IV. COMBUSTIBLE PARA EL MOVIMIENTO: BIOENERGÉTICA
HASTA aquí hemos descrito cómo las células fabrican, siguiendo
las instrucciones contenidas en su código genético, los elementos necesarios
para construir largas estructuras lineales que pueden entrecruzarse o hacinarse
en extensas redes y fuertes armazones internos. La gran resistencia mecánica
de tales complejos estructurales permite a la mayoría de las células mantener
una forma definida y característica, a la vez que brinda puntos de apoyo para
que se ejerza y transmita el movimiento. Es por esta razón, como hemos dicho,
que al conjunto organizado de microtúbulos, filamentos de actina y filamentos
intermedios se le ha dado el nombre de citoesqueleto. Pero, salvo en los cuentos
de terror, los esqueletos no se mueven por sí mismos, y el esqueleto de la célula
no es la excepción de la regla. Por tanto, la siguiente etapa de nuestro análisis
la destinamos a considerar qué es lo que mueve al citoesqueleto, actividad de
la cual depende la generalidad de los movimientos de las células.
Por supuesto, cualquier cambio en la organización del citoesqueleto, sea por la adición de una nueva red estructural, sea por la ampliación, conversión o desmantelamiento de otra ya existente, o por una combinación de todas estas causas, puede dar como resultado una alteración en la forma de la célula. Esta clase de reacomodo es sin duda un tipo de movimiento, en la medida en que lo son también el hundimiento de la carpa de un circo cuando se elimina uno de sus postes de sostén, o las dramáticas metamorfosis que experimenta un paragüas cuando cambian de posición sus varillas. Más adelante veremos cómo algunas células —como las amibas y los glóbulos blancos de la sangre, que de continuo cambian de forma, o los espermatozoides de ciertos invertebrados marinos, en los que una súbita polimerización de la actina produce un largo tentáculo que les permite anclarse al óvulo— muestran ejemplos notables de esta suerte de movimiento. Sin embargo, la sola transformación de estructura no es suficiente para explicar la vasta diversidad de movimientos que se observan en la materia viviente. La contracción activa del cuerpo celular, la agitación de apéndices en su superficie, y el transporte de materiales por el interior de las células, que en general depende estrictamente de los microtúbulos o de filamentos de actina, requieren además de otros componentes indispensables.
El movimiento celular es producto de la actividad de una complicada maquinaria compuesta a su vez por diferentes tipos de máquinas individuales, distribuidas y arregladas conforme a distintos diseños, según el tipo de célula y de movimiento de que se trate. Ahora bien, toda máquina consta de algunas partes fijas que sirven de soporte y de las partes móviles que ejecutan la función. Pero, si nos enfocamos a examinar las máquinas que son capaces de movimiento propio, es decir; que no son accionadas por el esfuerzo corporal del hombre o de algún animal, es fácil percatarse de que por lo común las piezas móviles comprenden dos categorías: aquellas que únicamente son jaladas o empujadas, y las que se encargan de originar el movimiento. Estas últimas son los motores, que convierten alguna forma de energía —térmica, química, hidrodinámica, eléctrica, nuclear, etc.— en energía mecánica. La maquinaria que produce el movimiento celular tiene también sus motores. Al igual que todo esqueleto, el citoesqueleto depende de elementos generadores de fuerza para efectuar los cambios de posición que determinan el movimiento. Hace no muchos años se acuñó el nombre de citomusculatura para referirse al conjunto de tales factores clave para el cinetismo biológico. En la actualidad el término ha caído en desuso, pero el concepto gana validez día con día, a medida que se descubren nuevas proteínas especializadas en convertir energía metabólica en fuerza mecánica. Son estos potentes motores moleculares los que, con apoyo en el citoesqueleto, impulsan a los seres vivos. El movimiento de las células es por tanto activo, producto de los componentes del propio citoplasma y se denomina motilidad para distinguirlo de movilidad, es decir, de movimientos debidos a causas externas. Antes de describir sus mecanismos de operación, conviene mencionar brevemente de dónde obtienen la energía que utilizan.
La energía que mueve al mundo viviente proviene del Sol. La energía solar es
capturada por algunos organismos —ciertos microbios y las plantas verdes—
que la aprovechan directamente para construir, a partir de materiales inorgánicos
simples que absorben del medio, las complejas moléculas orgánicas que los constituyen
(Figura lV.1) El resto de los seres vivientes, que no tienen la facultad de
utilizar directamente la energía solar de esta manera, dependen de un proceso
inverso para funcionar. Ingieren las moléculas orgánicas creadas por el proceso
anterior y las desarman hasta convertirlas otra vez en sustancias simples, con
lo que se libera la energía invertida para la construcción de las primeras.
Los motores que mueven a las células, al igual que el resto de los mecanismos
fisiológicos, consumen la energía derivada del Sol, tanto por la vía directa
como por la indirecta. Por cualquiera de ambas vías dicha energía queda almacenada
temporalmente en un combustible de uso general llamado trifosfato de adenosina,
al que los biólogos acostumbran referirse por sus siglas en inglés: ATP.
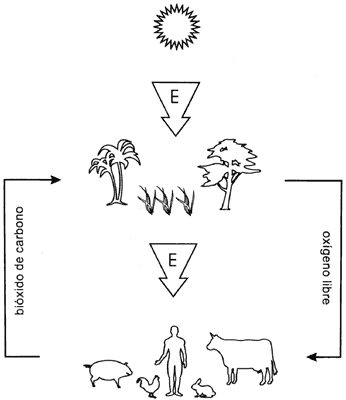
Figura IV. 1. Utilización de la energía solar por los seres vivos.
El ATP es un nucleótido idéntico a uno de los cuatro eslabones
que componen cada una de las cadenas del ADN, excepto que en lugar
de un solo grupo fosfato tiene tres, los dos últimos conectados en serie con
el primero (Figura IV.2). Su propiedad de servir como intermediario en el flujo
de energía en las células estriba en que el tercer grupo fosfato se encuentra,
por así decir, acomodado a presión en la molécula. Un ejemplo mecánico puede
ayudar a explicar de qué manera el ATP guarda energía que es capaz
de transferir a otras moléculas para promover una reacción bioquímica.
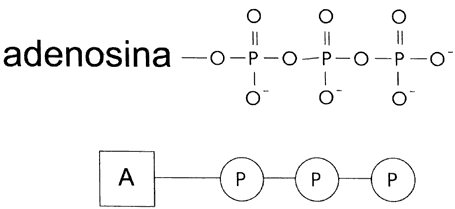
Figura IV. 2. Fórmula del ATP, la fuente principal de energía
dentro de la célula.
Imaginemos una caja de madera en la que se empacan tres balones de hule, aunque únicamente caben dos sin dificultad; el tercero alcanza a entrar sólo parcialmente, por lo que para cerrar la caja es necesario oprimir la tapa y asegurarla con un broche. La elasticidad de los balones les permite deformarse para ser encajonados si se les fuerza a ello, pero estarán listos para tomar su forma original a la primera oportunidad.
Para construir este sistema se ha requerido invertir la energía en varios momentos:
en primer lugar, hay que considerar la energía que tomó la producción de los
materiales básicos (digamos, el crecimiento de los árboles que proveyeron la
madera para la caja y el caucho para los balones); se tiene además la energía
gastada para la confección de cada uno de los elementos del sistema (fabricar
la caja y los tres balones); por último, está la energía aplicada para integrar
el sistema con base en los elementos (colocar en la caja los tres balones y
cerrarla venciendo la resistencia que resulta de la presencia del tercer balón).
(Figura IV.3).
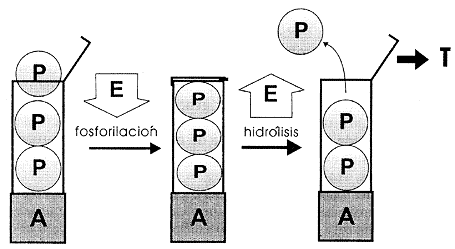
Figura IV. 3. Modelo mecánico para ilustrar la formación del enlace de alta
energía en el ATP durante su fosforilación y la liberación de esta
energía por la hidrólisis del enlace. La liberación de energía se aprovecha
para realizar un trabajo T. Gran parte del trabajo en una célula se expresa
como movimiento.
La energía invertida en este sistema no es recuperable por completo, pero sí una buena parte de ella. Por ejemplo, un método para extraer del sistema el máximo de energía recuperable es prenderle fuego, lo que produce una generación de calor que se puede aprovechar para realizar trabajo si la combustión ayuda a vaporizar agua en una caldera y la presión del vapor se emplea para poner en movimiento una máquina. Sin embargo, a pesar de que este procedimiento es muy efectivo, implica la destrucción total del sistema, lo que no siempre es deseable. Una manera menos drástica de recuperar parte de la energía contenida en el sistema consiste simplemente en desabrochar la tapa de la caja; los tres balones volverán a su forma original y uno de ellos saldrá disparado, empujando la tapa tan pronto como desaparezca la barrera que los mantenía aprisionados en el interior. La fuerza desarrollada espontáneamente por el sistema, cuando se relaja, también podrá realizar un trabajo; por ejemplo, impulsar hacia arriba un objeto relativamente ligero colocado encima de la tapa. Parte de la energía almacenada habrá sido transferida al objeto, que gracias a ella efectúa un movimiento ascendente a pesar de la atracción gravitacional. Esta energía fácilmente liberable es una consecuencia exclusiva de la presencia del tercer balón, porque sin él la energía restante del sistema no realizaría el trabajo de manera espontánea. Por otra parte, el sistema puede ser puesto en servicio en múltiples ocasiones, pues para ello basta con introducir nuevamente el tercer balón en la caja, oprimir la tapa y abrocharla.
Una situación equivalente existe en el ATP. Al igual que cualquier
otra molécula, contiene energía —la invertida en la creación y el ensamble
de sus componentes. La descomposición total de la molécula liberaría el máximo
de energía recuperable, pero las células prefieren aprovechar sobre todo su
capacidad de guardar energía fácilmente liberable por la presencia del tercer
grupo fosfato, que se encuentra agregado de manera forzada en la estructura.
Desde luego, en este caso no se trata de una compresión mecánica, sino de una
posición desfavorable a la estabilidad de la molécula, debida principalmente
a repulsiones eléctricas entre átomos vecinos. Sin embargo, el sistema opera
de manera semejante al modelo descrito. Tan pronto como se abre la posibilidad,
el tercer grupo fosfato se desprende del conjunto y la energía liberada es aprovechable
para efectuar una alteración que no ocurriría de manera espontánea en moléculas
cercanas. De esta manera el ATP queda convertido en ADP
(difosfato de adenosina), que puede ser "recargado" nuevamente si, a expensas
de una fuente extra de energía, se le añade otra vez un tercer grupo fosfato
(Figura IV.4).
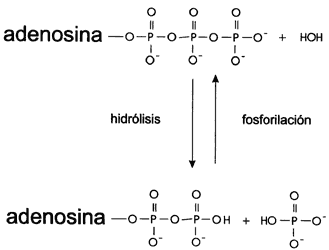
Figura IV. 4. Formación del ATP y su hidrólisis.
A diferencia de los balones del ejemplo mecánico anterior; la división del
ATP implica la ruptura de un enlace químico entre el segundo grupo
fosfato y el tercero. Los dos cabos sueltos de este enlace tienen cargas eléctricas
de signos contrarios y tienden a asociarse de manera natural con algo que las
neutralice; para ello utilizan los dos fragmentos en que puede partirse una
molécula de agua (H+ y OH-). A este fenómeno se le denomina
hidrólisis (del griego hydros agua y lysis, descomponer), por
lo que se dice que el ATP es hidrolizado cuando se divide en ADP
y fosfato libre. En la célula, la hidrólisis del ATP es promovida
por enzimas llamadas ATPasas, que al mismo tiempo capturan la energía
liberada para aplicarla a un fin particular. Cuando, como veremos, la aplicación
consiste en que la ATPasa modifica transitoriamente su propia conformación
para realizar un trabajo mecánico, se le da el nombre de mecanoenzima. Otras
ATPasas utilizan la energía derivada de la hidrólisis del ATP
para distribuir solutos de manera asimétrica entre ambos lados de las membranas
celulares, por lo que constituyen un verdadero sistema de bombeo. Muchas otras
ATPasas aplican la energía para vincular moléculas durante la construcción
de cómpuestos orgánicos mayores.
La adición de un grupo fosfato a una molécula de ADP para regenerar
ATP se conoce como fosforilación. Como mencionamos en un párrafo
anterior, en el comienzo del flujo energético a través de los sistemas vivientes
la energía necesaria para incorporar el tercer fosfato proviene del Sol. La
luz es energía radiante capaz de actuar sobre la materia, como lo atestiguan
todos los recursos que ofrece la fotografía, el cambio de tono de la piel después
de asolearse, y la posibilidad de incendiar una hoja seca mediante una lente
que concentre en un punto los rayos solares. Las plantas verdes y algunos microorganismos
tienen la facultad de capturar esta energía y aprovecharla para producir ATP.
Este fenómeno, llamado fotofosforilación, tiene lugar gracias a la existencia
de sustancias que son excitadas por la luz, como la clorofila (Figura IV.5).
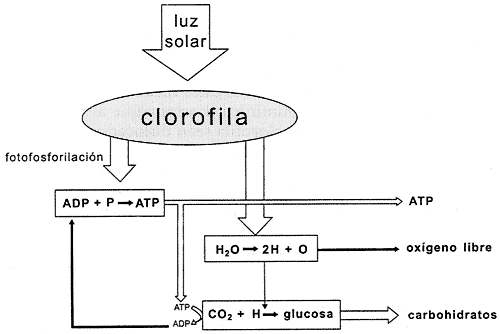
En las plantas dichas sustancias se encuentran enclavadas en las membranas internas de numerosos corpúsculos citoplásmicos llamados cloroplastos. Cuando éstos reciben la radiación, la incidencia de la energía lumínica ocasiona que algunos electrones de los átomos de la clorofila sean dislocados de su posición habitual y se desplacen hacia moléculas contiguas de otro tipo, que a su vez los descargan en otras moléculas, y éstas los hacen pasar a otras más, a lo largo de una compleja cadena dentro de la propia membrana de que forman parte. La energía de este flujo de electrones es empleada para acumular iones de hidrógeno en uno de los dos espacios separados por la membrana. Al verse más concentrados en uno de los compartimientos, los iones de hidrógeno tienden naturalmente a difundirse hacia el lado contrario buscando alcanzar el equilibrio. Y sucede que el único camino para el regreso a través de la membrana es una enzima que saca provecho del empuje de los iones de hidrógeno en su retorno para unir un grupo fosfato al ATP. Estas enzimas se denominan ATP- sintetasas.
Una fracción de los electrones que la luz desprende de la cloroflia regresa
a ésta por una ruta molecular alterna, pero el resto termina por abandonar la
membrana, uniéndose a compuestos solubles que los aceptan. Esta segunda parte
tiene que reponerse de algún modo, puesto que de no ser así el proceso se interrumpiría
rápidamente al agotarse los electrones que pueden ser dislocados de la clorofila.
La fuente inagotable de electrones que mantiene en marcha el mecanismo es el
agua, que los cede a la clorofila. El resultado total es la descomposición de
la molécula de agua en dos protones y un átomo de oxígeno. La inmediata asociación
de dos de estos átomos produce oxígeno molecular; que puede difundirse a través
de las membranas del cloroplasto hacia otras regiones de la célula e incluso
más allá, fuera de la planta, hacia la atmósfera. Por su parte, los electrones
que emergen de la membrana, asociándose con aceptores solubles, proveen la energía
necesaria para la fabricación de carbohidratos o azúcares a partir de bióxido
de carbono, que los vegetales absorben de la atmósfera. A través de estos dos
procesos paralelos, que en conjunto reciben el nombre de fotosíntésis, la energía
solar queda atrapada en forma de ATP y compuestos orgánicos, con
lo que ingresa en el mundo viviente, donde es distribuida por medio de las cadenas
alimentarias. La nutrición de unos seres vivos a expensas de otros implica no
sólo la obtención de materias primas para crecer y resarcir el desgaste natural
de los componentes corporales, sino también —y de manera capital— la
apropiación de la energía indispensable para dicho mantenimiento y para la reproducción.
La degradación de los materiales orgánicos ingeridos, en particular de los
carbohidratos, constituye el principal recurso energético con que cuentan los
animales y otros organismos incapaces de efectuar la fotosíntesis. Dicha degradación
se lleva a cabo con la participación de numerosas enzimas que conducen paso
a paso el proceso, y puede seguir dos vías principales. La más sencilla y antigua,
aunque menos eficiente, consiste en fragmentar moléculas de algunos azúcares
como la glucosa, y se denomina por tanto glucólisis (Figura IV.6). La división
de una molécula de glucosa libera energía suficiente para la generación de dos
moléculas de ATP, a partir de ATP y de fosfato inorgánico.
Si bien esta clase de reacción se encuentra representada de manera casi universal
en los organismos, su rendimiento energético es bajo en comparación con la cantidad
total de energía recuperable que existe en la glucosa. No sorprende, por tanto,
que la evolución haya favorecido el desarrollo y la expansión de la segunda
vía de degradación de carbohidratos, en general subsecuente a la primera, que
permite una explotación más completa de su potencial energético.
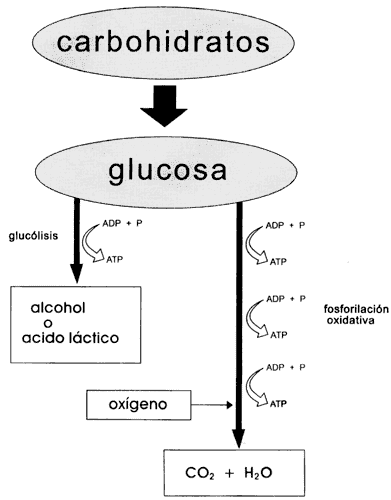
En este último proceso la glucosa es descompuesta a través de una completa
serie de reacciones enzimáticas hasta revertir a sus ingredientes originales,
es decir, el bióxido de carbono y el agua que las células vegetales utilizaron
durante la fotosíntesis. Buena parte de la energía solar que hizo posible dicha
transformación es retenida mediante la fosforilación de 36 moléculas de ADP
por cada molécula de glucosa degradada totalmente. Este método de reconstitución
del ATP requiere de oxígeno, cuya combinación con el carbono de
la glucosa para dar como resultado terminal el bióxido de carbono constituye
una oxidación, por lo que se ha dado el nombre de fosforilación oxidativa. Es
indispensable un continuo abastecimiento de oxígeno para mantener en operación
este eficiente proceso de extracción de energía metabólica. Los animales superiores
cuentan con un aparato respiratorio y un sistema circulatorio que colaboran
en la tarea de ingresar y distribuir el oxígeno en sus cuerpos. Sin embargo,
la respiración es un fenómeno mucho más general, que ocurre en la gran mayoría
de las especies animales y vegetales. El sitio donde tiene lugar la respiración
esta en el interior de las células, en las membranas de pequeños corpúsculos
funcionalmente similares a los cloroplastos: las mitocondrias.
El paralelismo inverso evidente entre la fotosíntesis y la respiración se refleja
en una clara similitud de sus mecanismos. La degradación metabólica de carbohidratos
y otros compuestos orgánicos aporta electrones que son conducidos a lo largo
de una cadena de moléculas enclavada en la membrana mitocondrial, que termina
por cederlos al oxígeno con la consiguiente producción de agua. El movimiento
de los electrones brinda la energía para un bombeo de iones de hidrógeno a través
de la membrana, y el empuje de dichos iones para redistribuirse de manera uniforme
a ambos lados de la misma impulsa la operación de una ATP-sintetasa.
La energía del ATP es aprovechada después por diversos tipos de
ATPasas, entre las que se encuentran las mecanoenzimas o motores
moleculares, cuyo funcionamiento examinaremos a continuación.