IV. PROPAGACIÓN INTRACELULAR Y AMPLIFICACIÓN DE LA SEÑAL
EN LOS CAPÍTULOS ANTERIORES se ha visto que el acoplamiento del mensajero con su receptor en la membrana plasmática es el primer paso para que se lleve a cabo la acción de algunas hormonas. También se ha aclarado que no basta solamente con el acoplamiento hormona-receptor, sino que es necesario que el receptor se active y que, a su vez, se asocie con un sistema transductor, el cual se encarga de generar la señal intracelular o segundo mensajero. En este capítulo se explicará cómo ejercen su acción estos segundos mensajeros; es decir, cómo se propaga y amplifica la señal en el interior de la célula, y se ilustrará, con algunos ejemplos, el funcionamiento del sistema.
Los segundos mensajeros no actúan directamente, sino que son reconocidos ("oídos") por receptores extracelulares. Así como el primer mensajero, la hormona, es "oído" por la célula a través de sus receptores, la señal intracelular o segundo mensajero es "oído" por estos receptores intracelulares. Es decir, el primer paso es el reconocimiento molecular para después pasar a la acción. Dicho reconocimiento es llevado a cabo por proteínas, y la información precisa para su síntesis se encuentra codificada en nuestro material genético. Estas proteínas reconocen al segundo mensajero con una extraordinaria afinidad y especificidad y participan en la propagación de la señal. Dicha propagación no es tampoco un proceso lineal, sino que se lleva a cabo en forma de "cascada de amplificación"; esto quiere decir que, en cada paso que se da, el proceso se va haciendo más amplio, más grande.
Ilustraré lo dicho con un ejemplo para aclarar los conceptos. Imaginemos que un oficinista está en un ayuno más o menos prolongado; se le ha descompuesto el coche y no puede ir a su casa a comer, por lo que no ingerirá alimento hasta la cena. Durante el desayuno, el individuo consumió una cierta cantidad de azúcares, de los cuales, una pequeña parte fue utilizada por el organismo y el resto se almacenó en el hígado en forma de glucógeno (el carbohidrato de reserva de los animales constituido por múltiples moléculas de glucosa unidas). Sin embargo, nuestro amigo oficinista necesita que durante todo el tiempo la concentración de glucosa en su sangre permanezca más o menos constante para que los tejidos puedan nutrirse debidamente. Esto es especialmente importante en el caso del sistema nervioso, pues si la glucosa baja de cierto nivel se presentan diversos trastornos hasta llegar a la pérdida de la conciencia o coma. Si a nuestro oficinista le disminuye notablemente la concentración de glucosa, su capacidad de atención disminuirá. (Nota: esto ha llevado a muchos a pensar que la falta de atención, que observamos cotidianamente en algunas oficinas públicas, se debe a disminuciones importantes en los niveles de glucosa en la sangre, por lo que han proliferado las horas de desayuno, las dos horas de la comida, los 30 minutos del café, los 25 minutos del refresco..., etc. A pesar de tales suplementos nutricionales, el problema subsiste con exasperante insistencia, lo cual indica, sin temor a equivocarse, que la causa debe ser otra. Sin embargo, las prácticas de nutrición continua en las oficinas públicas parecen haber llegado para quedarse.) Pero, volviendo al hilo de nuestra historia, decíamos que el hígado acumula glucosa en forma de glucógeno, y que este órgano es el encargado de regular los niveles de glucosa en la sangre. Naturalmente, el hígado no hace todo por sí solo. En el páncreas existe un tipo de células muy especializadas, las cuales son lo que podríamos llamar los "sensores" de la glucemia. Cuando los niveles de glucosa en la sangre disminuyen, secretan una hormona llamada glucagon, la cual avisa al hígado que el organismo necesita que libere glucosa al torrente sanguíneo.
Pero, ¿cómo actúa el glucagon? Básicamente activando a la adenilil ciclasa
(a través de su receptor, que estimula a la proteína Gs, que a su vez induce
la activación de la enzima, como ya hemos visto), y por lo tanto aumentando
la producción de AMP cíclico. Pero aquí estamos entrando de lleno
en lo que vamos a llamar propagación y amplificación de la señal, una vez dentro
de la célula. Este proceso es complejo, por lo que lo analizaremos detenidamente.
Permítaseme, entonces, hacer otro paréntesis.
En el citoplasma celular existe un tipo de proteínas llamadas "proteínas cinasas", las cuales fosforilan (le añaden fosfato) a algunas proteínas de la célula, de modo tal que, al ser fosforiladas, dichas proteínas cambian en su actividad, unas se inhiben y otras se activan. Este proceso es uno de los mecanismos moleculares de regulación de la función celular más importantes que existen, y participa prácticamente en todas las funciones celulares: la regulación del metabolismo, la contracción, la secreción, la proliferación celular, la diferenciación, etc. Los investigadores pioneros que descubrieron el papel de la fosforilación de proteínas en la regulación de la función celular fueron Edmond Fisher y Edwing G. Krebs y recibieron conjuntamente el Premio Nobel de Medicina y Fisiología hace algunos años.
Dicho esto, prosigamos con el mecanismo de acción del glucagon. El AMP
cíclico generado es reconocido por su receptor en el citoplasma: la proteína
cinasa A (para el AMP cíclico). Esta proteína cinasa tiene dos
tipos de subunidades (véase la figura 9), las "R" (reguladoras) y las "C" (catalíticas,
las cuales son las que tienen la actividad de proteínas cinasa propiamente dicha).
Las subunidades "R" mantienen inhibida a la enzima. Cuando los niveles de AMP
cíclico aumentan, éste se pega a las subunidades e induce un cambio conformacional
tal en esta subunidad, que la afinidad por "C" disminuye, lo que conduce a la
separación de las subunidades "R" y "C". Ahora sí, las subunidades "C" quedan
libres para hacer de las suyas (véase nuevamente la figura 9). Como es de esperarse,
dada su actividad enzimática, la subunidad "C" de la proteína cinasa A se encarga
de fosforilar a algunas de las proteínas del citoplasma celular. Como decíamos
anteriormente, dichas proteínas, al ser fosforiladas, modifican su actividad
(la aumentan o la disminuyen); algunas de ellas son proteínas cinasas a su vez,
y al activarse van a fosforilar a otras proteínas, las cuales también se modifican
por este hecho y cambian su actividad... etcétera, etcétera. Así, sigue desencadenándose
esta "cascada" de fosforilaciones y, por lo tanto, amplificándose la señal en
el interior de la célula.
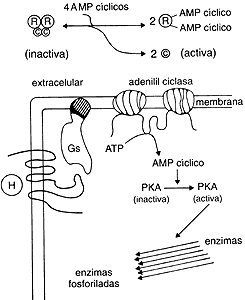
Figura 9. En la parte superior está una representación
de la activación de la proteína cinasa A (con sus subunidades reguladoras (R)
y catalíticas (C) por el AMP cíclico. En la parte inferior se presenta
la activación hormonal de la adenilil ciclasa y la propagación intracelular
de la señal. (PKA = proteína cinasa A.)
Aclaremos lo que sucede en el caso del glucógeno. Al activarse la proteína cinasa A, es decir, al quedar libre su subunidad "C", ésta produce la fosforilación de otra enzima: la fosforilasa b cinasa; esta enzima, a su vez, se activa y va a fosforilar a otra enzima: la fosforilasa; esta última rompe al glucógeno en múltiples moléculas de glucosa y permite que el hígado secrete glucosa a la circulación.
Como mencioné arriba, no todas las enzimas se activan al fosforilarse. La enzima que sintetiza el glucógeno, la "glucógeno sintetasa", se inhibe al ser fosforilada. Esto tiene gran importancia, ya que, de este modo, el mismo mensajero, el glucagon en el caso que nos ocupa, aumentó los niveles de AMP cíclico, lo que finalmente conduce coordinadamente a inhibir la síntesis de glucógeno y a aumentar su degradación. Esto hace que la liberación de glucosa por el hígado sea mucho más eficaz.
Es conveniente aclarar dos aspectos. Primero: así como los receptores tienen gran especificidad para reconocer a un mensajero, las enzimas la tienen para sus sustratos. La activación de las proteínas cinasas provoca la fosforilación de una serie de proteínas, pero, desde luego, no de todas las proteínas de la célula, solamente de aquellas que son sustrato de la cinasa. Segundo: el sistema se activa rápidamente, y también se desactiva con facilidad. Como hemos mencionado, los sistemas de señalamiento hormonal tienen la característica particular de tener una acción muy rápida, casi inmediata; pero también se desactivan rápidamente, al desaparecer la estimulación por la hormona. Imaginemos que nuestro oficinista sale a sus "45 minutos del chocolate con churros". Al recibir azúcares, su organismo los absorbe; sube la concentración de glucosa en su sangre y de inmediato cesa la secreción de glucagon por el páncreas. Por lo tanto, la concentración de glucagon en la sangre disminuye, deja de haber una cantidad suficiente para activar a los receptores hepáticos. La adenilil ciclasa del hígado ya no es activada por el receptor de glucagon y el AMP cíclico pasa a AMP no cíclico por la acción de la fosfodiesterasa. Las subunidades "R" y "C" de la proteína cinasa A se reasocian, con la subsecuente inhibición de su actividad. Las enzimas que habían sido fosforiladas, durante las cascada de amplificación, son desfosforiladas por enzimas encargadas de quitarles el fosfato (proteínas fosfatasas); regresando así a su actividad basal. Todo este complejo proceso se lleva a cabo en segundos.
He mencionado arriba que las enzimas se fosforilan por proteínas cinasas, y que también se desfosforilan por proteínas fosfatasas. Este juego de las cinasas y las fosfatasas es el que determina, en un momento dado, el estado de fosforilación de las enzimas clave o reguladoras de los procesos celulares. Es importante mencionar que, si bien el ciclo fosforilación/desfosforilación es quizá el proceso de regulación más importante de la célula, de ninguna manera es el único.
Veamos ahora la propagación de la señal intracelular por medio de otro sistema
de trasducción: el del calcio-inositol trisfosfato (IP3). Recordando
lo que decíamos anteriormente, en este sistema en particular el receptor activado
por la hormona estimula a una fosfolipasa C a través de proteínas de la familia
Gq. Esta fosfolipasa rompe el fosfatidilinositol bifosfato en dos productos:
el inositol trisfosfato y el diacilglicérido. El inositol trisfosfato, como
lo ha descrito Berridge, actúa sobre el retículo endoplásmico de la célula,
provocando la liberación de calcio. El calcio, libre entonces, ejerce su acción
corno segundo mensajero activando a una serie de proteínas cinasas y a otras
enzimas cuya acción depende de la concentración de este ion. Luego, la cascada
de amplificación de la señal se lleva a cabo por medio de la fosforilación de
enzimas, a semejanza de lo que describí anteriormente para el AMP
cíclico.
El calcio, libre también, puede actuar como segundo mensajero en la liberación de glucosa por el hígado. La hormona antidiurética, también llamada vasopresina, actúa básicamente a través del sistema Ca++ e IP3 en el hígado. Supongamos ahora que dicha hormona ejerce su acción sobre el hígado de nuestro oficinista; podríamos describir el desencadenamiento de los eventos como sigue: la vasopresina se acoplará a su receptor de membrana; éste se activará, estimulando a su vez a las proteínas Gq, las que a su vez modularán a la fosfolipasa C; de modo que ésta romperá al fosfatidilinositol bifosfato y se generará el inositol trisfosfato; éste, al actuar sobre sus receptores del retículo endoplásmico, provocará un aumento del calcio libre en el citoplasma, el cual activará la fosforilasa b cinasa que, a su vez, fosforilará a la fosforilasa, la cual romperá al glucógeno almacenando en la célula, liberándose así múltiples moléculas de glucosa.
Por otro lado, es importante señalar que el diacilglicérido, el otro producto
de la hidrólisis del fosfoinosítido, activa en forma conjunta con el calcio
a otra enzima: la proteína cinasa C (descubierta por el japonés Yasutomi Nishisuka).
Esta proteína colabora en el proceso de propagación y amplificación de la señal
hormonal, junto con las cinasas dependientes de calcio. En este mecanismo de
acción (Ca++ e IP3), al igual que en el del AMP cíclico,
la señal se transmite en segundos y se suspende también rápidamente; el diacilglicérido
y el inositol trifosfato son atacados por enzimas, las cuales los convierten
en compuestos inactivos.
Es necesario repetir que no sólo existen las proteínas cinasas A (activadas
por el AMP cíclico) y C (activada por el diacilglicerol), sino
que también existen, como ya mencionamos, las proteínas cinasas que amplifican
la señal en las cascadas de propagación intracelular y una creciente cantidad
de otras cinasas activadas por diversos factores. Veremos más adelante que hay
receptores con actividad de proteína cinasa, otros con actividad de fosfatasa,
y otros que generan segundos mensajeros como el GMP cíclico (que
activa a la proteína cinasa G). También es necesario mencionar que existen diversas
isoformas de las proteínas cinasas a las que nos hemos referido. Dicha existencia
nos hace preguntarnos ¿para qué se molestó la naturaleza en fabricar una variedad
de isoformas? Esto, casi en ningún caso lo sabemos, pero sí se conoce que hay
una expresión diferencial por tejidos, lo que nos lleva a suponer que las isoformas
se ocupan de funciones diferentes y, por lo tanto hay todo un campo de investigación
abierto para los aventurados en estos terrenos.
Para tener una mayor visión de lo que ocurre dentro de la célula también resultarán útiles algunas consideraciones generales. Las enzimas que se encargan de quitar los fosfatos, es decir, las fosfatasas, han sido estudiadas con gran interés durante los últimos años. Se sabe que existen, por lo menos, cuatro tipos diferentes de fosfatasas, y que hay dos inhibidores naturales de su acción. En los casos de la insulina y de algunos factores de crecimiento, se ha observado que, al ejercer su acción, aumentan la fosforilación de algunas proteínas en la célula, pero disminuyen la de otras. Estos datos sugieren que si bien las hormonas activan algunas enzimas fosforilantes (cinasas) posiblemente también activen fosfatasas.
El doctor Philip Cohen y su grupo en Inglaterra han realizado una extraordinaria labor estudiando las cinasas y fosfatasas que participan en la regulación de la síntesis y degradación del glucógeno; sus estudios indican que existe una gran interacción en la propagación de las señales generadas por los diferentes sistemas de trausducción. En realidad, hemos separado los mecanismos de trausducción con fines de explicación, pero dentro de la célula hay una gran relación entre ellos. Esta interacción se lleva a cabo básicamente por fosforilación y defosforilación de las enzimas participantes.