VIII. EJEMPLOS DE LOS MECANISMOS DE ACCIÓN DE ALGUNAS HORMONAS Y NEUROTRANSMISORES
ESTE CAPÍTULO está destinado a analizar con mayor detalle las acciones (qué hacen) y los mecanismos de acción (cómo lo hacen) de algunos de los mensajeros, en mi opinión, más interesantes. Por supuesto, empieza por la adrenalina, mi hormona favorita, también llamada epinefrina.
La adrenalina fue descubierta en 1895 por Oliver y Schäfer en extractos de glándula suprarrenal. Estos extractos tienen la propiedad de aumentar la tensión arterial y la frecuencia cardiaca. Posteriormente Stolz y Dakin identificaron su estructura química y finalmente fue sintetizada. La estructura de esta hormona se muestra en la figura 16. Se forma a partir del aminoácido tirosina principalmente en la médula de la glándula suprarrenal y en algunas neuronas, las llamadas neuronas simpáticas. Funciona, por lo tanto, como hormona y como neurotransmisor. La noradrenalina o norepinefrina es un precursor en la biosíntesis de la adrenalina. En realidad ambos compuestos se encuentran tanto en las neuronas como en las suprarrenales. Sin embargo, en general se acepta que la adrenalina es la hormona y la noradrenalina el neurotransmisor.
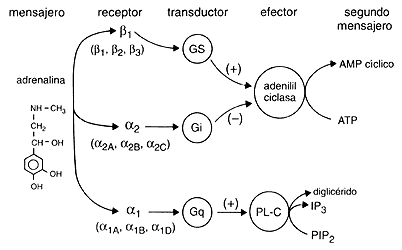
FIGURA 16. Estructura de adrenalina y representación esquemática de
su mecanismo de acción.
La adrenalina ejerce importantes funciones en todo el cuerpo. Puede asegurarse con facilidad que no hay una función de grande o mediana importancia para el organismo en la que no participe. Por lo tanto, no es de sorprender que una enorme proporción de las células de nuestro cuerpo tengan receptores adrenérgicos. Todo ello es ventajoso para quienes nos dedicamos a este campo; por lo mismo, la competencia es enorme. Es un campo apasionante, prácticamente en constante ebullición.
Para mencionar algunas de las principales funciones de los agentes adrenérgicos diré que, dado que son de los neurotransmisores más abundantes, participan en un gran número de las llamadas funciones superiores, además de que regulan la frecuencia cardiaca, la tensión arterial, la secreción por glándulas tanto de secreción interna como externa, el metabolismo global de la economía al regular los metabolismos específicos de órganos como el hígado, el tejido adiposo y el músculo, en fin, en casi todo tienen que ver. Sería interminable una lista de las células sobre las que puede actuar la adrenalina; entre ellas tenemos a la inteligente neurona, al infatigable miocito cardiaco, al humilde adipocito, a los versátiles hepatocitos, a las pequeñas plaquetas, etcétera.
Como resultará obvio de lo anteriormente mencionado, existe una gran cantidad de enfermedades en las que de una forma u otra están implícitas las funciones de los agonistas adrenérgicos naturales, la adrenalina y la noradrenalina. Así, se ha sugerido que en algunos trastornos mentales, como son los estados maniacos y los depresivos, existe una alteración en el metabolismo y función de estas catecolaminas. En la hipertensión arterial tienen una función importante y de hecho algunos de los fármacos que se usan para su tratamiento son agentes con propiedades adrenérgicas. Se ha propuesto que los receptores adrenérgicos son un factor clave en el asma, y para el tratamiento de los cuadros asmáticos agudos se usan agonistas adrenérgicos. Muchos de los descongestionantes nasales contienen agentes adrenérgicos.
Es fácil, entonces, imaginar por qué es importante saber qué hacen y sobre todo cómo hacen lo que hacen, los agentes adrenérgicos. Para ello hay razones científicas básicas: el deseo de saber más y profundizar en los fenómenos esenciales de la vida; y razones aplicativas: lo que se descubra es importante para muchas enfermedades (desde el resfriado común hasta las enfermedades cardiovasculares, pasando por los problemas mentales).
Existen además razones económicas: la industria farmacéutica investiga (desafortunadamente no en nuestro país) muy diversos agentes adrenérgicos para su posible utilización. Por ejemplo, la venta de un agente antiadrenérgico de uso clínico representó 50% de los ingresos de una enorme compañía trasnacional durante la década de los setenta. El investigador que logró el compuesto, sir James Black, contribuyó además con otro compuesto (bloqueador de la acción de la histamina) de uso clínico importantísimo y con grandes beneficios económicos para la industria en la que trabajaba. Hace algunos años, recibió el premio Nobel de Fisiología y Medicina por su trabajo pionero en farmacología molecular.
Pero dirijámonos a lo más interesante. La adrenalina inicia sus acciones al asociarse con receptores membranales. En 1948, un investigador mexicano, Arturo Rosenblueth (quien trabajaba con el doctor Canon en el noreste de Estados Unidos), observó que la adrenalina producía contracción en un músculo liso: la aorta; por el contrario, el músculo liso de los bronquios se relajaba. Perplejos por estos resultados contradictorios, dichos investigadores propusieron que se generaban dos tipos de mediadores, según el tejido en el que actuaba la hormona, y que conducían a los diferentes efectos. Esta sugerencia no fue acertada, pero sí constituyó un inicio conceptual para dividir las acciones de la hormona. Otro investigador, Alhquist, también en 1948, llegó a una conclusión más válida y que de hecho, constituye la piedra angular de lo que hoy sabemos sobre acciones adrenérgicas. Este investigador recientemente fallecido, al usar una serie de análogos de la adrenalina, observó que la potencia relativa de estos compuestos para producir contracción o relajación era claramente diferente. Estas observaciones y, por supuesto, su capacidad, lo llevaron a la conclusión de que la contracción ocurría por la activación de un tipo de receptor al cual llamó a; mientras que la relajación se daba por la activación de otro tipo de receptor al que denominó b. Hoy sabemos que estos tipos de receptores se subdividen aún más, y que en realidad hay tres familias de receptores para la adrenalina: los a1-, los a2- y los b-adrenérgicos. Cada una de estas familias tiene tres miembros y por lo tanto tenemos nueve receptores: a1 (A, B y D), a2 A, B y C) y b (1, 2 y 3).
Quizá parezca más un entretenimiento que otra cosa eso de andar subdividiendo receptores para una hormona dada, pero tiene una importancia teórica y práctica increíble. Pondré un ejemplo: durante muchos años se ha sabido que las catecolaminas aumentan las necesidades de oxígeno del corazón al activar a los receptores b-adrenérgicos del miocardio. Existen condiciones en las cuales es conveniente bloquear en un paciente este tipo de receptores. Para ello se han diseñado diversos bloqueadores b-adrenérgicos. Por otro lado, en los cuadros asmáticos se utilizan agonistas b-adrenérgicos para relajar la musculatura bronquial. Ahora bien, imaginemos a un paciente que tenga ambos tipos de problemas: cardiovasculares y asmáticos. Si el galeno prescribe un bloqueador b-adrenérgico general, el paciente mejorará de su padecimiento cardiovascular, pero, ¿qué sucederá si presentara un cuadro de asma? Los receptores b de sus bronquios estarían también bloqueados. Afortunadamente, hoy en día la situación no es tan complicada. Me explicaré mejor: el receptor b del miocardio es b1-adrenérgico, mientras que el de los bronquios es b2 Dado que la industria farmacéutica ya ha logrado sintetizar agentes selectivos para los diferentes subtipos, podemos administrar al paciente un bloqueador b1-adrenérgico selectivo y dejar en paz sus b2 para cualquier emergencia asmática.
Decía entonces que hay tres familias o tipos básicos de receptores adrenérgicos, todos ellos pertenecen a la familia de los siete dominios transmembranales o acoplados a proteínas G. En general estos receptores se encuentran distribuidos en forma preferencial en los diferentes tejidos; por ejemplo: el corazón es rico en receptores b1-adrenérgicos, mientras que la aorta contiene múltiples receptores a1-adrenérgicos. El conocimiento de que existen tantos receptores diferentes para esta hormona y neurotransmisor es muy reciente, y aún se ignora la distribución por tipo y subtipo en cada tejido y su participación en las acciones de este mensajero. Por otro lado, una célula puede contener más de un tipo de receptores para la adrenalina. No hace muchos años, en 1981, demostramos que el adipocito humano contiene receptores adrenérgicos de las tres familias principales, es decir al, a2 y b-adrenérgicos. De hecho, de la familia b, el adipocito expresa los tres subtipos: el b1, el b2 y el b3. Ahora sabemos que este hecho es bastante general y que frecuentemente las células expresan vanos de los subtipos para una misma hormona.
Otro aspecto sumamente interesante es que los receptores están asociados a sistemas específicos de transducción. Mi maestro, John Fain, y yo fuimos los primeros en establecer claramente este hecho. Como hemos visto, los diferentes receptores b-adrenérgicos están acoplados activadoramente a la adenilil ciclasa a través de Gs como hemos visto. Los receptores a2-adrenérgicos se acoplan a la ciclasa en forma inhibitoria a través de Gi, y los receptores a1 se acoplan al recambio de fosfoinosítidos a través de Gq (ver la figura 16). Además de la existencia de los diferentes subtipos, en los últimos años nos ha quedado claro que cada receptor no enciende un señalamiento lineal, sino una red, como discutimos cuando hablamos de las proteínas G. Así, es claro que estos receptores, además de modular la adenilil ciclasa y el recambio de fosfoinosítidos, tienen acciones sobre otros sistemas de transducción, como son algunos canales para sodio, calcio y potasio. Más aún, el acoplamiento de un receptor no sólo depende del receptor sino del repertorio de proteínas G y de efectores (canales iónicos, otras fosfolipasas, etc.) que expresa cada célula.
Una hormona que ha atraído el interés de miles de científicos y del público en general es la insulina. Esta hormona es un polipéptido con un peso molecular de 6 000 Daltones, y es producida y secretada por las células beta de los islotes de Langerhans del páncreas. Sin duda, gran parte del interés que se ha generado, se debe a su importancia en el mantenimiento de los niveles de glucosa en la sangre y en el tratamiento de la diabetes mellitus.
Alrededor de 1890 Mering y Minkowsky habían demostrado que la extirpación del páncreas produce, en animales de laboratorio, un padecimiento similar a la diabetes mellitus. Fueron unos investigadores canadienses del Hospital General de Toronto, Banting y su alumno Best, un joven estudiante de medicina, quienes lograron extraer el principio activo del páncreas y demostraron su utilidad terapéutica tanto en perros diabéticos como en humanos; estos estudios se realizaron entre 1921 y 1922. En esos trabajos participó McLeod, en ese momento jefe del Departamento, y con quien Banting tuvo innumerables conflictos. Recibió años después junto con Banting el premio Nobel de Fisiología y Medicina; Banting, por su cuenta decidió compartir su parte del premio con Best.
El primer paciente en recibir el beneficio de tales descubrimientos fue Leonard Thompson, un muchacho diabético de 14 años internado en el Hospital General de Toronto. La diabetes de este muchacho se encontraba totalmente descompensada y Banting y Best decidieron emplear su extracto como un tratamiento experimental. El resultado fue espectacular y abrió, sin duda, una de las grandes avenidas en la historia del tratamiento de las enfermedades endocrinas en general y de la diabetes en particular. Para 1926 ya se contaba con insulina cristalina, y en 1960 Langer estableció su secuencia de aminoácidos. Este investigador obtuvo el premio Nobel de Química.
Los estudios acerca de la insulina han continuado avanzando. Actualmente podemos cuantificar con precisión la cantidad de insulina que hay en la sangre y en los diversos tejidos (metodología que está asociada a otro premio Nobel); además existen muy diversos preparados de insulina con diferentes velocidades de absorción para un mejor tratamiento de los pacientes. Dos avances notables merecen también ser mencionados; dado que el consumo de insulina (obtenida principalmenté del cerdo) es muy importante, se ha considerado que su disponibilidad podía llegar a ser limitante para el tratamiento del gran número de pacientes que la requieren. Para evitar esto, en la década de los sesenta se desarrollaron los procesos de síntesis química completa. Estos procesos, aunque costosos, ofrecen una alternativa y se han mejorado en los últimos años. Otra opción que ya ha resultado de utilidad práctica es la inserción de la información genética para la síntesis de la hormona en el ADN de microorganismos. Estos microorganismos producen la hormona y es posible cultivarlos en grandes fermentadores industriales. Esta alta tecnología de ingeniería genética se emplea con éxito en los laboratorios Lilly de Estados Unidos, que producen desde hace varios años insulina por estos medios, para uso en pacientes.
La insulina es la principal hormona encargada de disminuir los niveles de glucosa en sangre. De los años treinta a los setenta se descubrieron muchas de las principales acciones de la insulina. Esta hormona aumenta el transporte de glucosa al interior de las células y su conversión a glucógeno; además aumenta la oxidación del azúcar. Favorece el proceso de síntesis de lípidos y disminuye tanto la movilización de grasa de los depósitos, como su oxidación en el hígado; además, aumenta el transporte de algunos aminoácidos en las células blanco. Estas acciones, y muchas otras descritas hasta nuestros días, ocurren rápidamente, en cuestión de minutos. Además, hay clara evidencia de que la insulina ejerce acciones más tardías, las cuales toman de horas a días; dentro de estas últimas está el ser un factor de crecimiento celular.
Pero, ¿cómo hace lo que hace la insulina? Sin duda uno de los grandes avances que han ocurrido en los últimos 15 años es el conocimiento de la estructura del receptor de la insulina. El trabajo pionero de Pedro Cuatrecasas aclaró varios aspectos. Este investigador acopló la insulina a moléculas de gran tamaño que no podían entrar a las células; la insulina, así acoplada, aun podía ejercer sus efectos. Ello establecía que el receptor se localizaba en la membrana plasmática. Por otro lado, intentó purificar el receptor, con relativo éxito. Las técnicas que se usaban en ese momento no le permitían lograr la absoluta reproducibilidad de los resultados. Su trabajo fue sujeto de múltiples críticas, justas e injustas. Hoy, más de 20 años después, muchos de sus hallazgos han sido totalmente comprobados y su labor pionera es reconocida por los expertos.
Hacia 1980 trabajé en Brown University con mi maestro John Fain. En los laboratorios que estaban al final del corredor trabajaba Mike Czech, un joven y brillante ex alumno de Fain que lideraba a un grupo de entusiastas y capaces posdoctorales. Paul Pilch, químico de corazón, había logrado entrecruzar la insulina radiactiva con su receptor y, por medio de técnicas refinadas (electroforesis y radioautografía), había llegado a establecer el peso molecular del receptor. Poco después se incorporó al grupo Joan Massagué, joven catalán quien avanzó audazmente en el estudio de las subunidades del receptor. Así, en unos cuantos años este grupo logró establecer la estructura del receptor de la insulina. Un sueño que durante muchos años habían acariciado muchos investigadores. Vale la pena mencionar que nadie en este mundo está aislado, y así otros grupos, incluyendo al de Cuatrecasas, hicieron importantes contribuciones para afinar el modelo actual que se presentó en la figura 10. El receptor de la insulina es una proteína de peso molecular aproximado de 310 000 Daltones, está formada por dos subunidades llamadas alfa con peso de 125 000 y dos beta con peso aproximado de 90 000 Daltones. Estas subunidades están enlazadas por uniones disulfuro. Parece existir solamente un gen para el receptor de la insulina, pero por procesamiento alternativo del ARN que lo codifica da origen a dos subtipos de receptores, A y B, para la hormona. Hay evidencia de que el receptor es sintetizado como una sola proteína y posteriormente es dividido y procesado. El procesamiento de este precursor del receptor no sólo involucra el fraccionamiento en sus subunidades, sino que además participan otros procesos como la incorporación de azúcares; dicho procesamiento posiblemente ocurra en vesículas especializadas del aparato de Golgi.
Las subunidades alfa contienen el sitio de fijación de la insulina. Hay evidencia de que podría existir más de un sitio para la hormona y de que quizá haya cierta interacción de un sitio con el otro. Las subunidades beta han sido motivo de interesantísimos hallazgos. Kasuga y Kahn demostraron que el receptor tiene actividad de proteína cinasa de tirosina. Posteriormente el gen que codifica el receptor de la insulina fue donado y ha sido expresado en muchos sistemas, así como sujeto a diferentes manipulaciones (mutaciones, formación de quimeras, etc.) para avanzar en el mecanismo de su acción. El receptor se fosforila tanto por su propia actividad de tirosina cinasa (autofosforilación) como por otras cinasas (fosforilación heteróloga); el mecanismo general de acción de estos receptores con actividad de proteína cinasa ha sido descrito en el capítulo V. Indudablemente, mucho se ha avanzado; sin embargo, faltan muchos aspectos por aclarar.
Una las principales acciones de la insulina es disminuir la concentración de glucosa en la sangre, lo cual se logra al aumentar el transporte de azúcar al interior de las células; este efecto puede observarse fácilmente en células aisladas. Durante muchos años se ha debatido acerca de los mecanismos moleculares responsables de dicho efecto. Sabemos que el transporte de la glucosa se lleva a cabo por medio de un transportador específico, el cual se ha aislado, e incluso reconstituido, en membranas artificiales. La pregunta fundamental ha sido, ¿cómo aumenta la insulina el transporte del azúcar? Se buscaron posibles activadores sin encontrarse ninguno. No hace mucho tiempo, dos grupos, el de Cushman y el de Kono, encontraron un hecho que ha cambiado nuestra concepción del fenómeno. En pocas palabras, se descubrió que el número de transportadores en la membrana plasmática aumenta considerablemente bajo la acción de la insulina. La pregunta inmediata fue, ¿de dónde vienen? Y surgió que, al igual que con los receptores, los transportadores se localizan no sólo en la membrana plasmática, sino también en vesículas intracelulares. Más experimentos confirmaron lo anterior y además aportaron la siguiente observación: bajo la acción de la insulina los transportadores intracelulares de glucosa se incorporan a la membrana plasmática; por lo tanto, el número de transportadores disminuye en las vesículas intracelulares y aumenta en la membrana plasmática. Además, al terminar la acción de la insulina el proceso se revierte. Pero, ¿cómo se relacionan la actividad de tirosina cinasa del receptor y la translocación de transportadores de glucosa? Ésta es una de las grandes lagunas que quedan aún en nuestro conocimiento.
C) HISTAMINA, SEROTONINA, PROSTAGLANDINAS Y ADENOSINA
Existe un amplio grupo de sustancias de intensa actividad biológica y enorme importancia que funcionan como hormonas locales. Muchas de ellas son producidas por el organismo en respuesta a cambios o agresiones localizadas; su función, desde un punto de vista teleológico, es decir finalista, es restablecer el equilibrio perdido; en este sentido son un remedio endógeno, esto es, del mismo organismo, para sus males. En 1916, sir Edward Schäfer acuñó el término "autacoide" para referirse a las hormonas en general. Sin embargo, la palabra autacoide describe en forma corta y con mucha mayor propiedad a las hormonas locales; es por ello que se utiliza principalmente en este último sentido. Como mencioné ya en un capítulo anterior, desde el punto de vista etimológico este término proviene del griego autos que significa propio, y de akos que significa remedio o medicamento. Es decir, el significado etimológico se adapta claramente a la idea de la hormona local, la cual es un automedicamento para restablecer el equilibrio del organismo. Dentro de este amplio grupo de sustancias he seleccionado a cuatro de ellas, relativamente conocidas como hormonas locales y con gran importancia fisiológica. Ellas son: la histamina, la serotonina, la adenosina y las prostaglandinas.
Este autacoide, cuya fórmula presento en la figura 17, es el producto de la descarboxilación (eliminación de C02) de un aminoácido: la histidina. Es muy interesante recordar que este producto fue aislado y sintetizado como una curiosidad química mucho antes de conocer su función en el organismo. Existía la creencia que era sólo un producto de la acción de las bacterias. Para 1910 ya se sabía que se producía en muchos tejidos, principalmente en el músculo liso de los vasos sanguíneos; sin embargo, no fue sino hasta casi 20 años después, en 1927, que Best, Dale, Dualery y Thorpe la aislaron de tejidos frescos. Este hallazgo estableció, sin dudas, que la histamina es un constituyente natural de los tejidos. Es interesante además hacer notar que la palabra histamina proviene del griego histos que quiere decir tejido y de amina: la amina de los tejidos.
La inmensa mayoría de quienes no se dedican a las ciencias biomédicas, cuando oyen hablar de antihistamínicos (bloqueadores o antagonistas de la acción de la histamina), piensan exclusivamente en reacciones de tipo alérgico; lo cual no es sorprendente, ya que durante muchos años casi toda la investigación y por ende el conocimiento estuvieron enfocados a este aspecto. Los antihistamínicos clásicos, que todos conocemos, bloquean sólo un tipo de los receptores para la histamina, los H1.
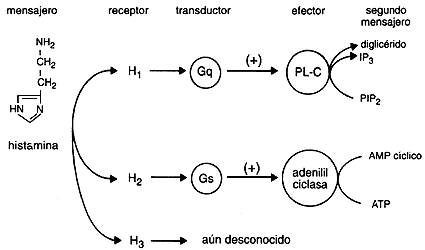
FIGURA 17. Estructura de la histamina y representación esquemática de su mecanismo de acción.
Ya en los años cincuenta había evidencia de que existía más de un tipo de receptor para este autacoide, pero no fue sino hasta los setenta cuando se sintetizaron compuestos con selectividad para el otro tipo de receptores, para la histamina, los H2. En estos desarrollos fue fundamental el trabajo de sir James Black, a quien nos referimos en líneas anteriores. Poco después de la síntesis de los antihistamínicos selectivos H2, éstos fueron incorporados al arsenal de medicamentos usados en la práctica clínica. Desde hace unos dos o tres años se han publicado trabajos que sugieren la existencia de un tercer tipo de receptor para la histamina: el H3.
Represento en la figura 17 lo que sabemos de estos receptores. Como puede apreciarse, el receptor H1 —el asociado a los fenómenos alérgicos— está acoplado al recambio de fosfoinosítidos y al calcio, mientras que el receptor H2 está acoplado en forma activadora a la adenilil ciclasa a través de Gs. En el caso del receptor H3 aún se desconoce el sistema transductor al que se acopla. Mencionaré algunas de las principales acciones de la histamina y el tipo de receptor involucrado. Se ha observado que la histamina produce dos tipos de efectos sobre los vasos sanguíneos: 1) dilatación, mediada a través de receptores H2, y 2) contracción, mediada por receptores H1. El enrojecimiento y el edema localizado, observados en las reacciones alérgicas se producen al activarse los receptores H1. Otro fenómeno alérgico que involucra a receptores histamínicos de este tipo es la bronco-constricción, asociada a cuadros de tipo asmático. El corazón tampoco escapa de tener receptores para la histamina; en este órgano, la activación de receptores H1 disminuye la conducción eléctrica, y en algunas especies hay receptores H2 que provocan taquicardia. La histamina también modula la presión arterial: aparentemente la activación de los receptores H1 la eleva y la de los H2 la disminuye. La histamina es un importante neurotransmisor en el sistema nervioso central; los tres subtipos conocidos de receptores para la histamina, parecen tener muy diversas funciones en el cerebro.
Otro efecto importante de la histamina es aumentar la secreción gástrica. Durante muchos años se supo que la histamina aumentaba la secreción de ácido clorhídrico en el estómago. Sin embargo, el hecho de que ninguno de los antihistamínicos conocidos hasta entonces (de tipo H1) mostrara utilidad para reducir la secreción gástrica hacía dudar de la importancia fisiológica de dicho fenómeno y se consideraba como una curiosidad con cierta utilidad diagnóstica. Con la síntesis de antihistamínicos H2 resultó claro que la histamina sí juega un papel fisiológico en la regulación de la secreción gástrica. Además, estos antagonistas H2 han resultado de enorme utilidad para el tratamiento de los pacientes con úlcera gastroduodenal.
Durante muchos años se supo que si se dejaba coagular la sangre, en el suero se encontraba una sustancia vasoconstrictora, es decir, que aumenta el tono vascular. En 1948 Rapport y sus colaboradores aislaron y cristalizaron este compuesto, al cual le dieron el nombre de serotonina (unión de las palabras suero y tono muscular). Este compuesto, sintetizado a partir del aminoácido triptofano (la serotonina es la 5-hidroxi-triptamina, 5-HT), se encuentra localizado básicamente en tres lugares: en las células cromafines del intestino, en algunas neuronas del sistema nervioso central y en las plaquetas. Han sido identificados muy diversos tipos de receptores para este compuesto; por lo menos siete familias, y algunas con varios subtipos. Aparentemente los diferentes subtipos de la familia 5-HT1 están acoplados en forma inhibitoria a la enzima adenilil ciclasa y a diversos canales iónicos, a través de Gi; mientras que los 5-HT2, están acoplados al recambio de fosfoinosítidos y al calcio. Los receptores 5-HT3 son receptores canal, los 5-HT4, 5-HT6 y 5-HT7 se acoplan activadoramente a la adenilil ciclasa, y de los 5-HT5 aún se desconoce su mecanismo de acoplamiento (figura 18). Como podrá observarse este es un grupo de enorme complejidad con muy diversos tipos de receptores. En este momento la tipificación de los receptores que participan en cada una de las acciones de la serotonina es un campo de gran actividad.
Como es de esperarse, con base en su distribución, la serotonina afecta el funcionamiento intestinal. Este autacoide aumenta notablemente la motilidad del intestino delgado. Por otro lado, produce vasoconstricción y aumenta tanto la fuerza como la frecuencia del latido cardiaco. En la hipófisis la serotonina parece ser un importante regulador de la secreción de algunas hormonas, como la del crecimiento, la prolactina y las gonadotróficas. Hay clara evidencia de que la serotonina es un neurotransmisor en el sistema nervioso central; se ha postulado que alteraciones en el metabolismo o en la acción de este compuesto pueden tener relevancia en algunos desequilibrios mentales como la esquizofrenia o el autismo infantil. Otro dato de importancia es que algunos compuestos alucinógenos (como la psilocina y la psilocibina, productos de los hongos alucinógenos y el LSD) interactúan fuertemente con receptores serotonínicos. Además este tipo de receptores parece tener una gran importancia en la modulación del estado de ánimo y por lo tanto asociado a trastornos depresivos. Ésta es un área de intensa actividad.

Figura 18. Estructura de la serotonina (5-hidroxi-triptamina) y representación
esquemática de su mecanismo de acción.
En 1930 dos ginecólogos estadounidenses, Kurzrok y Lieb, descubrieron que el útero se relajaba y contraía al exponerse el semen humano. Pocos años más tarde, Goldblat en Inglaterra y Wuler en Suecia reportaron que el extracto de la próstata tenía actividad sobre el músculo liso. El material fue identificado por Euler como un lípido, quien lo bautizó con el nombre de "prostaglandina". Fue necesario un gran avance técnico para aislar e identificar estos compuestos, lo cual no se logró sino hasta 1960. De esa época a la fecha el número de publicaciones que hablan de estos compuestos y que describen sus acciones ha aumentado en forma sorprendente. Ahora sabemos que el ácido araquidónico (un ácido graso polinsaturado, presente en muchos fosfolípidos de la membrana) es el precursor de un grupo de compuestos muy activos biológicamente, entre los cuales se encuentran las prostaglandinas, los leucotrienos y los tromboxanos.
Me centraré en las prostaglandinas, no sin antes mencionar que una de las drogas más ampliamente usadas en el mundo, la aspirina, es un inhibidor de la síntesis de prostaglandinas. Como ustedes saben, la aspirina se utiliza como analgésico (quita el dolor), antipirético (suprime la fiebre) y antiinflamatorio. Muchas de estas acciones de la aspirina se deben a que bloquea la formación de prostaglandinas en el organismo. En este momento hay evidencia de diversos tipos de receptores para estos compuestos con acoplamientos a la adenilil ciclasa y al recambio de fosfoinosítidos. Todos estos receptores parecen corresponder a la familia de los siete dominios transmembranales.
Algunos de los efectos más prominentes de las prostaglandinas son los siguientes: 1) inhiben la agregación de las plaquetas al oponerse al efecto de los tromboxanos (la aspirina bloquea la síntesis de prostaglandinas pero también la de tromboxanos, y así ejerce su acción antitrombótica); 2) el músculo liso presenta varios tipos de respuestas: a) el músculo bronquial se relaja bajo la acción de las prostaglandinas, por lo que se han usado en el tratamiento de pacientes asmáticos, b) las prostaglandinas aumentan la contracción del útero —por lo mismo, se han tratado de usar como abortivos—, y c) el músculo liso gastrointestinal varía en su respuesta según la región anatómica; 3) otra acción importante es la disminución de la secreción gástrica. Esta acción ha permitido tener la esperanza de diseñar análogos que puedan ser útiles en el tratamiento de la úlcera péptica; 4) en el sistema nervioso las prostaglandinas aumentan o disminuyen la actividad eléctrica, según la región; 5) además, sabemos que son importantes en la regulación de la secreción de algunas hormonas, y 6) desde el punto de vista metabólico su acción más importante es disminuir la movilización de grasa de los depósitos.
Este compuesto es un producto del metabolismo del ATP, la moneda
energética de la célula; se libera en muchos tejidos cuando las células tienen
una deficiencia en su aporte de oxígeno. Así, se ha visto que cuando hay falta
de oxigenación en el corazón se libera adenosina, la cual tiende a compensar
el problema aumentando el riego coronario y disminuyendo el gasto energético.
Al igual que en el caso de las prostaglandinas, este compuesto que inhibe la agregación plaquetaria, dilata los vasos sanguíneos principalmente de las circulaciones coronaria (cardiaca) y cerebral, disminuye la liberación de algunos neurotransmisores de tipo excitador en el sistema nervioso central, disminuye la contracción del músculo intestinal, disminuye la lipólisis en el tejido adiposo, e incluso actúa como protector de la función hepática. Esta diversidad de acciones nuevamente nos ilustra la importancia fisiológica que posiblemente tenga el compuesto, pero también nos habla de las dificultades para cualquier aplicación terapéutica. Sin embargo, se han diseñado compuestos que alteran el transporte o el metabolismo de la adenosina, los cuales sí se han usado ya con fines terapéuticos. Ésta es también un área activa y lo va a ser más en los próximos años.
Existen diversos receptores para la adenosina que junto con los del ATP constituyen la familia de los receptores purinérgicos. Todos ellos parecen también formar parte de la gran familia de los receptores con siete dominios transmembranales.
Otro aspecto de interés, respecto a la adenosina, es su relación con las metilxantinas. Estas últimas son compuestos bloqueadores de la fosfodiesterasa, la cual rompe e inactiva el AMP cíclico, y además son potentes antagonistas de las acciones de la adenosina. Las metilxantinas están consideradas entre las drogas de mayor consumo por los seres humanos en nuestros días. La cafeína, la teofilina y la teobromina son algunos de los integrantes principales de este grupo, y son constituyentes activos del café, el té y el chocolate. Cierto es que, cada vez con mayor frecuencia, se ingiere café descafeinado, pero para compensar se consume una gran cantidad de cafeína en los refrescos de cola. Es muy posible que el efecto estimulante de estas bebidas se deba al antagonismo de la acción de la adenosina que ejercen las metilxantinas.
D) GLUCAGON, VASOPRESINA Y ANGIOTENSINA II
En esta sección trataré de revisar brevemente algo de lo que sabemos acerca de tres hormonas de naturaleza peptídica, las cuales ejercen una profunda acción sobre el metabolismo: el glucagon, la vasopresina y la angiotensina II. Comenzaré por el glucagon.
Esta hormona es un polipéptido con un peso molecular de 3 500 Daltones aproximadamente que se produce en las células alfa de los islotes de Langerhans del páncreas y es una de las principales hormonas hiperglicemiantes, es decir, aquellas que aumentan los niveles de glucosa en la sangre. Recordemos que la insulina también es producida en los islotes pancreáticos y ejerce una acción opuesta. Estas pequeñas islas de células de secreción interna se encuentran dentro de la estructura del páncreas rodeadas por las células productoras de los jugos digestivos y su función es regular la concentración de glucosa en la sangre. Son un sistema de ajuste muy fino de la glucemia; cuando el nivel de glucosa en sangre se eleva, aumenta la secreción de insulina, la cual, como ya hemos descrito, favorece la entrada de este azúcar a las células y su posterior metabolismo, disminuyendo en consecuencia su concentración en la sangre. Por el contrario, cuando la glucemia baja, los islotes secretan glucagon, el cual se encarga de "indicar" al hígado que aumente su producción de glucosa, y al tejido adiposo que libere ácidos grasos y glicerol como fuentes de energía y material para la síntesis hepática de más glucosa. Visto así, nos percatamos de que estos islotes son un regulador de la glucemia; podríamos llamarles un glucostato. Los islotes se pueden aislar e incubar por tiempo relativamente largo sin que se deterioren. Se ha pensado en transplantarlos a pacientes diabéticos como una alternativa terapéutica y miles de estudios han sido realizados con ellos.
Pero volvamos al glucagon. Esta molécula fue descubierta por Murlin y colaboradores en 1923, tan sólo dos años después del descubrimiento de la insulina. Sin embargo, su historia ha sido un poco gris; en compensación con la historia de la insulina, su vecina de las células de al lado, que es brillante, espectacular, maravillosa, como las grandes avenidas: llena de luces. El descubrimiento del glucagon recibió poco reconocimiento; para ilustrar el escaso interés que generó diré que pasaron más de 30 años desde su descubrimiento hasta su purificación. Para todo existe una razón. ¿A qué se debió el contraste de interés y avance entre insulina y glucagon? Una razón importante es la rápida aplicación terapéutica, la utilidad. La insulina podía emplearse para el tratamiento de uno de los grandes males de la humanidad, la diabetes. El glucagon no tiene la misma suerte, su utilidad en la práctica médica es limitada: sólo se utiliza para algunos casos de hipoglucemia y recientemente para algunos problemas cardiovasculares. Ni modo, una nació con estrella y la otra estrellada. Lo que ha dado popularidad al glucagon es el hecho de que su acción en algunas condiciones es adversa. Voy a explicarme. Por allá en el inicio de los setenta, al desarrollarse las técnicas de detección de hormonas en el suero, se observó que en algunos diabéticos los niveles de insulina no estaban disminuidos como se suponía; estaban normales, pero los de glucagon estaban elevados. Así, el grupo de Unger propuso que el glucagon quizá era responsable en parte de los problemas del diabético. Hoy sabemos que la diabetes es una enfermedad (o grupo de enfermedades) mucho más compleja de lo que pensábamos. Muchos factores contribuyen, y posiblemente el exceso en la secreción de glucagon sea uno de ellos.
Para 1972 se había logrado la síntesis química del glucagon y con ello se empezaron a sintetizar compuestos similares (análogos) al glucagon con dos ideas fundamentales: conocer qué partes de su estructura son importantes para su actividad y, por otro lado, sintetizar antagonistas que pudieran bloquear las acciones de glucagon sobre el organismo y así poder, tal vez, aliviar algunos de los problemas del diabético.
Hasta el momento sólo se conoce un receptor para el glucagon, el cual está acoplado en forma activadora a la adenilil ciclasa y, en apariencia, en forma secundaria a la movilización de calcio, probablemente a través del sistema fosfoinosítidos-calcio. Al activar el glucagon a su receptor en el hígado, se produce un incremento rápido en los niveles de AMP cíclico, el cual, a su vez, activa a la proteína cinasa A y mediante la cascada de fosforilaciones, que describí capítulos atrás, se activan la ruptura de glucógeno y otros procesos.
La vasopresina, como su nombre lo indica, es una potente hormona vasopresora, es decir, aumenta la contracción de los vasos y la tensión arterial. También recibe el nombre de hormona antidiurética, mismo que describe otra de sus acciones principales: disminuir la diuresis, es decir, la pérdida de líquidos por la orina.
La vasopresina u hormona antidiurética es liberada por la hipófisis posterior (una glándula neurosecretora localizada en la base del cerebro) cuando la ingestión de líquidos es poca y cuando disminuye el líquido extracelular. Esta hormona es un péptido pequeño formado sólo por nueve aminoácidos. Su estructura fue determinada por DuVigneaud, quien además la sintetizó, por lo que se le otorgó el premio Nobel poco después de su descubrimiento. Este investigador y su grupo estudiaron también la oxitocina, una hormona muy parecida a la anterior (sólo cambia en dos aminoácidos), responsable de la contracción uterina durante el parto.
Esta hormona antidiurética se utiliza para el diagnóstico de la diabetes insípida (no confundir a esta enfermedad con la diabetes mellitus, la diabetes común que todos conocemos). Se utiliza también como medicamento para controlar el sangrado de várices esofágicas en los pacientes cirróticos.
La vasopresina ejerce sus efectos al activar a dos tipos de receptores: los V1 y los V2. Los efectos presores se ejercen al activar a los receptores V1, de los que existen dos subtipos (A y B) y los cuales están acoplados a recambio de fosfoinosítidos y a movilización de calcio. Los efectos antidiuréticos se ejercen mediante receptores V2 que están acoplados en forma activatoria a la adenilil ciclasa a través de Gs. Actualmente contamos con análogos sintéticos de la vasopresina, los cuales son selectivos hasta por 1 000 veces para algunos de los subtipos de receptores de dicha hormona. No debe pensarse que la vasopresina no tiene efectos en otros órganos; se ha observado que es un importante modulador del metabolismo hepático, además de tener efectos sobre las plaquetas e incluso sobre algunas neuronas.
Hace casi un siglo, en 1898, se descubrió que el extracto del riñón produce un fuerte efecto vasopresor; a este principio se le dio el nombre de renina. Posteriormente se descubrió que la renina no era vasopresora por sí misma, sino que era una enzima que convertía a un producto inactivo del plasma, el angiotensinógeno, en uno activo, la angiotensina. La angiotensina es una hormona que descubrieron el doctor Braun-Menéndez y su grupo, en Argentina hacia 1939. Casi al mismo tiempo, el grupo de Page, hizo el mismo descubrimiento. Cada uno de estos grupos dio un nombre al compuesto generado en el plasma, el primero lo llamó "hipertensina" y el segundo "angiotonina". Fue necesario que pasaran casi 20 años, para que se pusieran de acuerdo los investigadores del campo en el nombre adecuado para la hormona, y en 1957 se le dio el nombre híbrido de angiotensina.
Actualmente sabemos que hay tres angiotensinas: la I, la II y la III, las cuales son productos cada vez más pequeños; es decir, del angiotensinógeno se forma la angiotensina I, de ella la angiotensina II y de ésta a su vez la angiotensina III; la más activa es la angiotensina II.
La angiotensina II es el agente vasopresor más potente que se conoce hasta ahora; además es un importante regulador de la secreción de otra hormona por la corteza de la glándula suprarrenal: la aldosterona. En el corazón, la angiotensina II incrementa el flujo de calcio y la fuerza de contracción del músculo. Otra acción importante de este péptido es favorecer la secreción de la vasopresina. Durante los últimos años ha resultado evidente que este compuesto es también capaz de alterar el metabolismo hepático.
Pero ¿cómo ejerce sus efectos? La angiotensina II tiene receptores específicos localizados en la membrana plasmática de muchas células. Se han identificado dos tipos de receptores para la angiotensina II. El AT1 de distribución periférica y el que media las acciones más conocidas del péptido. En algunas especies el gen que codifica para el receptor se ha duplicado y modificado dando origen a dos variedades de este tipo de receptor (A y B). El receptor está acoplado al recambio de fosfoinosítidos a través de Gq y a la adenilil ciclasa en forma inhibidora por medio de Gi. Además parece estar asociado a otros sistemas de transducción, que le permiten actuar como un poderoso factor de crecimiento en muchas células.
El receptor AT2 parece estar acoplado a través de proteínas G (aún no bien identificadas) a una fosfatasa de proteínas para residuos de tirosina. Este parece ser un nuevo sistema transductor que ahora se está estudiando con mucho interés. Por otro lado, el receptor AT2 parece expresarse en el sistema nervioso y especialmente en ciertas etapas del desarrollo, lo que ha sugerido que ocupe un lugar en la diferenciación de este importante sistema. Nuevamente otro campo en el que veremos cambios en los próximos años.