VI.RECEPTORES CANAL Y RECEPTORES INTRACELULARES
ALGUNOS RECEPTORES funcionan de un modo muy distinto a los anteriormente descritos; tal es el caso de un tipo de receptor para la acetilcolina, llamado colinérgico nicotínico, el cual se encuentra ampliamente distribuido en el sistema nervioso y en la placa neuromuscular; un tipo de receptor para el GABA (ácido gamma amino butírico), y receptores para la glicina, entre otros. Estos receptores son proteínas integrales de membrana y están formados por varias subunidades. Son proteínas que funcionan como canal permitiendo el paso de iones a través de la membrana plasmática. Esta dualidad de funciones, receptor y canal, hace que, si los estudia un electrofisiólogo, los denomine canales activados por ligando y si los estudia un farmacólogo molecular o bioquímico diga que se trata de receptores canal. Ahora bien, independientemente de cómo los llamemos o de quién los estudie, son de enorme importancia.
Al activarse, su acción es formar un canal en su estructura que permite de ese modo el paso, a través de la membrana, de un ion como el sodio o el cloruro. Como es bien sabido, la célula gasta parte de su energía en mantener una distribución de iones, a un lado y otro de la membrana plasmática, alejada del equilibrio. La membrana es, por lo tanto, una barrera selectiva para los iones y mantiene cierto potencial electroquímico. Los receptores canal son bastante selectivos para los iones que dejan pasar. La apertura del canal tiende a colapsar los gradientes de concentración que existen para los iones que permean, lo que hace que la distribución de cargas en ambos lados de la membrana plasmática cambie drásticamente, es decir, que se desencadene una despolarización o hiperpolarización de la membrana. Esto conduce a que otros canales sensibles (es decir, que "sienten" ) al voltaje cambien su probabilidad de apertura (es decir, que estén más tiempo abiertos o cerrados, que en el estado basal), lo que puede traer consigo cambios importantes en la concentración de algunos iones moduladores del metabolismo, como el calcio. En el caso del receptor nicotínico, uno de los más estudiados y quizá el mejor conocido hasta ahora, se sabe que el canal a través del cual pasa el sodio es parte integral del receptor. Aquí, el agonista, al activar al receptor, produce un cambio en su estructura (cambio conformacional), o sea, una alteración tal en su forma que permite que se abra el canal y pase el sodio al interior de la célula. Esto desencadena respuestas como la contracción del músculo.
Pero, volvamos a la estructura de los receptores canal, particularmente del colinérgico nicotínico. Ésta se ha logrado esclarecer a través de la purificación de la proteína y del conocimiento y manipulación que se ha hecho de las secuencias genómicas que lo codifican. El receptor está formado por cinco subunidades: dos alfa, a las que se une el ligando para activarlo, y tres más llamadas beta, gamma y delta (figura 13). Estas subunidades se encuentran agrupadas formando una roseta con una depresión central que corresponde al canal. Es muy interesante que cada una de estas subunidades esté formada por una cadena amino terminal extracelular, cuatro segmentos transmembranales (llamados M1, M2, M3 y M4) unidos por asas intra y extracelulares, y una cadena extracelular carboxilo terminal. Aparentemente los segmentos transmembranales M2 de las cinco subunidades se encuentran en estrecha cercanía y son los que constituyen propiamente el canal. La especificidad para el paso de un ion y no de otros, depende aparentemente de los aminoácidos que constituyen este canal, pues cambiándolos por mutagénesis dirigida se ha logrado perturbar la selectividad iónica; en otras palabras, estas modificaciones pueden hacer que el receptor al activarse deje pasar un ion con carga negativa en lugar de uno con carga positiva.
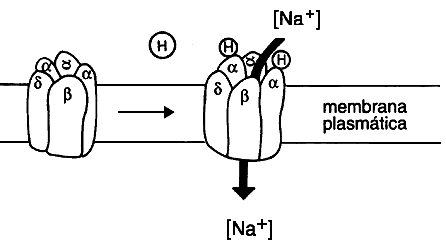
Figura 13. Representación de un receptor canal (colinérgico nicotínico)
con sus diferentes subunidades, la fijación de la hormona (acetil colina , H)
y la apertura del canal para la entrada de sodio.
Es importante señalar que las diferentes subunidades de los receptores canal para acetilcolina (nicotínico) para GABA (tipo A) y glicina tienen conservada esta estructura general con cuatro zonas transmembranales, lo que sugiere que todos estos receptores efectivamente forman parte de una superfamilia. Además existe una gran heterogeneidad entre sus subunidades, y por lo tanto el número de receptores relativamente diferentes para cada uno de estos neurotransmisores es amplísimo. Por otro lado, muchos de estos receptores tienen sitios accesorios, tanto intracelulares como extracelulares, para diversas sustancias que al ocupar estos sitios modulan la actividad del receptor-canal.
Algunas hormonas ejercen su acción sobre receptores intracelulares. Ello indica que, para estas hormonas, la membrana plasmática no es en realidad una barrera de permeabilidad. Dentro del grupo de hormonas que ejercen estas acciones tenemos a las hormonas tiroideas, al ácido retinoico y a los esteroides. Todos estos mensajeros tienen una gran importancia en los procesos de crecimiento y desarrollo, y en el mantenimiento de la homeostasis (equilibrio dinámico) de nuestro organismo.
Dada la facilidad con que atraviesan la membrana plasmática, a estas hormonas podemos asignarles dos caraterísticas fisicoquímicas fundamentales: I) son moléculas relativamente pequeñas, es decir, de bajo peso molecular, y 2) son, por lo menos, parcialmente hidrofóbicas, esto es, lipídicas. Estas características son necesarias si suponemos que las hormonas atraviesan la membrana por difusión, es decir, pasivamente. Se ha sugerido la existencia de sistemas activos para transportarlas al interior de la célula; pero la evidencia hasta el momento es escasa. En cualquier caso, los mensajeros que he mencionado cumplen con los requisitos fisicoquímicos descritos anteriormente. Las hormonas tiroideas son derivados de un aminoácido: la tirosina, cuyo peso molecular es relativamente bajo y cuya naturaleza es relativamente hidrofóbica. En el otro grupo, los esteroides son lípidos derivados del colesterol y por ende son tanto hidrofóbicos como de bajo peso molecular. Dentro de los esteroides hormonales tenemos a las hormonas sexuales masculinas (andrógenos) y femeninas (estrógenos y progestágenos), a los glucocorticoides (como el cortisol y la cortisona), a los mineralocorticoides (como la aldosterona), y a una vitamina-hormona, la vitamina D, de la que hablaremos más adelante.
Los receptores para las hormonas de este tipo son también proteínas. Por lo tanto, la información para la síntesis de estos receptores se encuentra también en nuestro ADN. Dicha información se expresa en algunas células y en otras no; dicho de otra manera, hay células que tienen estos receptores y otras que no. Estos receptores también muestran gran especificidad y reconocen las diferencias esteroquímicas que existen entre unas y otras hormonas. Esto no quiere decir que la especificidad sea absoluta. A semejanza de lo que mencioné en el capítulo de receptores de membrana, si aumentamos la concentración de un mensajero, éste llega a interactuar con otros receptores, además del propio, lo cual es un problema importante en la práctica médica cotidiana. Pondré un ejemplo: en el caso frecuente de la administración de cortisona por periodos prolongados, el médico debe controlar cuidadosamente la dosis, pues, además de los efectos propios de la hormona, se pueden presentar complicaciones indeseables que son fácilmente atribuibles a la interacción de la hormona con otros tipos de receptores; así podemos observar efectos similares a la aldosterona o aun de tipo testosterona. Estos receptores intracelulares, a diferencia de los que están localizados en la cara externa de la membrana plasmática, se encuentran libres en el citosol.
Dado que estas hormonas no interactúan con receptores de la membrana plasmática, la alteración de las proteínas de la membrana de la célula ("rasurar" con proteasas, por ejemplo) no hace desaparecer el efecto. Otra característica muy importante de la acción de estas hormonas de acción intracelular es el tiempo requerido para observar su acción. En general, las hormonas que actúan sobre receptores de membrana tienen algunas de sus acciones casi instantáneamente, esto es, ejercen sus efectos en segundos o minutos. En contraste, las acciones de las hormonas que actúan sobre receptores intracelulares tienen una latencia, es decir, un periodo de minutos a horas, durante el cual no observamos ningún efecto. Es necesario que aclare dos excepciones a esta generalización. Primera: muchas hormonas con receptores en membranas, como la insulina y muchos de los llamados factores de crecimiento celular además de muchas otras que se acoplan a proteínas G, tienen acciones inmediatas, pero también acciones más tardías con latencia de minutos a horas. Los mecanismos de tales acciones tardías se están estudiando activamente y tienen muchas semejanzas con las de los receptores intracelulares a que nos referiremos en unos momentos. Segunda: hormonas con receptores intracelulares que ejercen algunas de sus acciones en forma relativamente rápida. Ejemplos de ello serían la bien conocida acción antialérgica de la cortisona y algunos efectos sobre el transporte de iones de algunos esteroides. Desafortunadamente, poco se sabe aún cómo ocurren estas acciones rápidas; la evidencia con que se cuenta hasta el momento indica que no involucran los mismos procesos de las acciones "lentas". Se piensa, pues, que existen acciones en el ámbito de la membrana plasmática; incluso se ha dicho que "estabilizan" la membrana, lo cual es simplemente una más de las bellas palabras con que los científicos ocultamos nuestra ignorancia. Y que quizá describa algo de la fenomenología observada, pero desafortunadamente no nos habla de los mecanismos involucrados.
Los receptores intracelulares reconocen a la hormona, la fijan, y pasan a su configuración activa. Se ha demostrado que muchos de los receptores de este grupo, forman, en ausencia de la hormona, un gran complejo con algunas proteínas que facilitan el doblaje adecuado de las proteínas y que reciben el nombre de chaperoninas; algunas de ellas forman parte de las llamadas proteínas de "estrés" que se inducen rápidamente en las células cuando hay condiciones de emergencia. Una vez que llega la hormona y se fija al receptor formando un complejo hormona-receptor, las otras proteínas se van disociando aparentemente. El complejo hormona-receptor viaja al núcleo. El mecanismo del viaje es desconocido aún; se ha propuesto que algunas de las chaperoninas pudieran participan en dicha migración, pero la información con que contamos es escasa como para asegurarlo. Una vez en el núcleo, el complejo hormona-receptor se fija al material genético; dicha fijación parece no llevarse a cabo en cualquier lugar del genoma (ADN) sino en puntos concretos en los que interactúan con secuencias específicas. Estos sitios específicos corresponden frecuentemente a zonas de la región llamada promotora de los genes que se van a transcribir. El siguiente paso es la apertura de la doble hélice del ADN, para permitir que sea transcrito un mensajero específico; esto es, se hace una copia de ARN mensajero a partir del ADN (transcripción). Este ARN (ácido ribonucleico) mensajero lleva al citoplasma la información para la síntesis de una o varias proteínas; una vez allí, el mensaje es traducido en los ribosomas, formándose las nuevas proteínas. Pongamos un ejemplo: en el caso del oviducto de la gallina, la síntesis de la proteína del huevo, la ovoalbúmina, está controlada por las hormonas sexuales femeninas. En ausencia de hormonas, la síntesis de ovoalbúmina es baja. Ahora bien, si añadimos estrógenos a las células del oviducto, la síntesis de ovoalbúmina aumenta muchísimo. ¿Cómo ocurre esto? Como mencioné arriba (ilustrado en la figura 14), la hormona se fija al receptor citoplásmico, el cual, activado, viaja al núcleo junto con la hormona. Esto induce la síntesis de ARN mensajero que viaja al citoplasma y conduce a la síntesis de más ovoalbúmina. Es necesario mencionar que estos mensajeros no sólo incrementan la síntesis de algunas proteínas como hemos dicho; en algunos casos se produce un bloqueo de la transcripción de ciertos genes. Es decir, la acción sobre la transcripción puede ser de modulación positiva o negativa y una misma hormona puede tener efecto positivo sobre la transcripción de unos genes y negativo sobre otros.
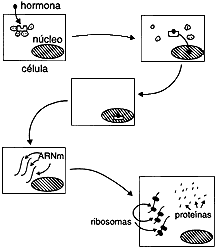
FIGURA 14. Mecanismo de acción de las hormonas con receptores citoplásmicos.
Resultará claro que la acción de estas hormonas no ocurre de inmediato; los procesos de transcripción (síntesis de ARN mensajero) y traducción (síntesis de proteínas) tardan varios minutos en las células de mamífero. Además, los efectos se observan por la acumulación de las proteínas cuya síntesis se induce, o por la disminución de la cantidad de aquéllas cuya síntesis se bloquea. Por otro lado, es importante mencionar que hay algunos agentes capaces de bloquear la transcripción o la síntesis de proteínas; estos agentes, por lo tanto, también bloquean todas las acciones de las hormonas que impliquen la biosíntesis de proteínas.
Quisiera acabar el capítulo hablando un poco de la interesante historia de la vitamina D. Las vitaminas son componentes esenciales de la dieta, de las cuales sólo necesitamos pequeñas cantidades. Para la sana existencia, son esenciales algunos compuestos que somos incapaces de sintetizar. Dentro de estos componentes esenciales están: algunos aminoácidos y lípidos y las vitaminas. Me referiré sólo a estas últimas. Aunque muchas de estas sustancias no son aminas, se ha conservado el nombre por tradición. En general, requerimos una pequeñísima cantidad de ellas, porque funcionan como parte de los sistemas enzimáticos (como coenzimas), así que son reutilizados en múltiples ocasiones.
La vitamina D no es una amina, se trata de un derivado del colesterol. La requerimos porque no podemos propiamente sintetizarla; pero, en principio, no es necesario ingerirla. ¿Cómo está eso? Parece una contradicción, pero no lo es. Basta la exposición a los rayos solares para que una fracción del colesterol de la piel se convierta en vitamina D. Se puede administrar por vía oral, pero es mucho más agradable y barato darse simplemente una prudente asoleada.
La carencia de vitamina D en los niños da lugar al raquitismo. Esta enfermedad era relativamente rara en nuestro país dada su localización geográfica. Sin embargo, los cambios en nuestra forma de vida van haciendo que frecuentemente se vean más casos. Cada día son más las madres trabajadoras y las "muy ocupadas" que no cuentan con 15 minutos para sacar a sus hijos a que les dé un poco la luz del Sol. Los padres tampoco parecen muy dispuestos a colaborar en ello. Otro factor, posiblemente importante en las ciudades, es la contaminación, que filtra la iluminación que recibimos. El raquitismo da lugar a serias alteraciones en el crecimiento de los huesos.
La vitamina D es transformada tanto en el hígado como en el riñón para convertirla propiamente en la hormona: el dihidrocolecalciferol. Esto coloca, pues, a la vitamina D en el papel de prohormona (precursor de la hormona). Muchas de las aportaciones más significativas sobre esta vitamina han sido realizadas por el doctor Héctor F. de Luca. El esclarecimiento de la síntesis del dihidrocolecalciferol ha sido de gran importancia, no sólo desde el punto de vista científico ya que tiene claros matices aplicativos. Pondré un ejemplo: se había observado en Inglaterra que en algunos grupos de pacientes de origen oriental existían niños que desarrollaban raquitismo a pesar de recibir el Sol e ingerir suficientes cantidades de vitamina D. El tratamiento que recomendaban los médicos era aumentar fuertemente las dosis, se observaba alguna mejoría pero dicha terapéutica era en general insatisfactoria. Lo que parece ocurrir es que estos pacientes tienen bloqueada o disminuida la biosíntesis de la hormona. Con el advenimiento de los nuevos conocimientos es posible determinar cuál es el problema en cada paciente y, lo que es más importante, ofrecerle una esperanza con un tratamiento más racional.
Decía, entonces, que la prohormona (vitamina D) se transforma en hormona (dihidrocolecalciferol) y se almacena para ser secretada cuando se requiera. Pudiera decirse que dentro de las principales acciones de la hormona tenemos: 1) aumentar la absorción de calcio, y 2) favorecer su fijación en el hueso. Suena lógico, por lo tanto, que sea el calcio en sangre uno de los reguladores de la secreción de esta hormona; esto es, cuando la concentración de calcio en sangre baja, se incrementa la secreción de dihidrocolecalciferol. Una vez liberada, la hormona viaja al intestino y allí activa a sus receptores intracelulares específicos; viaja al núcleo y favorece la transcripción de ARN mensajero. Este mensajero es traducido, y da lugar a la síntesis de una proteína encargada de la captación del calcio. Es en esta forma como la vitamina D favorece la absorción intestinal de calcio.