XII. TECTÓNICA, DERIVA Y BIOLOGÍA
LA REVOLUCIÓN moderna de la tectónica global ha contribuido considerablemente al cambio de la visión de los biólogos con respecto a la evolución y distribución geográfica de los organismos. Vimos hasta ahora que el proceso de tectónica de placas puede unir continentes en un único supercontinente (por ejemplo, Pangea), también puede fragmentar un supercontinente en diversos bloques continentales menores, y generar grandes cadenas de montañas por la colisión de placas litosféricas. Las respuestas de la biota y sus organismos a esos procesos son, en el primer caso, la dispersión y creación de simpatría, esto es, la superposición de las áreas geográficas de distribución de distintas especies; en el segundo caso, la especiación, generada por la fragmentación de las poblaciones antecesoras en poblaciones descendientes que están incapacitadas para intercambiar información genética y que divergen en consecuencia de ello, pues mantiene poblaciones aisladas por barreras físicas. Veamos algunos ejemplos de esos procesos.
ALTERACIONES EN LAS CONDICIONES DEL AMBIENTE MARINO
Según Valentine y Moores (1972), el proceso de la tectónica afecta directamente la distribución geográfica de los organismos marinos por medio del surgimiento de barreras, alteraciones en el patrón de temperatura de las aguas, y modificaciones en las corrientes oceánicas y en la estabilidad vertical de las columnas de agua. Las alteraciones en el nivel del mar (transgresiones/regresiones) también son un importante reflejo de los procesos de la tectónica de placas, debido a las modificaciones en la capacidad volumétrica de las cuencas oceánicas o al desplazamiento de los continentes hacia posiciones más cercanas a los polos. Normalmente, los acontecimientos regresivos están asociados a la coalescencia de los continentes, mientras que los transgresivos están ligados a su fragmentación (Windley, 1978). Por ejemplo, durante los episodios de expansión acentuada del piso oceánico, el surgimiento de las dorsales meso-oceánicas, que poseen un volumen considerable, condujo a la formación de máximos topográficos. Como respuesta, las aguas oceánicas fueron desplazadas, causando transgresión en los continentes. Contrariamente, cuando el proceso de expansión cesa, las dorsales entran en subsidencia, lo que aumenta el volumen de las cuencas oceánicas —hay entonces una "ascención" de los continentes por la regresión de las aguas. De acuerdo con Valentine y Moores (1970), el volumen actual de las dorsales meso-oceánicas es de aproximadamente 2.5 x 10 8 km³, suficiente para afectar el nivel del océano en casi 0.5 km alrededor de todos los continentes. Durante episodios de regresión habría eliminación de hábitat de aguas rasas, con el consiguiente aumento en la competencia por espacio y nutrientes, y la disminución de formas diversas o incluso, en algunos casos, la extinción de muchos organismos marinos. Por supuesto, un patrón opuesto se verifica en casos de episodios de transgresión. Es curioso señalar que la extinción en masa al final del Pérmico coincide con la coalescencia de los continentes en Pangea (figura 46). Por otro lado, la diversificación de las faunas marinas del Mesozoico y Cenozoico coincide con la ruptura de ese supercontinente (figura 46).
ALTERACIONES EN EL AMBIENTE TERRESTRE
En el ambiente terrestre, la unión de placas tectónicas a veces permite un
intercambio entre faunas antes aisladas. Un ejemplo clásico de ese fenómeno
se encuentra en la ligazón de Sudamérica y Norteamérica, al final del Plioceno,
a través del establecimiento del istmo de Panamá, probablemente debido al desplazamiento
de la Placa de Cocos, en el Pacífico. Según la hipótesis tradicional, los mamíferos
sudamericanos se habían mantenido aislados durante casi todo el Terciario, mientras
que los de Norteamérica mantenían intercambio con los de Eurasia a través del
estrecho de Bering. Cuando se estableció la comunicación entre Norteamérica
y Sudamérica hubo gran intercambio de especies, estableciéndose simpatría entre
ellas en toda América, con la subsecuente competencia y eliminación de diversas
formas. Muchas familias y órdenes enteros de ambas faunas se extinguieron, de
tal manera que el número total de especies disminuyó extraordinariamente. Tales
extinciones parecen relacionarse no sólo con la competencia, sino también con
grandes cambios físicos que ocurrieron en el ambiente.
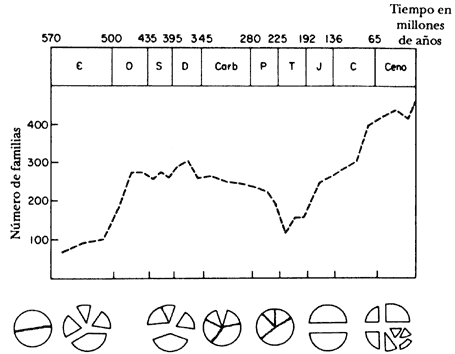
Figura 46. Correlación entre los índices de diversidad de la fauna con los
patrones de coalescencia y fragmentación continental, durante el Fanerozoico
(Valentine y Moores, 1970).
EL CASO ESPECTACULAR DE LA LARGA JORNADA DE LAS TORTUGAS VERDES
Un caso espectacular de cómo la moderna teoría de la tectónica de placas puede iluminar ciertos aspectos de la biología es el de la migración de ciertas tortugas marinas. Normalmente, las tortugas continentales (lacustres o terrestres) ponen sus huevos en nidos situados en las áreas donde viven. Sin embargo, las áreas de alimentación y anidación de las tortugas marinas se encuentran muy distantes de su domicilio habitual. Dicen Pough et al. (1993) que: "...la capacidad que tienen las tortugas de navegar por miles de kilómetros de océano y encontrar su camino hasta sus playas de anidación, que pueden consistir en sólo un poco más que estrechas ensenadas de una pequeña isla, es un fenómeno extraordinario."
El ejemplo más notable de ese fenómeno lo brindan las tórtugas verdes (Chelonia mydas) que anidan en la isla de la Ascensión, cuya superficie es minúscula (cerca de 90 km²) y se encuentra perdida en medio del Atlántico, a 2 200 km al este de la costa de Brasil y próxima a la línea del ecuador (figura 47). La población de tortugas de la Ascensión se alimenta en la costa de Brasil, pero gran parte de los mecanismos de orientación que utilizan para encontrar sus zonas de anidación son desconocidos; esto ocurre en los meses de diciembre a marzo, cuando cruzan fuertes corrientes ecuatoriales y evitan los promontorios y las escarpas rocosas que protegen la costa de la pequeña isla.
Si en la costa de Brasil existen centenares de playas, ¿por qué esos animales eligen las playas arenosas de la isla de Ascensión para hacer sus nidos? ¿Cómo pueden saber que existe esa islita a medio Atlántico de distancia? El hecho de que la hayan elegido es intrigante (Pough et al., 1993) y ha sido, por más de 30 años, objeto de la investigación del geólogo Archie Carr, de la Universidad de Florida. Según Carr y Coleman (1974), las tortugas verdes de la isla de la Ascensión heredaron un comportamiento que ha evolucionado desde hace casi 70 millones de años, cuando el Atlántico sur se originó, luego de la separación de las placas de Sudamérica y África. Posibles antecesores mesozoicos de la actual tortuga verde se encuentran en los depósitos sedimentarios de Brasil.
Los continentes se han separado a una tasa de 2 cm por año y la cresta oceánica central, localizada entre ellos, ha generado una sucesión de islas volcánicas. La Ascensión, que tiene alrededor de 7 millones de años de edad, es la más reciente de esas islas. Una montaña marina, ubicada cerca de 15 km hacia el oeste de la isla de la Ascensión y que ahora se encuentra sumergida a una profundidad de 1 500 m, probablemente representa un remanente de la isla anterior. De acuerdo con la teoría de Carr, las tortugas verdes habían empezado a anidar en las primeras islas volcánicas desde 70 millones de años atrás, cuando ese comportamiento exigía una jornada de apenas unos 300 km a través del mar. A medida que la separación entre Sudamérica y África proseguía las islas viejas se erosionaron y hundieron en el mar, y nuevas islas se generaron a partir de la cresta oceánica. [...] Obviamente, cada isla nueva ha estado más distante de la costa brasileña que las islas anteriores, porque el Atlántico sur se ha ensanchado; hoy día, las tortugas emprenden una jornada de más de 2 000 km desde sus regiones de alimentación hasta las playas donde anidan (Pough et al., 1993).
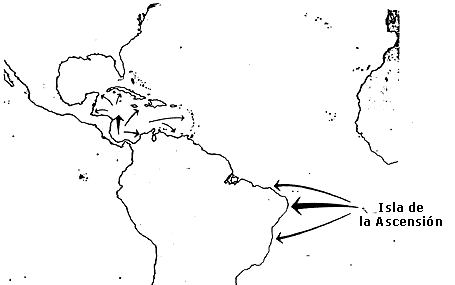
Figura 47. Movimientos migratorios de las tortugas verdes marinas (Chelonia
mydas)en el Caribe y en el Atlántico Sur. La población que anida en las
playas del Caribe es atraída a las regiones de alimentación del Caribe y el
Golfo de México. Las tortugas que lo hacen en la isla de la Ascención se alimentan
a lo largo de la costa norte de Sudamérica (según Pough et al., 1993).
Si la hipótesis de Carr es correcta, el proceso de migración de esos animales es la confirmación viva de la expansión del piso oceánico (Eicher y McAlester, 1980).
LA BIOGEOGRAFÍA POR VICARIANZA
El postulado fundamental de la biogeografía por vicarianza es que Tierra y vida evolucionan juntas. En la figura 48 vemos cómo se procesa esta idea teóricamente.
Supongamos que en un horizonte de tiempo T1 existiera cierta especie de organismos bisexuados de fecundación cruzada, que nombraremos G, ocupando cierto supercontinente S11. Supongamos en seguida que ese supercontinente S11 se dividió, generando los continentes S10 y S1. La población de la especie G se divide simultáneamente en dos poblaciones descendientes, A (que ocupa el continente S1) y H (que ocupa el continente S10). Como A y H quedan aisladas por el surgimiento de una barrera física que no les permite intercambiar información genética, cualquier mutación que ocurra posteriormente en A (y que no sea eliminada por la selección natural) no podrá transmitirla a H, y viceversa. Con el tiempo, por la acumulación de mutaciones, las dos especies A y H se diferenciarán morfológicamente. Supongamos que el continente S1 nunca más volvió a dividirse, llegando hasta el presente (horizonte de tiempo T6); la especie A, en consecuencia, continúa como una sola entidad, pero pueden continuar ocurriendo mutaciones y así acentuarse la diferenciación morfológica. A su vez, la especie H, en un tiempo posterior T2 se dividió al partirse el continente S10 en otros dos continentes, S8 y S9. Así, H genera dos poblaciones descendientes, J (que ocupa el continente S8) e I (que ocupa el continente S9); con el tiempo también pueden adquirir diferencias morfológicas que las distingan una de otra. El proceso puede continuar indefinidamente —toda vez que se divida un continente, la población de organismos que lo ocupa se divide con él y se diferencia. Así, J, en el horizonte de tiempo T4, respondió a la partición de S8 en dos continentes, S7 y S4, por especiación, generando las especies K y D. La especie I, en el horizonte de tiempo T3, se dividió junto con la partición del continente S9 en los continentes S3 y S2 resultando las especies descendientes C y B. Estas llegaron hasta el presente sin subdividirse, ya que sus respectivas áreas, S3 y S2, permanecieron íntegras. En el horizonte de tiempo T5, finalmente, el continente S7 ocupado por la especie K, se dividió en otros dos, S6 y S5; así, con base en K, por especiación, se generaron las especies F y E.
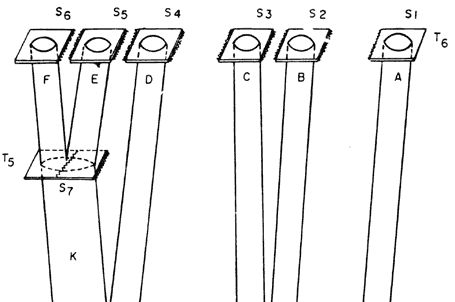 |
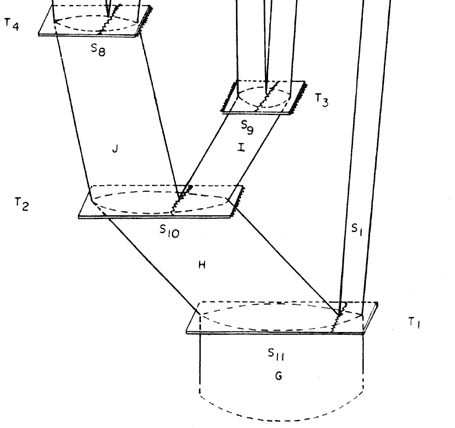 |
Figura 48. Esquema teórico de la coevolución de la Tierra y de la vida.
La figura 49 representa la distribución de las especies A, B, C, D, E y F en
el presente. Es obvio que, cuando miramos esa figura, sólo vemos el patrón de
distribución geográfica actual, pero no hay que olvidar que es el resultado
final de un largo proceso evolutivo. ¿Cómo, a partir de ese resultado final,
podemos recuperar la historia de la evolución de ese grupo y compararlo con
la historia de la Tierra? Con la ayuda de otros conceptos y métodos.
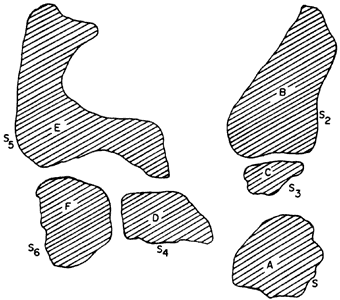
Figura 49. Distribución geográfica de las especies A, B, C, D, E Y F.
Estudiemos inicialmente cómo se establece la genealogía de un grupo, o sea,
las relaciones de parentesco entre especies. Examinemos las figuras 50 y 51.
Supongamos una población P que desde hace cierto tiempo (T) existe en un área
S (figura 50 (a). Supongamos también que un determinado carácter de los organismos
pertenecientes a P cambie de estado, por mutación, y que pase al patrimonio
genético de todos los organismos P, por ejemplo, que pase de un estado c a un
estado c'. Al estado c, que antecede a c' en el tiempo denomina estado plesiomórfico;
al estado c', que sucede a c en el tiempo, estado apomórfico. Nótese
que c' (el estado apomórfico) representa una novedad evolutiva para P;
como pertenece sólo a P (y, supongamos, a ninguna otra especie) lo denominamos
entonces una autapomorfía de P. El estado c' caracteriza necesariamente
a P, después de que se establece. Podemos representar la misma idea por medio
de un diagrama, como en la figura 51(a). Ahí están representados tres caracteres,
a, b y c'; c' es una autapomorfía de P; a y b están en el estado plesiomórfico.
Imaginemos ahora que el área S se modifique —puede partirse en dos, o puede
surgir una novedad evolutiva (como en el ejemplo de la figura 50, una cadena
de montañas). Sea por partición del área S en otras dos, o por el surgimiento
de cualquier otra barrera física (ocasionada por el magma o por razones meteorológicas),
la nueva barrera ocasionará que la población P se divida en dos poblaciones
descendientes P1 y P2 (ninguna de las cuales será más
la población antecesora total P). Eso representa un evento de vicarianza.
La nueva barrera impedirá el flujo génico entre las poblaciones P1
y P2; c' fue heredado de P tanto por P1 como por P2;
c' ahora se denomina como una sinapomorfía estricta de P1
y P2, pues provienen del antecesor inmediato de P1 y P2.
Una sinapomorfía estricta indica que dos (o más) especies son estrictamente
monofiléticas, o sea que provienen de un único antecesor común inmediato.
La figura 50(b) representa la partición de la población P en dos poblaciones
descendientes P1 y P2, separadas por el surgimiento de
una montaña. La figura 51(b) muestra diagramáticamente la situación de las ramas
y la sinapomorfía c'. Nótese que, desde el punto de vista morfológico, en esta
fase, P1 y Pv son idénticas; sólo las diferenciamos por ocupar áreas
disyuntas (respectivamente S1 y S2) Postulemos ahora que,
en la población P1, un determinado carácter cambie de estado; por
ejemplo, que a pase al estado a'; a es el estado plesiomórfico (anterior en
el tiempo) y a' el estado apomórfico; a' representa una autapomorfía para P1;
como existe una barrera física interpuesta entre P1 y P2,
esa mutación quedará restringida a P1, pues tal barrera le impide
pasar hacia P2. Imaginemos que una cosa semejante ocurra con P2—un
determinado carácter pasa del estado b al estado b' que será una autapomorfía
de P2, y será exclusivo de esa población, por no poder pasar a P1,
debido a la barrera (figura 50(c)). Tenemos entonces que P1 y P2,
después de esos procesos, serán diferentes desde el punto de vista morfológico.
La figura 51 (c) ilustra dos especies terminales, P1 y P2,
cada cual con su autapomorfía (respectivamente a' y b'), y ambas con la sinapomorfía
c'.
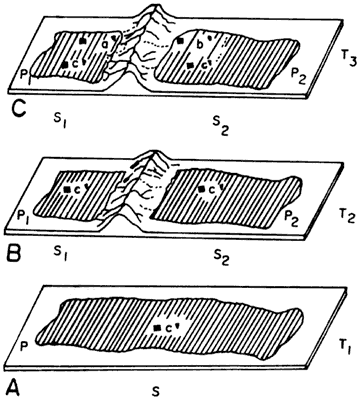
Figura 50. Un caso simple de especiación por vicarianza, con la subsecuente
diferenciación morfológica de las especies resultantes.
A la oposición de por lo menos un par de autapomorfías se le da el nombre de heterobatmia ("diferentes peldaños", en griego). La heterobatmia nos permite reconocer dos especies como morfológicamente distintas. Ya hemos visto que una sinapomorfía estricta de dos o más especies nos indica que son estrictamente monofiléticas, o sea, que descienden de un único antecesor inmediato. Si dos (o más) especies presentan heterobatmia y son estrictamente monofiléticas las llamamos especies hermanas. En el caso de la figura 51(c), P1 y P2 son claramente especies hermanas. Estos son, de manera muy general, los fundamentos de la teoría de la sistemática filogenética, propuesta por Willi Hennig (1968).
Supongamos ahora que P1 ocupe en la actualidad el área S1,
y P2 el área S2, como se muestra en la figura 52(b). Al
estudiar su morfología determinamos que presentan heterobatmia y que son estrictamente
monofiléticas (figura 51(c)). Entonces podemos proponer la hipótesis de que
sus áreas de distribución fueron separadas en el pasado por alguna barrera física.
En consecuencia postulamos que su antecesora, P, ocupó un área que es la unión
de S1 y S2, o sea, una única área antecesora S (figura
52(a)), si es que el proceso de especiación fue por vicarianza (aislamiento
geográfico). Nótese que P1 y P2 reemplazan o sustituyen
a P, así como S1 y S2 reemplazan o sustituyen a S (en
el tiempo); por eso la teoría se llama vicarianza (del latín vicarius,
sustituto).
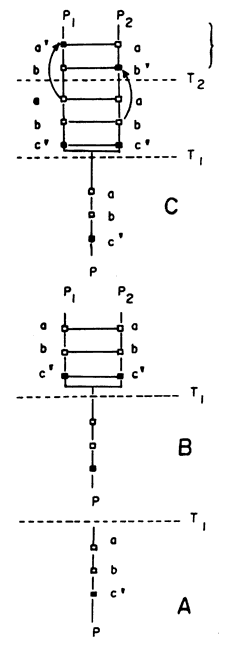
Figura 51. Representación diagramática de un caso simple de especiación,
con diferenciación morfológica.
Hagamos un ejercicio hipotético con el caso de la figura 49. Ya estudiamos
las especies a, b, c, d, e y f, y determinamos sus apomorfías (caracteres derivados).
Transformemos ahora esas especies en conjuntos de apomorfías:
| A= {a', b', c'} |
| B= {a', d', e', f'} |
| C= {a', d', e', g'} |
| D= {a', d', h', i', j'} |
| E= {a', d', h', i', k', l'} |
| F= {a', d', h', i', k', m'} |
Haciendo operaciones con esos conjuntos podemos recuperar la historia evolutiva de las especies correspondientes.
Así, a' es una sinapomorfía de A, B, C, D, E y F. Era, pues, una autapomorfía del antecesor común hipotético de todas esas especies, o sea:
Postulamos, pues, que en el pasado existió un antecesor hipotético, al cual denominaremos M, que poseía una autapomorfía (a') y todos los otros caracteres estaban en estado plesiomórfico, o sea, b, c, d, e, f, g, h, i, j, k, l, m (figura 53).
Como M (el conjunto unitario de a') está propiamente incluido en todas las
especies terminales A, B, C, D, E y F, lo eliminamos de todos esos conjuntos:
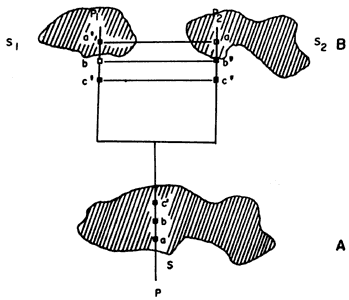
Figura 52.
Y obtenemos así los nuevos conjuntos:
| A1= {b', c'} |
| B1= {d', e', f'} |
| C1= {d', e', g'} |
| D1= {d', h', i', j'} |
| E1 = {d', h', i', k', l'} |
| F1 = {d', h', i', k'. m'} |

Figura 53. El antecesor hipotético M de las especies actuales A, B, C, D,
E Y F.
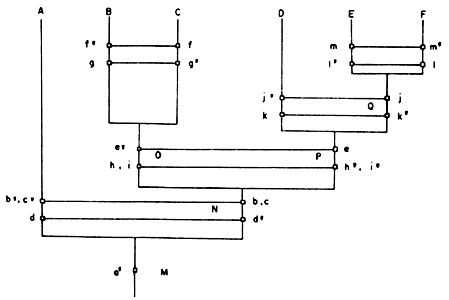
La siguiente operación es hacer intersecciones de esos conjuntos, tomándolos de dos en dos, para ver cómo están relacionados. Es obvio que A1 es diferente de cualquier otro y que B1 Ç C1 Ç D1 Ç E1 Ç F1 = {d'}. Una gráfica de esta se encuentra en la figura 54.
El diagrama de la figura 54 se interpretaría de la siguiente manera: el antecesor hipotético M, que poseía la autapomorfía a', se dividió (especió), originando las especies descendientes A (que llegó hasta el presente) y N /hipotética). A y N son especies hermanas, pues presentan heterobatmia (dada por las autapomorfías b' y c' de A, y d' de N) y son monofiléticas (estrictamente, en este caso), ya que presentan la sinapomorfía a' (de aquí en adelante queda sobreentendido que toda autapomorfía de un antecesor resulta en una sinapomorfía para sus descendientes).
Como el antecesor hipotético N, entendido como el conjunto unitario de d',
está propiamente incluido en B1, C1, D1, E1
y F1, lo eliminamos de esos conjuntos:
| B1 — N = {e', f'} = B2 |
| C1 — N = {e', G'} = C2 |
| D 1 — N = {h', i', j'} = D2 |
| E 1 — N = {h', i', k', l'} = E2 |
| F 1 — N = {h', i', k' m'} = F2 |
Salta a los ojos que B2 Ç C2 = {e'} y que D2 Ç
E2 Ç F2 = {h', i'}. Llamemos a esas intersecciones
no nulas (esto es, con elementos), respectivamente, como O y P (O = {e'}; P
= {h', i'}). La representación gráfica de esto se encuentra en la figura 55.
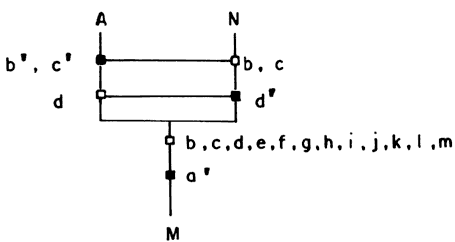
Obtuvimos las especies hermanas (hipotéticas) O y P. Nótese que A es también
especie hermana de O y P (pues presenta heterobatmia con cualquiera de las otras
dos) y es monofilética (pero no estrictamente) con O y P, pues las tres presentan
la sinapomorfía a'.
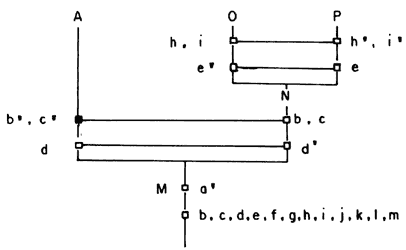
En seguida eliminamos O de B2 y C2. Obtenemos así que B2 y C2 presentan heterobatmia, o sea que son especies hermanas.
Eliminamos P de D2, E2, F2. Vemos que D2 es disyunta de E2 y F2. Estas dos últimas tienen intersección no nula (Q = {k'}) y presentan heterobatmia. Resulta entonces el diagrama de la figura 56, con lo cual termina nuestro análisis filogenético y tenemos así la historia evolutiva de nuestro grupo.
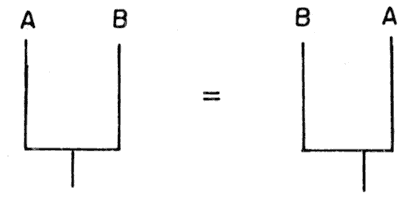
Es la única historia verdadera a la luz de los datos (conjuntos de apomorfías). Pero hay otras historias posibles. ¿Cuál es la probabilidad de que sea verdadera esa reconstrucción de la historia evolutiva de esos taxones? Consideremos lo siguiente: para dos especies terminales tenemos una sola posibilidad (figura 57).
Es equivalente poner A, B como B, A. En ambos casos tenemos que A y B son especies hermanas (estrictamente).
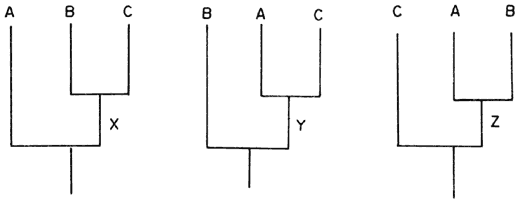
Pero en el caso de tres especies terminales tenemos tres posibilidades de representar su historia evolutiva, si el cladograma incluye secuencias dicotómicas (figura 58).
Las tres historias son distintas pues postulamos diferentes antecesores, según se consideren B y C, A y C, o A y B como especies hermanas estrictas (respectivamente X, Y y Z).
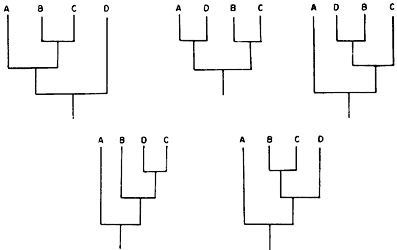
Si adicionamos una cuarta especie terminal, D, y mantenemos secuencias dicotómicas,
obtenemos 15 posibilidades de hacer cladogramas (representaciones gráficas de
las relaciones entre las ramas o clados). Así, para la configuración A (B, C)
podemos poner una cuarta especie D en cinco posiciones distintas.
Como para cada una de las tres configuraciones de la figura 58 la adición de una cuarta especie genera cinco configuraciones (figura 59), obtenemos así 3 x 5 = 15 configuraciones posibles para cladogramas de cuatro especies terminales, con secuencias dicotómicas.
El número de cladogramas posibles aumenta de manera extraordinaria a medida que se añaden especies terminales. La tabla 2 nos da el número de cladogramas posibles para n especies terminales (n varía de 2 a 22); los cladogramas siempre están representados por secuencias dicotómicas.
Por lo tanto, la probabilidad de que nuestro cladograma de la figura 55 sea verdadero es de 1/945.
Ahora podemos reconstruir la historia biogeográfica de nuestro grupo. Hacemos esto remplazando los nombres de las especies terminales del cladograma por sus áreas de distribución. Reconsideremos el mapa representado en al figura 49. Sustituyendo los nombres de las especies terminales por sus áreas de distribución obtenemos el cladograma de áreas de la figura 60.
TABLA 2. Número de cladogramas dicotómicos posibles para n especies terminales.
|
|
|
|
Número de especies terminales
|
Número de cladogramas dicotómicos
posibles
|
|
|
|
|
2
|
1
|
|
3
|
3
|
|
4
|
15
|
|
5
|
105
|
|
6
|
945
|
|
7
|
10 395
|
|
8
|
135 135
|
|
9
|
2 027 025
|
|
10
|
34 459 425
|
|
11
|
654 729 075
|
|
12
|
13 749 310 575
|
|
13
|
316 234 143 225
|
|
14
|
7 905 853 580 625
|
|
15
|
213 458 046 676 875
|
|
16
|
6 190 283 353 629 375
|
|
17
|
191 898 783 962 510 625
|
|
18
|
6 332 659 870 762 850 625
|
|
19
|
221 643 095 476 699 771 875
|
|
20
|
8 200 794 532 637 891 559 375
|
|
21
|
319 830 986 772 877 770 815 625
|
|
22
|
13 113 070 457 587 988 603 440 625
|
|
|
|
También ahora podemos proponer la hipótesis de que la historia del grupo
fue por vicarianza. Como áreas hoy disyuntas, ocupadas por especies hermanas
(en el sentido estricto), constituían sólo un área antecesora; ocupada por el
antecesor único inmediato de las especies que han derivado a partir de ese antecesor
procedemos de la siguiente manera: unimos las áreas ocupadas por dos especies
hermanas en una sola área, que atribuimos al antecesor inmediato de esas especies
hermanas y, bajo ese procedimiento, llegamos hasta el área ocupada por el antecesor
de todo el grupo monofilético. Para nuestro grupo de la figura 49, "volviendo
al pasado" tendríamos la historia que aparece en la figura 61.
Aquí termina el trabajo del biólogo. Pero quedan cuando menos dos problemas por resolver: 1) ¿Cómo podemos demostrar que la historia fue realmente ésa, y que se debió a vicarianza? El patrón actualmente formado por las especies A, B, C, D, E y F podría deberse al puro azar. 2) Si la historia es debida a vicarianza, ¿cuáles fueron los cambios ocurridos en la Tierra que causaron los eventos de especiación biológica y cuáles sus fechas?
Es tarea del biólogo responder a la primera pregunta. El geólogo responde a
la segunda. Veamos cómo. Ilustremos con un ejemplo semihipotético. Consideremos
las aves Ratitae que aparecen en la figura 62. Supongamos, para facilitar
nuestra tarea, que sólo existen estas especies terminales:
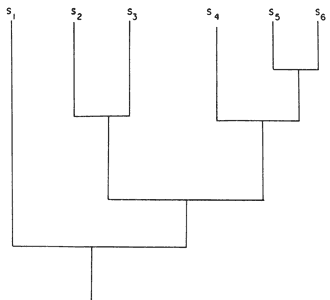
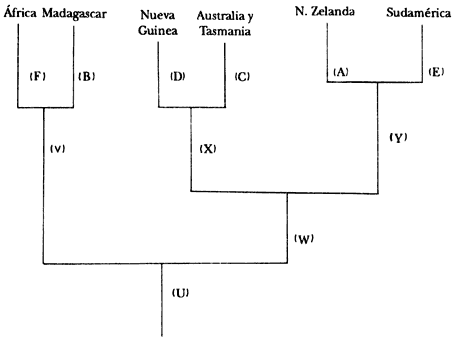
Figura 60. Cladograma de áreas.
B) el Aepyornis (extinto) en Madagascar;
A) el Dinornis (extinto) en Nueva Zelanda;
C) el emú en Australia y Tasmania.
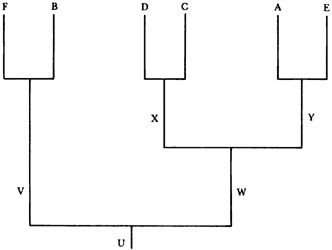
Lo primero que debemos averiguar es la filogenia del grupo, como ya vimos antes. Supongamos que la filogenia resultó como el diagrama de la figura 63. Lo que sigue es transformar el cladograma de especies en un cladograma de áreas. En nuestro ejemplo obtenemos el diagrama de la figura 64.
Entonces proponemos la hipótesis de que Y el antecesor inmediato de A y E,
habitaba un área única, formada por la unión de los bloques actuales llamados
Sudamérica y Nueva Zelanda. Que X, el antecesor inmediato de D y C, habitaba
un área formada por la unión de Australia y Tasmania y Nueva Guinea. Que W,
el antecesor inmediato de X y Y habitaba un supercontinente formado por la unión
de los actuales bloques de Nueva Guinea, Australia, Tasmania, Nueva Zelanda
y Sudamerica. Y finalmente, que U, el antecesor inmediato de V y W, habitaba
un supercontinente formado por la unión de África y Madagascar (en donde vivía
V, el antecesor inmediato de F y B) con el continente en donde vivía W.
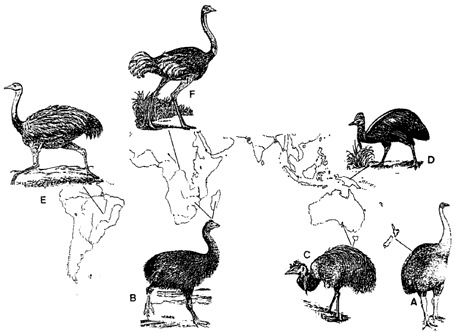
Figura 62. Diferentes especies de Ratitae ysu procedencia geográfica.
¿Cómo poner a prueba esta hipótesis?
Recordemos que, en un mismo lugar de la Tierra, hay muchísimos grupos de animales y de vegetales que están en simpatría. Los grandes eventos de vicarianza deben haber afectado simultáneamente a muchos de esos grupos. Entonces podemos someter a prueba nuestra hipótesis buscando otros grupos monofiléticos que ocupen las mismas áreas actuales que las especies de nuestro grupo y que tengan la misma historia evolutiva. Hay varias posibilidades:
A) Nuestro grupo constituye un patrón único de distribución geográfica;
cartografiando otros grupos distintos, sus respectivas especies nunca estarán
en simpatría con las primeras. En este caso abandonamos la hipótesis de la vicarianza
y adoptamos el azar (dispersión) para explicar el patrón de distribución de
nuestro grupo.
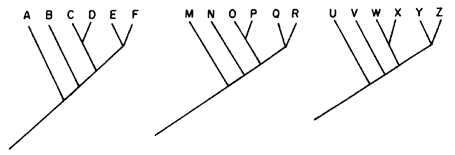
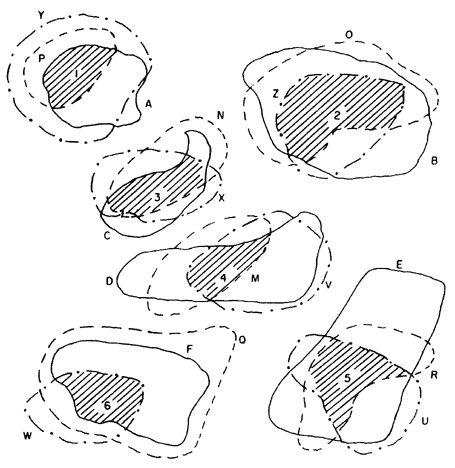
B) Supongamos que existen otros grupos monofiléticos que, al ser trazados en un mapa, muestran que existe simpatría con las especies del grupo que queremos. Por ejemplo, tomamos nuestro grupo A, B, C, E, F que queremos poner a prueba; y otros dos grupos más: M, N, O, P, Q, R, y U, V, W, X, Y, Z. Hacemos su localización en un mapa en forma separada y sobreponemos los tres mapas para comprobar que hay simpatría entre las especies. Obtenemos así un patrón generalizado (figura 65).
Podemos continuar nuestro análisis y hacer la filogenia de los tres grupos. Pueden resultar dos cosas distintas:
a) Los tres cladogramas tienen el mismo dibujo (como el ejemplo de la figura 66). Entonces, el cladograma de áreas sería igual.
b) Los tres cladogramas tienen diferentes dibujos (como, por ejemplo,
en la figura 67)
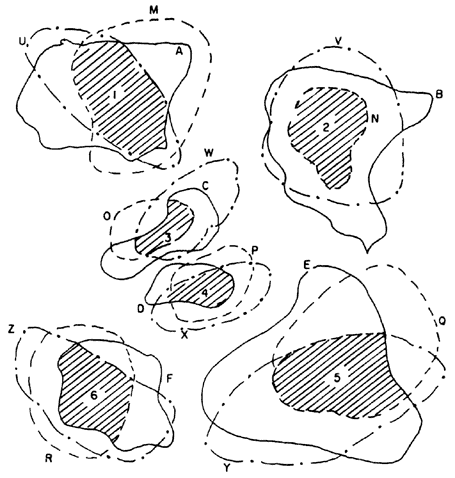
En el caso (a) proseguimos con nuestro análisis (ver C, en el párrafo siguiente). En el caso (b) queda claro que los tres grupos monofiléticos tienen distintas historias evolutivas, y, obviamente, distintas historias biogeográficas. El hecho de que sus especies estén en simpatría se debe, por lo tanto, al azar y no a la vicarianza.
C) En caso de que todos los cladogramas de los grupos comprendidos en
un patrón generalizado de distribución sean idénticos en su forma, continuamos
el análisis transformando los cladogramas de especies en cladogramas de áreas.
Pueden resultar dos cosas:
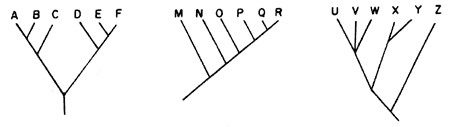
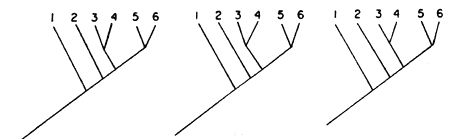
a) Que los cladogramas resulten idénticos (por ejemplo, véase la figura 68; compárela con la figura 66).
b) Que exista un patrón generalizado formado por los distintos grupos monofiléticos, pero que, una vez transformado en cladogramas de áreas, los cladogramas de especies resulten distintos (figuras 69 y 70).
En el caso (b), está claro, los tres grupos tienen historias biogeográficas
totalmente distintas. El patrón generalizado que forman (figura 69) fue debido
al azar.
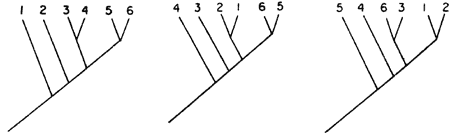
En el caso (a), finalmente, tenemos una probabilidad de que el patrón formado
por los tres grupos se deba a vicarianza. Decimos probabilidad porque nunca
podemos descartar el azar (la dispersión). Por otro lado, podemos calcular cuál
es la probabilidad de que estos cladogramas de especies transformados en cladogramas
de áreas resulten idénticos por azar. Utilicemos la fórmula:
1/x es la probabilidad que tiene cada cladograma de ser verdadero (consúltese
la figura 59); n es el número de cladogramas de especies transformados
en cladogramas de áreas que resultaron idénticos. En nuestro caso hipotético,
pues, cada cladograma tiene la probabilidad de 1/945. Como estamos utilizando
tres cladogramas, la probabilidad de que hayan resultado idénticos los cladogramas
de área única y exclusivamente debido al azar es de:
Podemos aumentar nuestra probabilidad incrementando el número de cladogramas
(si resultan idénticos transformados en cladogramas de áreas). Si utilizamos,
por ejemplo, 10 cladogramas, la probabilidad de que resulten idénticos debido
al azar (y no por vicarianza) se elevará a:
Cuantas más especies terminales haya en nuestros cladogramas, mayores serán
las probabilidades de que su concordancia, una vez transformados en cladogramas
de área, sea debida a vicarianza. Si trabajamos con tres cladogramas de 22 especies
terminales que, una vez transformados en cladogramas de áreas resultan idénticos,
la probabilidad de que el patrón generalizado no sea debido a la vicarianza,
y sí al azar (o sea, a la dispersión), es de exactamente
La debilidad de la hipótesis filogenética se transforma ahora en una fuerza extraordinaria de la hipótesis de vicarianza.
Llega entonces el momento en que el biólogo tiene que pedir a su colega geólogo que le prepare un cladograma geológico del área con la cual trabajó. Si el geólogo puede hacer ese cladograma, es posible que sucedan tres cosas:
1) El cladograma del geólogo es más completo que el del biólogo. Supongamos que para nuestro caso de las aves (figuras 62-64) hayamos obtenido muchos más cladogramas que, reducidos a cladogramas de áreas, resulten como el de la figura 64, y que el geólogo nos da un cladograma tal como el de la figura 44. Como ya dijimos el cladograma geológico es más completo que el biológico, pero este último se "acopla" como un subconjunto propio del primero. Así, podemos interpretar la historia conjunta de la Tierra y la vida y completar la historia de esta última. Nótese que nuestro grupo de aves tenía su antecesor en Gondwana, por lo tanto, podemos decir que es necesario que haya habido una especie hermana (estricta) en Laurasia. El ancestro de ambos grupos estaba en Pangea. Esto apoyaría la hipótesis de que nuestro grupo tiene por lo menos 180 millones de años; que por esa época, al dividirse Pangea en Laurasia y Gondwana, la especie antecesora de los Ratitae especió, con lo cual apareció una especie en Laurasia y otra en Gondwana; la primera aún no ha sido encontrada como fósil. El antecesor que ocupaba Gondwana también especió cuando ese supercontinente se dividió en Afroindia y el resto, hace unos 80 millones de años. Después, Madagascar se separó de África (ese hecho no está representado en el diagrama de la figura 44), y la especie que ocupaba todo ese continente se separó, originando (en nuestro caso semihipotético) el avestruz de África y el Aepyornis de Madagascar, que son (en nuestro ejemplo) especies terminales (el Aepyornis hoy está extinto). Pero vemos en el diagrama de la figura 44 que India estaba unida a África en ese tiempo, por lo cual debemos esperar encontrar también en la India un representante fósil de ese grupo de aves. El resto de Gondwana era entonces ocupado (hace 80 millones de años) sólo por una especie, que se fragmentó posteriormente con la división de ese supercontinente, generando una especie en Australia (que según nuestra hipótesis sería el actual emú, y una especie hermana que ocupaba, hace 70 millones de años, el bloque constituido por Nueva Zelanda, Antártida occidental y Sudamérica. Con la partición de ese bloque en dos la especie se separó y surgió el casuar de Nueva Zelanda y su especie hermana, que debería estar en Antártida occidental y Sudamérica. Finalmente, esta especie dio origen al ñandú en Sudamérica y su especie hermana (desconocida) en Antártida occidental (si es que no estaba congelada por ese tiempo). Esa última especiación ocurrió hace 40 millones de años. Claro que todo esto es hipotético, pero necesario a la luz de la teoría de la vicarianza. Así, gracias al cladograma geológico podemos completar "la historia de lo vivo" y, también, "predecir" la existencia de especies fósiles (con todos sus caracteres apomórficos) antecesoras en todas las uniones de bloques continentales pasados. Gracias a la teoría de la vicarianza tenemos ahora, con muchos detalles, la historia de la coevolución de la Tierra y la vida, las fechas de los acontecimientos de especiación (de la Tierra y la vida), podemos saber cuáles fueron las barreras físicas que los causaron, cuántas especies antecesoras han existido, cuáles fueron sus caracteres, dónde vivían y durante cuál periodo geológico. Y podemos igualmente conocer el número mínimo de especies terminales que, o se extinguieron y aún no han sido encontradas como fósiles, o que viven pero no han sido recolectadas u observadas.
2) El cladograma del geólogo es idéntico al cladograma de áreas de los biólogos. En este caso sabemos que no hubo extinciones en los grupos de organismos y podemos poner fechas a todos los eventos de especiación y saber cuáles han sido las barreras causantes de la especiación.
3) El cladograma de áreas obtenido por los biólogos tiene una altísima probabilidad de ser verdadero (superposición de muchos cladogramas de áreas idénticos), pero está en total desacuerdo con el cladograma del geólogo. En este caso, el geólogo es quien está equivocado y tiene que rehacer su cladograma pues, estadísticamente, si distintos grupos, con distintas ecologías y biologías, tuvieron la misma historia biogeográfica (tal como lo revela el cladograma de áreas), la probabilidad de que esa identidad sea debida al azar (dispersión) es astronómicamente baja. Es el geólogo quien tiene que reexaminar sus datos pues trabaja con un solo sistema, la Tierra. Los biólogos podemos trabajar con muchísimos sistemas que están en simpatría.
La vicarianza es la teoría central de la biología, una nueva ciencia que engloba todas sus ramas, desde la morfología hasta la ecología. Es una síntesis revolucionaria, la mayor revolución surgida en la milenaria historia de la biología. Sobre ella se apoya el análisis de las relaciones históricas de sus distintas entidades.
La elegancia suprema de la teoría de la vicarianza es que la historia de la Tierra se encuentra escrita en los cromosomas de los seres vivos.
![[Inicio]](../img/begin.gif)
![[Anterior]](../img/prevsec.gif)

![[Siguiente]](../img/nextsec.gif)