el orden o secuencia en el que se encuentran las bases en el ADN debe dirigir un mensaje codificado que la maquinaria celular debe traducir a un lenguaje distinto, el de las proteínas, que en última instancia están organizadas por los genes.
La replicación del ácido desoxirribonucleico se realiza de acuerdo con un principio muy simple. Cada hebra de la doble hélice sirve como molde para la biosíntesis de una nueva cadena complementaria, proceso que está catalizado por enzimas específicas. Imaginemos a la doble hélice como si fuera una cremallera. Si la abrimos se rompen los puentes de hidrógeno y cada lado correspondería a una hebra de ADN, de manera que las bases quedan expuestas y pueden dirigir la incorporación de sus complementarias; los nuevos nucleótidos se van adicionando (que provienen de una poza de ribonucleótidos que existe en la célula) y la biosíntesis de desoxirribonucleótidos es casi simultánea a la replicación del ADN. Al final de la réplica quedan formadas dos dobles hélices, cada una de ellas con una de las hebras originales (molde) y una hebra recién sintetizada (hija). En la figura 15 se muestra un esquema del proceso.

Figura 15. Replicación del ADN.
La unidad funcional más sencilla en una molécula de ADN es un gene, y no hay que olvidar que las características tanto visibles como fisiológicas de los seres vivos dependen de la estructura y de la expresión precisa de los genes. Los productos de los genes son, por una parte, distintas moléculas de ARN, y por otra, las proteínas, que en las células realizan varias funciones: estructurales, enzimáticas y reguladoras. Las enzimas, y en general todas las proteínas, están formadas por moléculas llamadas aminoácidos, cuyo orden determina su estructura primaria. En los organismos existen 20 aminoácidos esenciales, los cuales se unen por enlaces covalentes, llamados peptídicos, que se forman liberándose agua en la reacción. La conformación de las proteínas en el espacio y su enrollamiento produce estructuras secundarias, terciarias y cuaternarias que crean sitios específicos en los cuales los sustratos se pegan, permitiendo así que ocurran las reacciones enzimáticas. Debido a que los genes codifican la estructura primaria de las enzimas, se puede decir que controlan su función.
La producción de un polipéptido se realiza en la célula mediante un proceso complejo en el que intervienen enzimas y diferentes moléculas de ARN. El mensaje codificado en el ADN es primero transcrito a una molécula intermediaria, el llamado ácido ribonucleico mensajero (ARNm) que lleva la información a los ribosomas. Allí ocurre la traducción del mensaje genético y la unión de los aminoácidos que conforman la estructura primaria de la cadena polipeptídica. Durante el proceso de expresión de los genes intervienen tres moléculas distintas de ácido ribonucleico, y las tres son copias complementarias de secuencias específicas del ADN. El ARN se caracteriza por ser de una hebra generalmente sencilla, su azúcar siempre es la ribosa y en lugar de timina siempre existe otra base pirimídica, el uracilo.
El desciframiento del código genético se llevó a cabo mediante una serie de experimentos elegantes que mostraron inequívocamente que el lenguaje molecular está conformado por cuatro letras, que si se combinan en forma de tripletes, 43, producen 64 palabras, las que pueden ordenarse de manera definida para producir escritos peculiares, las proteínas, que son altamente específicas.
A veces el cambio de un solo aminoácido es suficiente para alterar la función de una proteína. Estos cambios en la arquitectura de las proteínas son producto de las alteraciones o mutaciones en la secuencia de bases de un gene en particular.
Las mutaciones son cambios al azar que ocurren en el material genético y que se heredan a la siguiente generación. Hay mutaciones puntuales, o microlesiones, que se dan entre los genes, los cuales cambian a una nueva forma alélica; también hay mutaciones de la estructura, o del número cromosómico, llamadas mutaciones cromosómicas o macrolesiones.
Las mutaciones puntuales ocurren en forma natural debido a errores poco frecuentes durante la duplicación del ADN, o a daños espontáneos en esta molécula. Los errores ocasionales en el apareamiento de las bases durante la síntesis del ADN ocasionan sustituciones de una base original por otra. Por ejemplo, el desplazamiento de un protón que cambia las propiedades químicas de la molécula, puede llevar a una alteración de la geometría de los puentes de hidrógeno, que quizá ocasione que una guanina se parezca temporalmente a la adenina.
En la conformación normal del ADN la guanina y la timina se encuentran en forma ceto (C=O) y pueden cambiar a la forma enol (COH), lo que se conoce como cambio tautomérico. Por su parte, la adenina y la citosina se presentan normalmente en forma amino (NH2) pero pueden tautomerizarse a formas imino (NH), tal como se muestra en la figura 16.
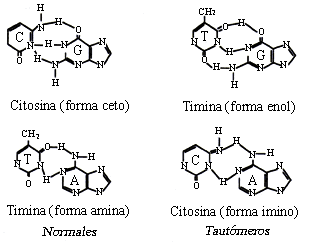
Figura 16. Formas normales y tautoméricas de las bases nitrogenadas.
Este tipo de sustituciones se conoce como transición, de una purina a otra purina o de una pirimidina a otra pirimidina. 1) Al pasar la adenina de la forma amino a la imino se comporta como guanina; el resultado es que frente a la adenina habrá una citosina; 2) cuando la guanina pasa a la forma enol es equivalente a la adenina; por lo tanto, habrá un error al aparearse con la timina; 3) cuando la timina pasa a la forma enol se comporta como citosina; 4) cuando la citosina pasa a la forma imino se comporta como timina; se apareará por lo tanto como adenina (Figura 17).
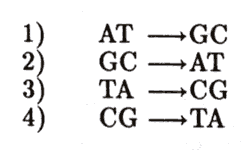
Figura 17. Mutaciones puntuales por sustitución (transiciones).
Durante la replicación puede también sustituirse una base púrica por una pirimídica o viceversa, produciéndose una transversión, tal como se muestra en la figura 18.
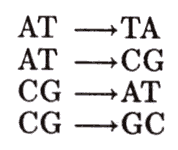
Figura 18. Mutaciones puntuales por sustitución (transversiones).
El origen de las transversiones es menos conocido, ya que la propia conformación espacial de la doble hélice impide este tipo de errores.
Existen otras mutaciones, como las que ocurren por corrimiento del marco de lectura, que se deben a la adición o a la deleción de una o más bases nitrogenadas del ADN durante la replicación (Figura 19).

También hay otras lesiones espontáneas en el ADN, como la depurinización. Ésta se presenta cuando el enlace glucosídico entre la base nitrogenada y la desoxirribosa se rompe, lo cual conlleva a la eliminación de una purina, A o G, del ADN. El resultado es una lesión que se conoce como sitio apurínico (Figura 20).
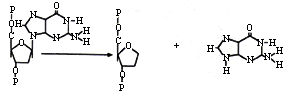
Sin embargo, en los seres vivos existen sistemas enzimáticos de reparación de lesiones del ADN que son muy eficientes y que restituyen rápidamente la base original. En ocasiones, en el sitio apurínico puede insertarse una base distinta, lo que da origen a una mutación puntual. Éstas también se producen por desaminación espontánea, como la que ocurre con la citosina. Al desaminarse esta pirimidina se transforma en uracilo, el que se apareará con adenina, generándose una transición de GC a AT (Figura 21).
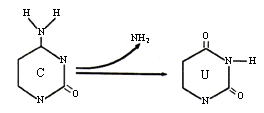
Figura 21. Desaminación de la citosina.
Las mutaciones cromosómicas comprenden cambios en el número o en la estructura de los cromosomas. Este tipo de alteraciones se detectan analizando las células de los eucariontes bajo el microscopio.
En la mayoría de los seres vivos es posible detectar a los individuos que presentan cambios numéricos, porque suelen ser distintos fenotípicamente de los individuos normales. Este tipo de aberraciones se producen por alteraciones durante la separación normal de los cromosomas en el curso de la división celular. En los seres humanos, la falta de un cromosoma, o su presencia en números mayores al diploide, genera severas alteraciones muy frecuentemente asociadas con retraso mental severo, periodos de vida cortos y con esterilidad.
Los cambios en la estructura de los cromosomas son macrolesiones y se producen por rompimientos en los cromosomas, con o sin rearreglos posteriores, lo cual genera alteraciones en la secuencia de los genes, y por lo tanto, en la expresión de los mismos en el organismo.
Así pues, las mutaciones son excepciones que se presentan durante la réplica o la distribución de los cromosomas durante la división celular. En condiciones naturales ocurren al azar, no tienen orientación adaptativa, es decir, ocurren de manera accidental e independientemente del valor que pudieran tener en el ambiente en que viven los organismos.
Las mutaciones son procesos que se presentan en frecuencias muy bajas en las poblaciones naturales y representan la fuente natural de variación biológica. Son la materia prima sobre la cual actúa la selección natural y otros mecanismos evolutivos.
Los cambios en el material genético pueden inducirse por agentes físicos o químicos, llamados mutágenos, que aumentan considerablemente su frecuencia. Los mutágenos químicos han sido utilizados experimentalmente para estudiar los mecanismos mediante los cuales se establecen las mutaciones. Estos mutágenos modelo o de referencia actúan reemplazando a una base, alterando la complementaridad, o dañando al ADN de manera tal que no puede replicarse.
Los análogos de bases son compuestos químicos similares a las bases nitrogenadas, pero con propiedades de apareamiento distintas, de manera que al incorporarse en el ADN producen mutaciones porque durante la duplicación provocan sustituciones de una base por otra. Por ejemplo, el 5 bromouracilo es un análogo de la timina que se presenta en condiciones normales en forma ceto, y por lo tanto se aparea con la adenina; puede cambiar a la forma enólica, adquiriendo así las propiedades de apareamiento de la citosina y generando una transición como se muestra en la figura 22.
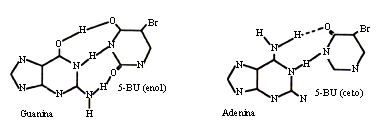
Figura 22. Microlesiones inducidas por un análogo de base.
Los agentes alquilantes son otro grupo de compuestos químicos que tienen la propiedad de añadir radicales alquilo, como metilo o etilo, a las bases nitrogenadas y producir como resultado transiciones, tal como se muestra en la figura 23.
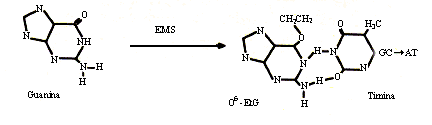
Figura 23. Transición inducida por etilmetanosulfonato.
Los agentes intercalantes constituyen un grupo de moléculas que mimetizan a las bases nitrogenadas originales y que al insertarse entre ellas producen mutaciones de corrimiento de marco de lectura (Figura 24).
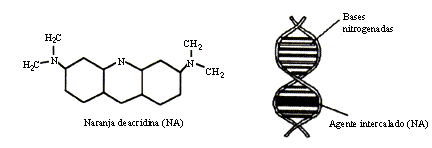
Figura 24. Mutación puntual inducida por un agente intercalante.