VIII. CÓMO SE ESTUDIAN LAS MEMBRANAS
DURANTE muchos años esencialmente se hicieron estudios en membranas celulares naturales obtenidas por diferentes procedimientos. Uno de los métodos tal vez más antiguos que exista es el de la preparación de membranas de glóbulos rojos. La técnica para obtenerlas es muy sencilla; simplemente se colocan estos corpúsculos sanguíneos en una solución muy diluida de algunas sales, y debido a la presión osmótica (véase el Capítulo VII), se rompe su membrana y el contenido escapa al exterior. Sin embargo, si el procedimiento se realiza en condiciones especiales, las membranas vuelven a sellarse y pueden separarse luego por centrifugación. Estas membranas se ven transparentes al microscopio por haber perdido su color y se les denomina "fantasmas" de eritrocitos.
Por diferentes procedimientos ha sido posible preparar membranas naturales de muchos otros orígenes, tanto de las células como de diferentes componentes u organelos. Pero hace aproximadamente 20 años se iniciaron estudios utilizando membranas llamadas "artificiales", debido a que no provenían directamente de las células. Su nombre en realidad no está bien aplicado, pues los fosfolípidos, a partir de los cuales se forman, sí se extraen, ya sea de células o de otros sistemas biológicos.
Si se quieren hacer estudios sobre la composición de las membranas es necesario aislarlas, pero cuando se trata de conocer su funcionamiento, se pueden utilizar desde células enteras o tejidos hasta componentes intracelulares, como pueden ser las mitocondrias, lisosomas, retículo sarcoplásmico, etc. De cualquier manera, ya sea que se trate de hacer los estudios en células u organelos aislados, los pasos para obtener una y otras preparaciones implican primero la separación de los componentes, ya sean las células de los tejidos, los organelos de las células, o bien las membranas mismas obtenidas de diferentes partes de las células o sus organelos.
Para obtener los componentes de un tejido (Figura 54), el procedimiento inicial
más común que existe es disgregarlos. Así, por ejemplo, si se busca obtener
células enteras, lo que se hace con frecuencia es utilizar algún producto capaz
de digerir el material intercelular que las mantiene unidas. Para separar células
hepáticas, por ejemplo, se utiliza una enzima llamada colagenasa, que permite
la separación de las células.
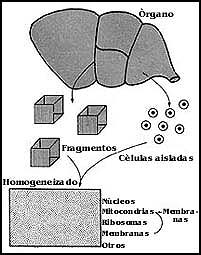
Figura 54. Diagrama general sobre la separación de los componentes celulares.
Si lo que se trata de separar son las mitocondrias u otros componentes intracelulares, a lo que se debe recurrir es a diferentes procedimientos de ruptura de las células, que en la jerga de laboratorio reciben el nombre de homogeneización. Los procedimientos son diversos según la célula de que se trate, pero uno de los más frecuentes consiste en el empleo del homogeneizador, que es un tubo de vidrio en cuyo interior se colocan las células o fragmentos del tejido que se desea homogeneizar. A continuación, al tiempo que se le imprimen movimientos hacia dentro y hacia fuera, se hace girar en su interior un vástago que puede ser de vidrio o de teflón, que termina por remoler a las células, dejando intactos a algunos de sus componentes, como núcleos, mitocondrias, lisosomas, retículo sarcoplásmico, etc. Pero también se pueden utilizar otros sistemas, como licuadoras, semejantes a las que se emplean para preparar alimentos, la agitación con perlas de vidrio o abrasivos, la vibración ultrasónica, la compresión y descompresión u otros aparatos.
Hay células como las de algunos microorganismos o plantas que tienen una pared celular que no es tan fácil de romper, y para preparar o separar los componentes intracelulares deben utilizarse procedimientos un tanto mas drásticos. Para el efecto se han diseñado diferentes instrumentos, cuyo funcionamiento no viene al caso discutir en este momento. También se utilizan enzimas capaces de digerir la pared celular antes de romper las células.
MÉTODOS DE SEPARACIÓN DE LOS ORGANELOS SUBCELULARES
Existen muchos métodos, pero uno de los más utilizados en el laboratorio consiste en separar los distintos componentes de las células, o las células mismas, aprovechando sus diferentes densidades, y el hecho de que, por lo tanto, la velocidad con que sedimentan los distintos componentes varía de acuerdo con esa densidad.
La figura 55 muestra en forma simplificada el procedimiento que puede utilizarse
para separar los componentes de una célula, y que consiste esencialmente en
dos sistemas; en el más usado de ellos, llamado de centrifugación diferencial,
se realizan centrifugaciones a distintas velocidades y tiempos para obtener
sus componentes. Por ejemplo, para sedimentar las mitocondrias de células hepáticas
basta con centrifugar alrededor de 10 minutos el homogeneizado a una velocidad
que produzca algo así como 8 000 veces la fuerza de la gravedad. Pero antes
de eso, será necesario sedimentar partículas más pesadas como los núcleos. Entonces,
lo que se hace primero es centrifugar a una velocidad menor y después sedimentar
las mitocondrias. Si lo que se trata de obtener son los ribosomas de las células,
es necesario centrifugar durante aproximadamente 30 minutos a una velocidad
que genere algo así como 100 000 veces la fuerza de la gravedad.
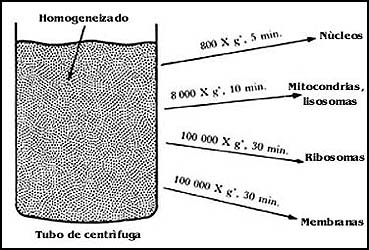
Figura 55. La sedimentación de componentes celulares por centrifugación.
Otro de los procedimientos consiste en la preparación en un tubo de centrífuga de una solución que puede ser de sacarosa o de otras sustancias, en la cual la concentración de la sustancia disuelta va aumentando de arriba a abajo en el tubo. Esto equivale a que existan entonces diferentes densidades a distinta altura. Si luego se coloca un homogeneizado o una preparación un poco más pura obtenida de alguna célula, y se le somete a una centrifugación, cada una de las partículas presentes en el homogeneizado se va a detener en el punto en donde la densidad del líquido sea igual a la propia. Así, los núcleos viajarán hacia la parte inferior del tubo, las mitocondrias se quedarán en una zona intermedia por arriba de la anterior, y los ribosomas en una zona todavía más arriba.
El procedimiento que se llama de centrifugación en un gradiente de concentración, en ocasiones permite separar componentes celulares que tienen densidades relativamente semejantes.
Como ya se mencionó, las propiedades de las membranas de las partículas subcelulares pueden estudiarse utilizándolas intactas; se pueden realizar los experimentos directamente con las partículas obtenidas en la centrifugación. Sin embargo, si lo que se busca es conocer más detalladamente las propiedades de las membranas, y en especial sus componentes, es necesario en principio utilizar un procedimiento semejante al ya mencionado para la preparación de las membranas de los glóbulos rojos; el procedimiento consiste en la ruptura de las membranas y su separación posteriormente, casi siempre por centrifugación diferencial. En todos los casos, cuando se hace la ruptura, cambian las densidades respecto a las células u organelos intactos, y las velocidades de centrifugación y los tiempos utilizados tienen que ser más largos.
Las técnicas para la separación de los componentes celulares y sus membranas han evolucionado a tal grado, que ya en esta época es posible obtener una preparación de membranas puras de prácticamente cualquier sistema biológico.
ANÁLISIS DE LOS COMPONENTES DE LAS MEMBRANAS
Los lípidos. Para estudiar la composición lipídica de una membrana, el método en general simplemente aprovecha las propiedades de solubilidad de los lípidos, y dado que algunos son más o menos solubles en ciertos líquidos o solventes, según la membrana, se utilizan extracciones; es decir, la solubilización de los lípidos usando distintos tipos de solventes o mezclas de ellos como cloroformo, alcohol metílico, hexano, acetona, etcétera.
Después de la extracción de los lípidos suele procederse a su separación e
identificación. Para ello hay un número enorme de técnicas, mediante las cuales
se pueden separar los componentes, pero probablemente el más utilizado sea la
separación por cromatografía, ya sea en una placa o en una columna. La Figura
56 muestra en forma esquemática el principio de la cromatografía. Tanto para
los lípidos, como para otras sustancias, se les puede colocar ya sea en una
placa recubierta de un material especial o en una columna o tubo de vidrio lleno
con el material que se desea analizar. Por sus propiedades de solubilidad y
carga, las moléculas se fijan con diferente fuerza a las partículas de la sustancia
que se ha utilizado para preparar la placa o la columna. Entonces se procede
a pasar una corriente de algún solvente o mezclas de solventes que arrastran
con distinta velocidad a los componentes que se han colocado en el sistema.
Debido a estas velocidades diferentes, las diversas moléculas se separan formando
bandas o manchas que pueden detectarse utilizando colorantes, oxidantes o simplemente
por calentamiento a temperaturas altas.
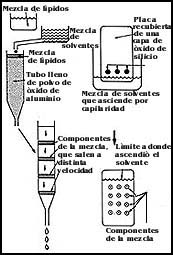
Figura 56. Esquema de dos tipos de cromatografía para separar mezclas de sustancias.
El empleo de las columnas de cromatografía permite no solamente la separación de las sustancias, sino eventualmente aislarlas en cantidades más importantes. Una vez que se cuenta con cantidades adecuadas de una sustancia, los procedimientos son muy variados. Hay un gran número de métodos analíticos que pueden ser más o menos complicados, que finalmente pueden llevar a definir su estructura.
Las proteínas. Los procedimientos empleados para estudiar o aislar,
inclusive purificar las proteínas de las membranas biológicas, no son en esencia
diferentes a los utilizados para separar los lípidos. Simplemente sucede que,
dado que las propiedades de las proteínas son diferentes a las de los lípidos,
los métodos utilizados para separarlas y aislarlas de las membranas son diferentes.
En el caso de las proteínas de la membrana que se encuentran en contacto con
los fosfolípidos, uno de los procedimientos que más se utiliza, consiste en
la extracción de las mismas con detergentes. Por tener éstos una estructura
más o menos semejante a la de los fosfolípidos, pueden sustituirlos y extraer
a las proteínas de las membranas. En forma muy general, lo que se extrae de
las membranas cuando se usa un detergente es la mezcla de las proteínas que
éstas contienen, pero en lugar de estar asociadas a los fosfolípidos se extraen
rodeadas de una capa de detergente (Figura 57).
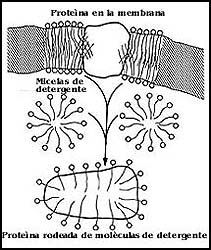
Figura 57. Esquema de la extracción de una proteína de la membrana con un detergente.
Dada la existencia de esta diferencia fundamental entre las proteínas de la membrana que están asociadas a moléculas de lípidos, y las proteínas de las células que en general se encuentran en el seno de un medio acuoso, los procedimientos que suelen utilizarse para separar una de otras y eventualmente purificarlas son semejantes a los empleados en la separación de otras proteínas. También en este caso se hace uso de columnas de cromatografía, de manera semejante como se mencionó para los lípidos; pero es obvio que los materiales utilizados para separar proteínas deben ser diferentes a los materiales utilizados para la separación de los lípidos. También sucede que, como en el caso de los lípidos, en la actualidad es factible, al menos en principio, purificar cualquier proteína de la membrana si se le da un tiempo y un esfuerzo suficientes.
Los carbohidratos. Dado que el contenido de carbohidratos en las membranas celulares no es tan grande como el de lípidos y proteínas, y varía no sólo en la proporción sino en el tipo de azúcares que contienen las membranas, no hay procedimientos universales para separarlos, principalmente por el hecho de que con frecuencia los carbohidratos se encuentran asociados a las proteínas. En el caso de los azúcares, por lo tanto, los procedimientos son todavía más diversos, pero también es posible separarlos, aislarlos y estudiarlos por métodos analíticos.
ESTUDIO DE LAS FUNCIONES DE LAS MEMBRANAS
Como en el caso de los componentes y algunas otras propiedades, es posible estudiar las funciones celulares utilizando distintos tipos de preparaciones. Se pueden emplear células enteras u organelos intracelulares, pero cuando se estudian las funciones, una de las preparaciones que tiene mayor utilidad son las vesículas obtenidas a partir de distintos tipos de membranas. Un ejemplo es el procedimiento para preparar vesículas a partir de mitocondrias mediante el ultrasonido, en el que ocurre la ruptura de las mitocondrias principalmente debido a la estructura que éstas tienen, dando lugar a las llamadas partículas submitocondriales, que además de estar constituidas esencialmente por los mismos componentes de las membranas de la mitocondria entera, están dispuestas en forma invertida en su mayoría, es decir, lo que era el interior de la mitocondria es ahora el exterior de la partícula y viceversa.
Hay muchos procedimientos de ruptura de las membranas para preparar vesículas. Como en el caso anterior, algunos producen vesículas en las que se invierte la polaridad original u orientación de los lados de la membrana, pero en otros ésta se conserva. Es así como se puede "jugar", por así decirlo, con los procedimientos, para preparar diferentes tipos de membranas a partir de los distintos sistemas biológicos que se estudian.
LOS LIPOSOMAS Y LOS PROTEOLIPOSOMAS
Tal vez una de las preparaciones que mayor información ha proporcionado en los últimos años dentro del estudio de las funciones de las membranas es la de los liposomas o proteoliposomas. Son vesículas que se forman utilizando diferentes fosfolípidos mediante distintos procedimientos, pero el más común es el siguiente: tomando una cierta cantidad de fosfolípidos, se les coloca en una solución adecuada a los estudios que se desean realizar y se les somete a un tratamiento variable según la preparación requerida, utilizando ultrasonido, es decir una vibración de muy alta frecuencia que tiene como objetivo fundamental dispersar a las moléculas en el líquido, y producir finalmente vesículas como las que se describieron en el capítulo II, formadas por una doble capa de fosfolípidos, con una cavidad líquida en su interior. Como también se mencionó en el capítulo II, estas vesículas tienen la estructura básica de las membranas celulares y comparten con ellas las propiedades fundamentales, principalmente en cuanto a permeabilidad.
Pero la utilidad más grande de los liposomas deriva de la posibilidad que hay
de purificar las proteínas de las membranas y de incluirlas o reincorporarías
a ellos, para formar los llamados proteoliposomas. De esta forma es como se
completa con frecuencia el proceso mediante el cual se demuestra que algún componente
de la membrana tiene cierta función. En principio, la manera ideal de lograr
esto consiste en aislar a la proteína que se supone responsable de un fenómeno,
incorporarla aislada a un liposoma, y demostrar que en efecto, cuando se le
ha aislado e incorporado en estas condiciones, es capaz de realizar la función
que hipotéticamente se le había atribuido (Figura 58).
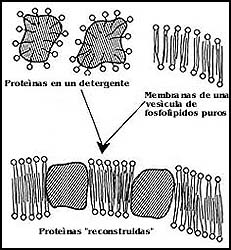
Figura 58. La incorporación de proteínas aisladas en vesículas de fosfolípidos puros.
Por ejemplo, la Figura 59 muestra algunos casos, desde el más simple y ya descrito
anteriormente de la valinomicina, de la cual se dice que es capaz de mover con
gran selectividad iones de potasio a través de las membranas biológicas. Si
se incorpora este antibiótico a liposomas que se han preparado en un medio rico
en potasio, y están llenos de éste, y se coloca luego a las vesículas en un
medio libre de potasio, es posible demostrar que, en efecto, el potasio sale
de las vesículas cuando se agrega el antibiótico.
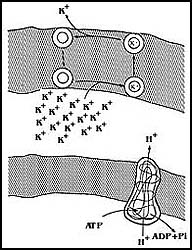
Figura 59. Esquema de dos sistemas de transporte que se hacen funcionar en liposomas formados con fosfolípidos puros.
Otro ejemplo que también se muestra en la figura es el de la ATpasa mitocondrial,
que se supone es capaz de romper moléculas de ATP y simultaneamente
bombear protones o hidrogeniones de un lado al otro de la membrana, o bien,
cuando se genera una diferencia suficientemente grande de la concentración de
hidrogeniones en ambos lados de la membrana, la enzima puede sintetizar ATP
a partir de sus componentes, el ADP y el fosfato. El experimento fue realizado
hace ya muchos años por Ephraim Racker, aislando la enzima e incorporándola
en las vesículas de fosfolípidos obtenidos de la soya.
Otro de los métodos que se han utilizado para hacer experimentos más o menos
semejantes a los planteados con los liposomas, consiste en la formación de membranas
abiertas, pero que se pueden estudiar según el sistema presentado en la Figura
60, y que estriba esencialmente en preparar una solución de fosfolípidos en
un solvente adecuado y colocar una gota de éste con un pincel sobre el orificio
de una pieza de plástico, que suele ser de teflón. El resultado del procedimiento
es el que se describe en la misma figura, y consiste en la formación de una
doble capa de fosfolípidos en el pequeño orificio de la pieza de plástico, que
parece conservar todas las propiedades de tina doble capa de fosfolípidos.
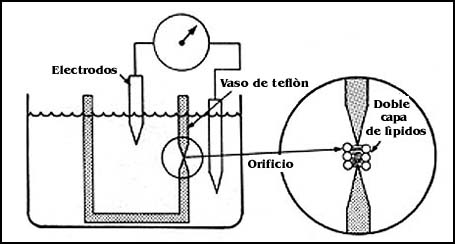
Con este sistema también es posible estudiar las propiedades de la doble capa de los fosfolípidos, así como de incorporar a ella distintos tipos de sustancias o proteínas, y demostrar ciertas funciones que puedan interesar al investigador.
Hay diferentes ventajas en las membranas planas con relación a los liposomas. En éstos, el volumen exterior es extremadamente pequeño en relación con el exterior, y no se pueden hacer estudios directos de sus cambios y algunas otras propiedades. En el caso de las membranas planas, si el orificio se coloca en medio de dos cámaras, la bicapa que se forma puede estudiarse utilizando electrodos o el transporte de sustancias cargadas; ésto se logra detectando con los electrodos el paso de iones a través de la bicapa, que se manifiesta de la misma forma como si se tuviera el paso de corriente a través de la doble capa de fosfolípidos.
ESTUDIOS SOBRE EL TRANSPORTE A TRAVÉS DE LAS MEMBRANAS
Una de las funciones más importantes y probablemente más estudiadas en las membranas biológicas es la del transporte de distintos tipos de sustancias. Se han diseñado distintos métodos experimentales para estudiarlo; sin embargo, todos tienen en principio la misma base de operación. Lo más importante para seguir el paso de una sustancia a través de una membrana, o su incorporación a una vesícula, es contar con un método para detectarla. Para algunos estudios de transporte se cuenta con muy diversos procedimientos analíticos, con el objeto de medir, ya sea su incorporación a una vesícula o a un organelo, o su paso a través de una bicapa plana.
No obstante que los procedimientos analíticos son extremadamente diversos, en muchísimos casos uno de los sistemas más utilizados es el empleo de isótopos radiactivos que se pueden seguir con relativa facilidad, estudiando simplemente la radiactividad incorporada o transportada de un lado a otro de una membrana o al interior de una vesícula u organelo, etcétera.
Para estudiar el transporte de sustancias en las membranas, dado que en general se tienen vesículas u organelos cerrados, el procedimiento consiste en colocar a éstos en presencia del material que supuestamente se va a transportar, dejar transcurrir un cierto tiempo que puede ser desde unos cuantos segundos hasta horas, y luego separar las vesículas del medio en que se les ha colocado por distintos procedimientos. La Figura 61 muestra en forma esquemática los procedimientos más frecuentes en este tipo de estudios que consisten en la separación de las vesículas por filtración, por centrifugación, o por el empleo de columnas. En todos los casos el principio es esencialmente el mismo, y se trata de obtener aisladas las vesículas y el medio en que se habían colocado, para detectar diferencias entre el estado inicial y el logrado después de un cierto periodo de incubación.
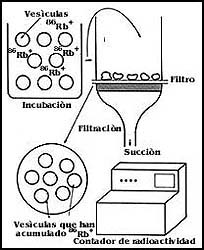
También para el estudio de los receptores, dado que lo que se trata en muchos de los casos de la interacción de una sustancia con estas moléculas, los procedimientos empleados son semejantes a los utilizados en el transporte. En este caso es necesario también contar con métodos para detectar, con frecuencia, concentraciones sumamente bajas de las sustancias que se fijan a los receptores. Igualmente para el estudio de los receptores las sustancias que mayor utilización tienen son las marcadas con isótopos radiactivos.
Los procedimientos son semejantes a los empleados para estudiar el transporte. La diferencia entre un transportador y un receptor es simplemente que en el caso del transportador la sustancia utilizada se transfiere de un lado a otro de la membrana, y en el caso de un receptor ésta suele fijarse sólo de un lado de la membrana, pero de todas formas la fijación se hace con tal intensidad que al separar la preparación membranal del medio en que se ha incubado es posible detectar cantidades variables de las sustancias unidas a los receptores.
Del mismo modo que para los estudios del transporte, los procedimientos de separación de las membranas del medio en que se han colocado durante la etapa experimental suelen ser la filtración y la centrifugación.
Hemos revisado algunos de los procedimientos más comunes que se utilizan en el estudio de las membranas biológicas. Sin embargo, debe quedar claro que hay una gran cantidad de funciones de las membranas, y que los enfoques utilizados para su estudio pueden ser de lo más diverso. Por ejemplo, durante el funcionamiento de algunos sistemas de transporte es frecuente que se generen diferencias de potencial eléctrico, es decir, diferencias de concentración de cargas en ambos lados de una membrana.
Estas diferencias de potencial se pueden estudiar utilizando, por ejemplo, membranas planas y electrodos colocados a ambos lados de ellas. Pero también pueden estudiarse las diferencias de potencial empleando células, liposomas o proteoliposomas, valiéndose de la propiedad que algunas sustancias tienen para cambiar sus características físicas, cuando se mueven de un lado a otro de la membrana, llevadas por una diferencia de potencial.
Por ejemplo, es posible detectar cambios de potencial eléctrico en una membrana
utilizando el colorante llamado oxonol V. Ésta es una molécula fluorescente
que puede moverse con cierta libertad a través de las membranas biológicas,
y si hay un potencial positivo dentro de una vesícula, la sustancia se acumula
dentro de ella. Al acumularse en el interior de la vesícula, la fluorescencia
de algunas de las moléculas es absorbida por las otras debido a su cercanía
por la misma acumulación de que han sido objeto, y si se mide ésta en un aparato
adecuado, un fluorómetro, es posible detectar en el tiempo los cambios de potencial
por los cambios de fluorescencia de esta sustancia (Figura 62).
Figura 62. Medida de la diferencia de potencial eléctrico por la acumulación de una sustancia fluorescente.
Así como hay indicadores capaces de seguir diferencias de potencial entre los lados de una membrana, se pueden también estudiar cambios en la concentración de hidrogeniones con sustancias adecuadas, en general colorantes o indicadores fluorescentes. También es posible utilizar algunos indicadores que son moléculas capaces de sufrir cambios de color o de fluorescencia cuando se fijan a determinadas sustancias, como el calcio, el magnesio, u otros iones.
Hay, en resumen, un número enorme de técnicas que se pueden utilizar en el estudio de las membranas solamente en el área del transporte biológico. Baste decir únicamente que igual que para otros estudios y actividades, un investigador científico no tiene más límites que su imaginación para diseñar los métodos que le permitan plantear los experimentos adecuados, para conocer la verdad y los detalles de algún proceso biológico.
Por otra parte, con frecuencia no solamente se busca conocer las propiedades funcionales de un componente de una membrana. En esta época son numerosos los casos en los cuales se busca conocer la regulación genética de la cantidad o las propiedades de un determinado transportador. Hay muchos estudios de sistemas de transporte en los cuales se utilizan técnicas genéticas, ya sea en cultivos de células o en microorganismos o inclusive las opciones que brinda la biología molecular y la ingeniería genética para producir cambios deseables en un sistema membranal.
Los estudios en el área de las membranas biológicas utilizan herramientas desarrolladas por muchas otras ciencias que incluyen a la genética, la física, la química, la biología molecular, la ingeniería genética, etc. Son tantas las facetas que requiere el estudio de éstos o cualesquiera otros sistemas o fenómenos biológicos, que prácticamente no hay ni debe haber limitación alguna en cuanto a las ciencias o herramientas que los investigadores utilicen para profundizar cada vez más sobre el conocimiento de esos fenómenos.