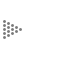II. ¿CÓMO DICES?
| Quedéme y olvidéme, El rostro incliné sobre el amado, Cesó todo, y dejéme, dejando mi cuidado entre las azucenas olvidado. |
SAN JUAN DE LA CRUZ. |
HAY MUCHAS maneras de entendernos. Naturalmente, cuando pensamos
en qué piensa, siente o imagina otra persona, lo primero que nos viene a la
mente es el lenguaje. Si quiero saber cómo está alguien a quien conozco o quiero
conocer, o acercarme a alguien que me interesa, me aproximo a él o ella y le
hablo. Pero si reflexionamos un poco, inmediatamente reconocemos que de ninguna
manera es el lenguaje la única manera de comunicarnos con otra persona ni de
saber algo de ella. La fotografía de la niña sonriente, de la cultura totonaca
del Golfo de México ( Figura 7.), es un excelente ejemplo de cómo podemos comunicarnos
sin hablar. La sonrisa es, en efecto, uno de los medios más eficaces para lograrlo,
tanto que casi inconscientemente (y a veces muy conscientemente) la usamos con
mucha frecuencia. Cuando lo hacemos, si hay reciprocidad y obtenemos otra sonrisa
como respuesta, sabemos que algo existe entre la persona que nos respondió y
nosotros, así sea ese algo meramente momentáneo. Pero muchas veces ni siquiera
es necesaria una sonrisa ni un gesto: basta una mirada en que dos pares de ojos
se encuentran por un segundo para que ambos sepan que hay algo en común entre
los poseedores de los ojos que se miraron. Ciertamente, si entre las personas
existen ciertas afinidades o cosas en común, esto se dará no sólo momentáneamente
sino de manera continua, y mientras más cerca estemos de alguien, más serán
los mecanismos de comunicación no verbales que constantemente se pongan en juego
para "hablarse". No es raro, inclusive, entre parejas que se han identificado
y convivido durante cierto tiempo, que se lleguen a "adivinar" los pensamientos,
lo cual no es sino una manifestación de la profunda identificación y el claro
conocimiento mutuo, lo cual permite conocer el significado de pequeños signos
como miradas, movimientos, gestos, sonrisas, exclamaciones o actitudes corporales.

Figura 7. Niña Sonriente. Escultura totonaca del Golfo de México.
Existen, pues, muchas maneras de saber qué dice otra persona. No olvidemos, sin embargo, que cada una de esas maneras, como cualquier otra manifestación del modo de ser o de pensar, implica necesariamente el funcionamiento del cerebro, y que éste a su vez requiere de la continua actividad de las neuronas y de la constante comunicación entre ellas.
Recordemos ahora que las neuronas, como cualquier célula viviente, están constituidas por átomos organizados en moléculas, y que las moléculas tienen también una peculiar organización, muy precisa, de la cual depende que la célula pueda vivir llevando a cabo todas las funciones. Con esta idea en mente, no debe extrañarnos que los mecanismos de comunicación entre las neuronas sean moleculares, es decir, realizados mediante ciertas moléculas específicamente organizadas para permitir y facilitar dicha comunicación interneuronal. Por esta razón, en este capítulo revisaremos los aspectos generales más importantes de la comunicación entre moléculas, empezando por el átomo mismo, e iremos paulatinamente aumentando el grado de complejidad de tal comunicación intermolecular, para llegar finalmente a la comunicación del cerebro con el resto del organismo, como órgano maestro que lo coordina, organiza y controla. Empecemos, pues, por el principio.
Prácticamente todo lo que existe está hecho de pequeñas partes que se ensamblan entre sí, de tal manera que forman otras unidades más grandes, pero al mismo tiempo más complejas. Éstas a su vez se organizan con otras similares para dar origen a nuevas formas o estructuras, aún más grandes y más complejas, y así progresivamente. Sin embargo, lo que más nos interesa para acercarnos poco a poco a los mecanismos de comunicación interneuronal no es que al unirse las unidades se formen estructuras de mayor tamaño, sino de mayor complejidad. Veamos por qué.
Entre los seres vivos esta unión que resulta en una mayor complejidad es una característica, como veremos, muy interesante. También se presenta en el mundo orgánico de los seres no vivientes.
Imaginemos inicialmente al átomo, cualquier átomo: está compuesto por su parte central, el núcleo, y por una nube de electrones que se mueven en la periferia del núcleo, siguiendo determinado patrón que les da su energía. Un átomo es, así, un conjunto de partículas de distinta naturaleza; aquéllas que forman el núcleo, notablemente los protones que poseen una carga eléctrica positiva, y los electrones, que son mucho más pequeños y que tienen carga eléctrica negativa. La estructura del átomo incluye ambos tipos de partículas, pero éstas en sí mismas distan mucho de ser un átomo. Así pues, de la atracción que existe entre los protones con carga positiva y los electrones con carga negativa (hay el mismo número de protones que de electrones), resulta la estructura del átomo, ya que las cargas opuestas se atraen. (Figura 8.)
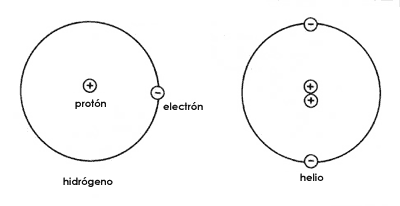
Figura 8. Los dos átomos más sencillos : a la izquierda el de hidrógeno, que
tiene un solo protón con carga positiva en su núcleo, y un solo electrón en
su periferia; a la derecha, un átomo de helio, que posee dos protones en su
núcleo y dos electrones en la periferia. La atracción o afinidad entre la carga
positiva de los protones y la negativa de los electrones mantiene la estructura
de los átomos.
Es precisamente debido a esta composición del átomo que éste puede unirse con otros átomos para formar moléculas. Un átomo completo es capaz de ganar un electrón, por su tendencia a formar pares de ellos hasta completar cierto número, y cuando esto ocurre resulta que ha adquirido una carga negativa, pues le falta un protón para compensar la carga negativa adicional. De manera similar, un átomo puede perder un electrón y en este caso le sobrará un protón que no tiene una carga negativa compensatoria y por lo tanto ese átomo tendrá una carga positiva. Cuando esto sucede, puede ocurrir que un átomo con carga negativa se relacione con otro con carga positiva, de manera que se constituye una molécula formada por dos átomos atraídos mutuamente por tener cargas contrarias. Las sales, por ejemplo la sal común de mesa (el cloruro de sodio), son moléculas formadas de esta manera; además las moléculas de este tipo pueden formar cristales regulares precisamente por tener esta propiedad de que sus átomos están cargados y los de carga positiva atraen a los de carga negativa (Figura 9.)
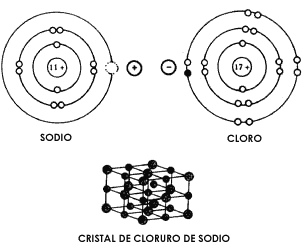
Figura 9. Afinidad por atracción de cargas opuestas entre un átomo de sodio que ha perdido un electrón (señalado con el círculo de puntos) y por lo tanto quedó con un protón cuya carga no está neutralizada, y uno de cloro que ha ganado un electrón (señalado con un círculo negro) y por lo tanto quedó con una gran carga negativa no compensada por ningún protón. Esta afinidad por cargas entre el sodio y el cloro hace que se unan para constituir la sal común o cloruro de sodio. Cuando la sal se encuentra en estado sólido forma cristales, cuya estructura se esquematiza en la parte inferior; los átomos de cloro son las esferas más grandes.
Existe otra manera mediante la cual los átomos se pueden unir entre sí para
formar moléculas. En lugar de que un átomo pierda y otro gane un electrón, lo
que sucede es que ambos átomos simultáneamente ceden y reciben un electrón,
con el resultado de que dos electrones son compartidos por los dos átomos, los
cuales quedan así enlazados. Este es el caso del agua, en que cada uno de sus
dos átomos de hidrógeno comparte un electrón con el átomo de oxígeno, formándose
la molécula H20, como se ilustra en la Figura 10.
Figura 10. Afinidad por compartición de electrones. En la molécula de agua (H2O)
, el oxígeno comparte dos de sus electrones de su última órbita (indicados con
un asterisco) con sendos átomos de hidrógeno, cada uno de los cuales, a su vez,
comparte con el de oxígeno su único electrón, señalado con un círculo negro.
El resultado de compartir electrones es que el átomo de oxígeno completa 8 electrones
en su última órbita, y cada uno de los átomos de hidrógeno completa 2. Como
ésta es la forma más estable de los átomos mencionados, se estabiliza la molécula
de agua en la forma H2O.
MACROMOLÉCULAS, AFINIDAD Y ESPECIFICIDAD
Existe en la naturaleza un átomo muy especial, que sólo puede unirse a otros
mediante el mecanismo de compartir electrones, y además lo puede hacer con cuatro
átomos distintos. Este interesante átomo es el carbono, el cual, debido a esa
propiedad, es un átomo clave en todas las moléculas que constituyen a los seres
vivos. (Figura 11.) Cuando el carbono se une de cierta forma peculiar con otros
átomos de carbono, de hidrógeno, de oxígeno y de nitrógeno, forma una enorme
variedad de moléculas, tanto pequeñas como muy grandes, que son parte estructural
y funcional de las células vivientes. Algunas de estas moléculas están formadas
por relativamente pocos átomos, 12, 20 ó 40, mientras que otras son gigantescas,
ya que tienen muchos cientos o hasta miles de átomos.
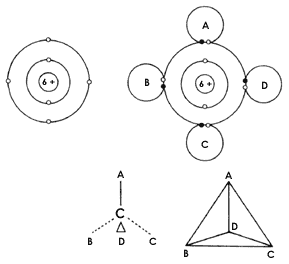
Figura 11. El átomo de carbono. Como se muestra en la parte superior izquierda,
este átomo tiene 4 electrones en su última órbita, por lo que tiende a completar
8 mediante el mecanismo de compartir que se muestra en la figura 10 para el
caso del agua. Así, el carbono puede unirse con varios átomos iguales o bien
distintos, de la manera que señala en la parte superior derecha (sólo se indica
aquí la última órbita y el electrón que comparten, en negro, los átomos A, B,
C y D). En los esquemas de la parte inferior se observa que las uniones que
cada átomo de carbono establece, se orienta en direcciones distintas, como si
el carbono estuviera en el centro de un prisma triangular y los cuatro átomos
con los que se une ocuparan los vértices del prisma. Esta propiedad del carbono
es muy interesante, pues da lugar a que las moléculas muy grandes formadas por
él, adquieran formas tridimensionales distintas.
Entre estas moléculas muy grandes, que reciben el nombre genérico de macromoléculas, existen unas que por sus muy especiales propiedades constituyen parte esencial de los mecanismos que mantienen las funciones de las células vivas. Estas moléculas son las proteínas, las cuales poseen una estructura de tal naturaleza, que permite la existencia de un mecanismo de comunicación nunca visto antes de su aparición sobre el planeta: la comunicación por medio del reconocimiento de la forma específica de cada molécula, de manera que las diferentes proteínas pueden identificar a otras porque su forma tridimensional en el espacio les permite acomodarse perfectamente una en la otra, en una especie de acoplamiento por similitud, o mejor dicho, por afinidad entre sus estructuras. (Figura 12.) Es decir, una molécula de determinada proteína será capaz de reconocer, y de combinarse con ella, sólo con otra cierta proteína, porque sus estructuras son afines. De hecho, es interesante que lo que hemos dicho antes respecto a la comunicación entre los átomos para formar moléculas encaja perfectamente en esta idea de la comunicación por mecanismos de afinidad. Por ejemplo, la carga opuesta del electrón con respecto al protón, sin duda indica un tipo de afinidad, definido precisamente como esa diferencia de carga que las hace opuestas y al mismo tiempo, atraerse mutuamente. De manera similar, entre los átomos la atracción para complementarse en electrones, representa también una afinidad, la cual les permite unirse para formar las moléculas. En el caso especial del átomo de carbono, la afinidad es tan particular que este átomo es capaz de compartir cuatro de sus electrones, cada uno de ellos con un átomo diferente, o bien con átomos iguales. Así pues, y volviendo a las proteínas, estas interesantísimas moléculas iniciaron una forma de reconocimiento entre ellas con características muy peculiares, ya que su forma particular en el espacio, en tres dimensiones y no solamente en dos, les permite reconocerse y asociarse. Esto es lo que está ejemplificado en la Figura 13, que muestra dos de estas grandes moléculas en el espacio reconociéndose mutuamente.
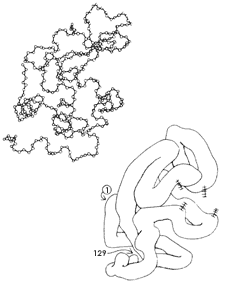
Figura 12. Estructura tridimensional de una macromolécula, una proteína. Cada
uno de los pequeños círculos en el esquema superior es un conjunto de átomos,
fundamentalmente de carbono, oxígeno, hidrógeno y nitrógeno, que constituyen
unidades pequeñas. Como se explica en el texto y en la Figura 14, estas unidades
tienen "brazos" laterales que se atraen entre sí por sus cargas, como los átomos
de la figura 9, lo cual origina los dobleces y da lugar a la forma particular
en el espacio, que se ilustra con más claridad en el dibujo inferior. Los números
1 y 129 indican, respectivamente, la primera y la última unidad, para dar mejor
idea de la disposición tridimensional.
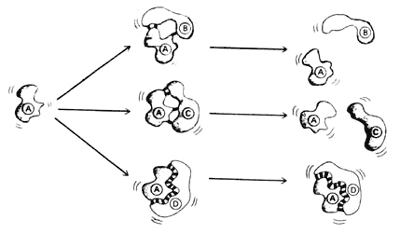
Figura 13. Reconocimiento específico de dos macromoléculas entre sí. La molécula
A puede asociarse momentáneamente con las moléculas B y C porque
puede establecerse una atracción parcial por cargas opuestas en ciertas partes
de las moléculas. Sin embargo, la estructura tridimensional de cada una de ellas
no es lo suficientemente "complementaria" como para permitir que tal asociación
sea completa y permanente. En cambio, entre las moléculas A y D,
su forma tridimensional les permite reconocerse con gran afinidad, por lo cual
permanecen unidas, como se muestra en la parte inferior.
Esta forma novedosa de poder reconocerse y acoplarse significa un gran paso en los mecanismos de afinidad entre las moléculas, debido fundamentalmente a una poderosa propiedad, que tendrá consecuencias de enorme trascendencia para las funciones celulares y particularmente para la función de comunicación entre las células nerviosas, como lo analizaremos en el Capítulo IV. La propiedad a que nos referimos es la especificidad, que puede definirse como la capacidad de las proteínas de reconocer y asociarse solamente con determinadas proteínas. Así, una molécula proteica reconocerá específicamente a otra molécula proteica, pero sólo a ella y no a ninguna otra, por más cerca que se encuentre. En otras palabras, esta propiedad de la especificidad impedirá que haya errores en el reconocimiento, ya que sólo aquellas que estén hechas "una para la otra" se podrán reconocer y asociarse, dejando de lado todas las demás.
Vale la pena detenernos un poco en este punto para analizar a qué se debe esta especificidad, ya que es una propiedad de extraordinaria importancia en la comunicación biológica.
Una primera aproximación sería pensar que las proteínas son tan grandes que su mismo tamaño sirve como lenguaje de comunicación entre ellas: las de tamaño X van a reconocer a las de tamaño X, las de tamaño Y a las de tamaño Y, etc. Sin embargo, sabemos que hay en las células muchas moléculas muy grandes que sin embargo son incapaces de reconocer a otras moléculas similares en tamaño. Es más, son incapaces de reconocer a ninguna otra molécula, sea del tamaño que fuere. Por ejemplo, las moléculas del almidón en las plantas, que tienen su equivalente en los animales, son mucho más grandes que muchas proteínas, y sin embargo no son capaces de reconocer a otras moléculas. Por otra parte, un reconocimiento por el tamaño no podría ser muy específico, pues seguramente habría muchas moléculas diferentes pero de tamaño similar.
Si no es el tamaño, ¿podría ser la carga eléctrica, como entre los átomos? claramente podemos ver que difícilmente podría ser la carga, ya que no podría explicarse entonces la especificidad, pues cualquier molécula de carga opuesta sería reconocida.
FORMAS TRIDIMENSIONALES: INFORMACIÓN
¿Qué otra característica podría explicar esa afinidad específica por otra molécula? La forma. En efecto, ya hemos dicho que su estructura tridimensional en el espacio es lo que concede a las proteínas esa capacidad maravillosa de reconocer específicamente sólo a ciertas otras moléculas. Pero esto nos lleva a hacernos una pregunta clave: ¿de qué depende la forma tridimensional de las proteínas? La respuesta es muy simple, aunque para explicarla en detalle podríamos ocupar todo el resto de este libro. Esa respuesta simple es que la forma en el espacio de las proteínas está determinada por su composición, es decir por las pequeñas unidades de que está compuesta. En efecto, como esbozamos más arriba, una proteína está hecha de unidades, es decir de moléculas más pequeñas, las cuales se unen entre sí para formar cadenas. Sin embargo, la estructura química de las unidades y el modo como se unen permite que queden hacia los lados de la cadena pequeños brazos formados por cadenitas cortas de átomos, y estos brazos no son iguales, sino que su tamaño y propiedades dependen a su vez del tipo de unidad pequeña a que pertenecen. Estos brazos son capaces de atraerse entre sí o de repelerse, siguiendo las reglas de atracción de cargas o de compartir electrones que ya mencionamos, pero como cada uno de ellos tiene su cabeza unida a la de las otras unidades que componen la proteína, el resultado es que, como se muestra en la Figura 14, la cadena de unidades que constituye la proteína se dobla sobre sí misma en varias direcciones. Precisamente es la orientación espacial de todos estos dobleces lo que hace que la proteína adquiera una forma definida tridimensional en el espacio.
Ahora bien, ¿qué pasaría si la cadena de unidades de una proteína estuviera
formada por unidades iguales entre sí? Es evidente que en esas condiciones la
molécula no podría tener una forma tan definida, pues como todos los brazos
laterales de esas unidades serían iguales, no tendrían manera de atraerse o
repelerse y, por consiguiente, habría muy pocas posibilidades de que la molécula
se doblara sobre sí misma para que adquiera su peculiar estructura tridimensional.
En cambio, si las unidades son diferentes entre sí, se facilitaría la posibilidad
de los dobleces. Y, en efecto, las proteínas están compuestas por 20 unidades
diferentes, cada una de las cuales puede repetirse muchas veces a lo largo de
la cadena, lo que da muchas posibilidades distintas de brazos que pueden atraerse
y por lo tanto de formas que pueden tomarse.
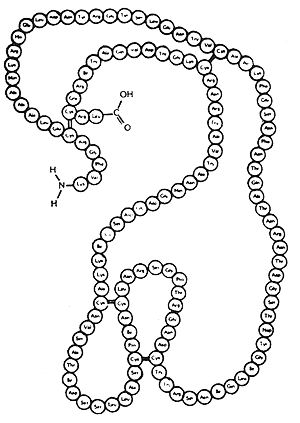
Figura 14. Ésta es la misma molécula de la Figura 12 pero con su estructura tridimensional simplificada para dejar ver los "brazos" laterales más importantes de algunas de las unidades que la componen y, cómo la unión de estos brazos entre sí, causa que la molécula se doble varias veces sobre sí misma y de esta manera adquiera su forma particular en tres dimensiones. Las letras en el interior de las unidades indican los nombres de las 20 unidades diferentes que forman proteínas, aunque evidentemente se repiten muchísimas veces casi todas ellas para dar lugar al enorme tamaño de las proteínas.
Examinemos este punto con más detenimiento estudiando la figura: Imaginemos por ejemplo que el brazo A atrae al brazo B. Es claro que, dependiendo de lo cerca o lejos que estén uno de otro a lo largo de la cadena, el doblez de ésta originará un asa de distinto tamaño: si A y B están muy cerca uno de otro, el asa que se formará será muy pequeña y, si están muy lejos será muy grande. Así, podemos concluir que, la forma de las proteínas en el espacio depende del orden en que estén colocadas las unidades que la componen, de lo que dependerá naturalmente la posibilidad de que sus correspondientes brazos interactúen para hacer que la molécula se doble. Y de esta conclusión extraemos otra que es de capital importancia para entender un poco la función comunicadora de las proteínas y de otras macromoléculas dentro y fuera de las células, la cual revisaremos enseguida. Esta conclusión es la siguiente: las proteínas son moléculas que llevan en su estructura una información.
¿Por qué decimos que esta conclusión es de capital importancia? Porque, además de tener enormes implicaciones para las funciones celulares, si hablamos de información uno debería esperar que las proteínas sean capaces de transmitir esa información. Veamos estos aspectos de función-información con cierto cuidado. ¿Cómo puede una molécula transmitir la información que contiene en su estructura? La respuesta a esta pregunta es doble. Por un lado, está la posibilidad de que literalmente se transmita la información de una proteína a otra, en el sentido de que se pueda copiar exactamente su composición, generándose una nueva molécula idéntica a la proteína de la cual se copió la información. Por otra parte, una segunda manera de transmitir la información sería la de poder "decirle" a la célula que realice o no determinada función o lleve a cabo determinado proceso. Si bien en este caso no se trata de una transmisión de la información en el sentido estricto del término, pues no se está copiando dicha información, sí es evidente que se trata de una transmisión del contenido de esa información en cuanto a la función que la célula debe realizar en un momento dado.
No podemos aquí revisar los interesantísimos mecanismos moleculares de la transmisión de la información en el sentido de copiarla en la generación de otras moléculas, pues esto se sale del tema y de los objetivos del libro que pretende concentrarse en las células del cerebro. Baste decir que dichos mecanismos involucran a otro tipo de macromoléculas que también poseen información en su estructura, llamadas los ácidos nucleicos, y que es gracias a estos mecanismos que, mediante el proceso de reproducción, las células hijas tienen las mismas proteínas que la célula madre y por lo tanto pueden realizar las mismas funciones.
A cambio de no revisar estos aspectos, veremos con algún detalle el otro modo que tienen las proteínas de transmitir información, mediante el dictado de qué es lo que la molécula debe hacer en un momento dado. Para revisar este aspecto, que si es esencial para el tema que nos ocupa, debemos hacer algunas consideraciones respecto al funcionamiento celular en general y posteriormente tocaremos ciertas funciones celulares específicas que nos interesan más en particular, para saber cómo se comunican las células nerviosas.
INFORMACIÓN MOLECULAR Y REACCIONES QUÍMICAS EN LA CÉLULA
Toda célula viva es un pequeñísimo laboratorio —pequeño en su tamaño pero extraordinariamente grande y variado en sus capacidades—, en el que se llevan a cabo una multitud de reacciones químicas de las que depende de manera absoluta el que la célula funcione adecuadamente. Es mediante estas reacciones químicas que la célula puede obtener energía de los alimentos, puede reproducirse y puede llevar a cabo la o las funciones especificas a que está destinada, como la contracción muscular, la secreción de líquidos y sustancias como hormonas, bilis, jugo gástrico o saliva, y el envío de señales hacia otras regiones. Ahora bien, todas estas reacciones químicas que deben ocurrir en el seno de la célula tienen que llevarse a cabo a una enorme velocidad, pues de otro modo no servirían para los fines que la célula requiere. En efecto, el laboratorio que es cada célula es incomparablemente más rápido que cualquiera de los laboratorios químicos que el hombre ha construido para sintetizar la multitud de compuestos que utilizamos en nuestra vida diaria, desde las medicinas, hasta los productos plásticos y los detergentes que empleamos constantemente.
¿Cómo es esto posible? ¿Cómo es que las células son capaces de realizar tantas reacciones químicas a tan alta velocidad? La respuesta es: las proteínas.
Aprovechando su increíble capacidad para reconocer moléculas específicamente, las proteínas pueden actuar sobre dichas moléculas para acelerar cientos o aun miles de veces la velocidad de sus reacciones de interconversión, mediante el proceso que los químicos conocen como catálisis. (Figura 15.) Además, y esto es extraordinariamente útil para la célula, lo hace en forma específica, es decir, cada proteína que cataliza una reacción lo hace sólo sobre esa reacción y ninguna otra, de nuevo haciendo uso de su formidable capacidad de reconocimiento específico de las moléculas involucradas en la reacción. Es por esto que puede decirse, sin temor a exagerar, que cada proteína realiza esta acción de acelerar una reacción específica en particular y que, al hacerlo, permite que funcionen los mecanismos vitales para la sobrevivencia y la función celulares, además de que está haciendo uso de la información que posee en su forma tridimensional; sin esta propiedad única de cada molécula proteica, no podría ni reconocer a la molécula que se debe transformar en otra, ni ser específica para ella, tampoco podría acelerar la velocidad de la reacción necesaria para que la célula funcione normalmente.
El atento lector que haya llegado hasta este punto estará sin duda pensando
que debe existir una cantidad enorme de proteínas distintas una de otra, si
es cierto que hay muchísimas reacciones químicas y que cada una de ellas es
acelerada en forma específica por una proteína diferente. En efecto, éste es
el caso. Una sola célula contiene miles de proteínas diferentes, y por supuesto
el número de moléculas de cada proteína es también muy grande.
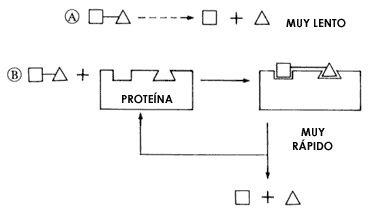
Figura 15. Muchas proteínas, por su particular forma tridimensional, pueden
reconocer específicamente a otras moléculas, también muy grandes o mucho más
pequeñas, y convertirlas en otras a una enorme velocidad, mediante lo que se
llama su actividad catalizadora. En el esquema A, representa al rompimiento
de la molécula formada por el triángulo y el cuadrado en cada uno de sus componentes,
es una reacción que ocurriría tan lentamente en ausencia de una proteína catalizadora,
que es posible decir que no ocurriría. En cambio, B señala que ante la
presencia de la proteína catalizadora, la molécula es acomodada en un sitio
específico y, como resultado, se facilita cientos de veces el proceso del rompimiento,
por lo que la reacción se acelera esos mismos cientos de veces; además la proteína
queda libre para repetir el proceso.
INFORMACIÓN MOLECULAR Y MENSAJES:TRANSMISORES
Pero lo que acabamos de revisar no es el único modo que tienen las proteínas de ejercer su función transmisora de información (estoy suponiendo que he sido lo suficientemente convincente para que el lector acepte que la acción catalizadora de reacciones de las proteínas es un modo de transmisión de la información contenida en su estructura). Muchas células utilizan las proteínas para otras funciones, de una manera que se acerca todavía más a una verdadera transmisión de la información: las secretan, las exportan para que actúen en otro lado, sobre otras moléculas o sobre otras células. Un ejemplo muy claro de esta transmisión mediante exportación lo tenemos en los procesos digestivos. Cada una de las células de las glándulas que participan en la digestión, desde las salivales que secretan saliva hasta las de la pared del estómago que hacen el jugo gástrico, secretan proteínas que van disueltas en su respectivo líquido. Estas proteínas son capaces de reconocer ciertas moléculas que son ingeridas en los alimentos y que deben ser convertidas en otras más simples, más pequeñas, que puedan ser absorbidas desde el intestino y aprovechadas por todas las células del organismo. Así, es posible afirmar que las células secretoras de estas proteínas digestivas transmiten la información de ellas hacia la boca o hacia el tubo digestivo. Este es un hermoso ejemplo de especificidad de la información de una proteína trabajando en otro sitio diferente a la propia célula que la produce.
No faltará quien, a pesar de todo lo que hemos dicho hasta ahora, tenga todavía dudas de hasta qué punto realmente las proteínas, mediante esa extraordinaria capacidad para reconocer moléculas e interactuar con ellas, pueden transmitir información en el sentido más estricto de enviar mensajes de la célula que la produce a otra que la recibe. Pues bien, como veremos enseguida, muchas proteínas, sin tener una acción catalizadora de reacciones químicas, se comportan precisamente de este modo, funcionando como mensajeros entre células. El ejemplo más demostrativo es el de las hormonas de naturaleza proteica. Analizaremos ahora con algún detalle esta función.
Una hormona es una sustancia que es sintetizada por las células de las llamadas
glándulas endocrinas, pero que no ejerce ninguna acción sobre la misma célula
que la produce, sino que ésta la secreta hacía la sangre para que sea transportada
a otras células, sobre las que sí actuará. Según el tipo de la hormona de que
se trate será la clase de efecto que produzca sobre la célula sobre la que actúa.
Sin embargo, en todos los casos debe haber, una vez más, un reconocimiento de
esa célula hacia la molécula hormonal que le está llegando a través de la sangre.
(Figura 16.) Para que tal reconocimiento ocurra —ya no debe extrañar al
lector—, es necesario que la célula que recibe la acción de la hormona
tenga en su membrana externa, que es lo primero con lo que la hormona se va
a encontrar, otra proteína con la afinidad suficiente para identificar a la
hormona. Gracias a esta identificación la célula podrá interaccionar específicamente
con la hormona y como resultado de esta interacción se ejercerán sus efectos.
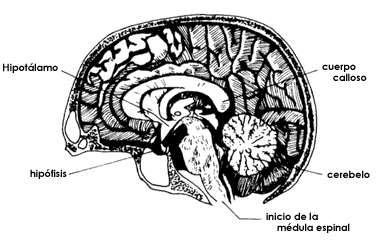
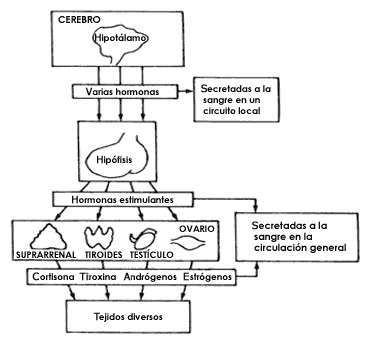
Figura 16. El cerebro es también el controlador de la actividad de las glándulas
de secreción interna que fabrican las hormonas. El dibujo superior, parecido
al de la Figura 6, muestra la cara interna de un hemisferio cerebral y el corte
del cuerpo calloso, del tallo cerebral, del hipotálamo y de la hipófisis. Estas
dos últimas estructuras son las encargadas de coordinar y regular la secreción
de las hormonas, las cuales funcionan como mensajeros químicos que van a actuar
sobre los distintos tejidos del organismo, como se señala en la parte inferior
del dibujo. Así, el hipotálamo regula a la hipófisis mediante ciertas hormonas
que se secretan en un circuito local de circulación sanguínea y, a su vez, la
hipófisis secreta otras hormonas a la circulación general que van estimular
al resto del conjunto de glándulas endocrinas como suprarrenales, tiroides,
glándulas sexuales, etc. Finalmente, las hormonas fabricadas y secretadas a
la sangre por estas glándulas, actúan sobre los distintos órganos.
En este punto es necesario hacer notar que por primera vez estamos hablando
del reconocimiento de una molécula que está fuera de la célula, soluble en el
agua (en éste caso en el agua de la sangre), por otra proteína que no está soluble
sino que se encuentra embebida en el seno de la membrana de una célula. Es importante
señalar, sin embargo, que aunque estas proteínas están embebidas en la membrana,
es claro que una porción de ellas, particularmente aquella porción involucrada
en el reconocimiento de la hormona, debe estar expuesta hacia el exterior de
la membrana, de modo que pueda "ver" a la molécula que se aproxima. (Figura
17.) A estas moléculas proteicas que se encuentran en la membrana de las células,
que pueden reconocer a otras moléculas e interactuar con ellas para que se produzca
una determinada respuesta célular, las llamamos moléculas receptoras,
o simplemente receptores. Y henos aquí ya hablando de moléculas transmisoras
que llevan un mensaje o simplemente transmisores, y de moléculas que
reciben o aceptan el mensaje, inicialmente mediante el reconocimiento del transmisor.
Podemos por lo tanto concluir, una vez más, que como este reconocimiento depende
de la forma de la molécula, de hecho lo que se está llevando a cabo es una transmisión
de la información contenida en la molécula transmisora.
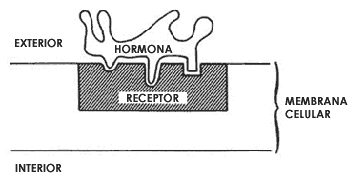
Figura 17. Reconocimiento de una hormona por una molécula receptora embebida
en el seno de la membrana celular. La estructura tridimensional de las dos moléculas
es tal que permite su reconocimiento específico, y por lo tanto su unión. El
receptor tiene una porción expuesta al exterior de la membrana celular por medio
de la cual reconoce a la hormona. El resultado de la interacción de la hormona
con su receptor es que otras proteínas de la membrana celular y/o del interior
de la célula se modifican y en consecuencia la célula cambia una o varias de
sus funciones, por ejemplo la velocidad de ciertas reacciones químicas importantes
para su nutrición. La hormona actúa así como un verdadero mensajero químico
cuya información está en su forma tridimensional.
Pero es tiempo de recapitular para retomar el hilo de nuestra discusión anterior, que se refería a cómo se van complicando los mecanismos mediante los cuales se puede establecer comunicación entre componentes diversos o entidades distintas. Así, de la afinidad por atracción de cargas entre el protón y el electrón o entre átomos cargados positiva o negativamente, pasamos a considerar la afinidad o comunicación por compartir electrones, mecanismos en el que los átomos de carbono resultan maestros. Dando un enorme salto, enseguida revisamos el caso de las moléculas muy grandes compuestas por unidades más pequeñas, y vimos que del orden en que estas unidades estén colocadas en las macromoléculas depende su forma tridimensional, y cómo de esta estructura tridimensional depende a su vez la capacidad de reconocer por afinidad a otras moléculas. De aquí concluimos que estas macromoléculas tienen información, la cual les sirve para reconocer otras moléculas pero también les permite transmitirla de diferentes maneras, sea en el sentido estricto del término (copiar la estructura molecular) o bien, porque al tener esa información son capaces de que la célula haga lo que tiene que hacer para seguir viviendo. Finalmente, llegamos también a revisar que muchas de esas moléculas son capaces de transmitir información funcional a otras células, actuando como mensajeros o como receptores.
Para seguir, pues, con el hilo de la creciente complejidad en los mecanismos de comunicación (o de afinidad, que en mucho hemos identificado el significado de estos dos términos), resulta evidente que en la discusión anterior hemos dado otro paso enorme, pues casi sin darnos cuenta pasamos de la comunicación intermolecular a la comunicación entre las células. Este paso es gigantesco, pues implica que ya estamos hablando de organismos formados por muchas células y que además se han ido especializando en llevar a cabo ciertas funciones que las demás no pueden hacer, por ejemplo, secretar sustancias para que actúen a distancia sobre otras células, y que éstas a su vez poseen moléculas receptoras para reconocer a las transmisoras.
De lo anterior podemos concluir que conforme los organismos se fueron haciendo más complejos en el curso de la evolución biológica, sus células se fueron diferenciando, de tal modo que algunas de ellas, como las que fabrican y secretan hormonas, se han especializado precisamente en eso, en desarrollar mecanismos de comunicación con otras células del organismo. Esta especialización de las células en las glándulas de secreción interna incluye la posibilidad —y la necesidad— de liberar al exterior una molécula mensajera, a la cual hemos llamado transmisor (en el caso particular, hormona), el cual viaja a distancia usando como elemento transportador la sangre, líquido vital que llega a todas las células del organismo. Entre paréntesis, la sangre lleva no solamente las hormonas sino también el oxígeno, el alimento, los anticuerpos (que por cierto también son proteínas, en este caso capaces de reconocer a proteínas ajenas y dañinas, como las de las bacterias o los virus), y recoge al mismo tiempo los desechos de todas las células para su posterior eliminación del organismo a través de otras células encargadas de hacerlo, como el riñón o los pulmones.
COMUNICACIÓN EN EL SISTEMA NERVIOSO
Necesitamos ahora establecer un nexo entre el tema de este capítulo y las ideas revisadas en el anterior. Lo que equivale a preguntarnos: dentro del esquema general de mecanismos de afinidad, reconocimiento y comunicación intercelular mediante moléculas, ¿qué lugar ocupan las células nerviosas?, ¿qué tipo de afinidad o afinidades utilizan las neuronas que, como hemos visto en el primer capítulo, son las que nos permiten entrar en contacto con el mundo exterior y al mismo tiempo controlan de manera involuntaria gran parte de las funciones de nuestro cuerpo?
Dejemos claro, desde este momento y como una respuesta parcial a estas preguntas
que, de entre todas las células vivientes no hay alguna que se haya especializado
tanto en los mecanismos de comunicación como las nerviosas. Su diferenciación
en este sentido es tan grande, que las células nerviosas adultas incluso han
perdido la capacidad de reproducirse. Dicho de otra manera, son células que
han abandonado prácticamente toda otra función para dedicarse exclusivamente
a comunicarse entre sí y con otras células del organismo. A esto habría que
agregar además que, conjuntamente con la progresiva especialización para optimizar
la comunicación, en el curso de la evolución de las especies las neuronas que
en los organismos primitivos estaban más o menos dispersas a lo largo y a lo
ancho del cuerpo, como en los invertebrados, progresivamente se fueron centralizando,
es decir, agrupando en una masa cada vez más compacta, más grande, más compleja
y con funciones globales cada vez más elaboradas y sofisticadas, hasta que finalmente
se llegó al cerebro humano. (Figura 18.)
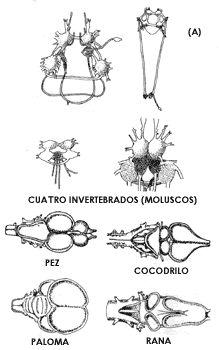
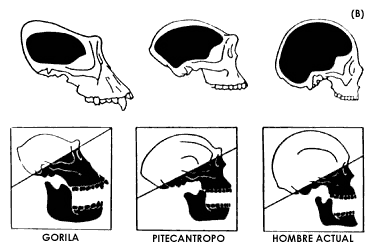
Figura 18. El sistema nervioso central progresivamente se fue concentrando en masas cada vez más compactas y más complejas en la porción cefálica de los organismos. En los moluscos, por ejemplo (sección superior de A) el cerebro está constituido por ganglios independientes unidos por cadenas de fibras nerviosas. En los peces, anfibios, reptiles y aves ya existe una masa alojada en el cráneo. En los mamíferos, particularmente en los primates (sección B), grupo al cual pertenecen el gorila y el hombre, el cerebro ocupa un espacio mucho mayor, en proporción tanto del resto de la cabeza como del tamaño del cuerpo en general. Entre el gorila, el Pitecantropo (uno de los hombres primitivos) y el hombre actual, es muy claro el progresivo desarrollo del cerebro que cada vez ocupa más volumen en relación con la cara y con la mandíbula. Esta diferencia se aprecia claramente comparando las superficies por arriba y por debajo de las líneas diagonales en la última parte de la sección B.
Debido a esta enorme centralización de las células específicamente dedicadas a la comunicación, la capacidad de almacenar información se incrementó prodigiosamente, pues no sólo fue aumentando el número de neuronas sino que simultáneamente, la complejidad de sus conexiones e interacciones se fue haciendo cada vez más notable por la extraordinaria multiplicidad de las combinaciones posibles en el funcionamiento de los conjuntos de neuronas. Pero éste será el tema de un próximo capítulo. Por lo pronto y para continuar con nuestra línea de alcances y progreso en distintos niveles de comunicación, consideremos brevemente qué sucedió cuando apareció el lenguaje como una consecuencia de la centralización y complejidad a la que nos referimos. Mediante este inesperado, poderoso medio, ya no una molécula, ni una célula, ni muchas células, sino un organismo completo mediante un órgano, el cerebro, era capaz de comunicarse con otros organismos de una manera inmediata, directa, clara y concisa, y además de transmitirles lo que simultáneamente había ido surgiendo en esa masa de neuronas centralizadas, organizadas e integradas: sensaciones, emociones, deseos, impresiones, tristezas, enojos, alegrías. De este modo la socialización —que ya había hecho su aparición en la evolución de varias maneras, particularmente en insectos como las hormigas y las abejas— alcanzó dimensiones insospechadas. Y como parte fundamental de esa nueva dimensión en la socialización, a un nivel de lo que es ya una persona, es decir, un ser consciente de sí mismo, capaz de reflexionar, se empezaron a manifestar afinidades de un carácter totalmente nuevo, más allá de los contactos físicos, en sensaciones y emociones como la amistad o el amor en todas sus manifestaciones. Y sin embargo, no debemos olvidar que para manifestar cualquiera de estos sentimientos y emociones, así sea a través del lenguaje, el cerebro debe emplear neuronas que muevan los músculos, que contraigan las extremidades, que flexionen o extiendan la lengua, los labios, las cejas, que endurezcan la mirada, que eleven la voz, que hagan vibrar las cuerdas vocales, que inclinen el cuerpo para abrazar, que alteren la expresión de la cara en una sonrisa, en un llanto, en un gesto de ternura...
Se forman así los grupos humanos, las sociedades, los países. Y con esto se establecen mecanismos de comunicación entre esos grupos. No seguiremos por el momento esta línea de continuidad en la evolución y diferenciación de los mecanismos de comunicación. Sí quisiera, sin embargo, tocar uno de los puntos centrales de esta evolución, tan importante como el lenguaje: me refiero al lenguaje escrito. Por primera vez en la historia de la vida sobre la tierra, se generó un mecanismo mediante el cual la comunicación podía pasar a otras generaciones por venir, mediante la preservación de las ideas, los conocimientos, los pensamientos y las emociones convertidas en poesía, cuentos, novelas, libros de texto, ensayos, monografías, historia, y todo cuanto fuera posible plasmar en símbolos más perdurables que las palabras. ¡El lenguaje escrito! ¡Los símbolos permanentes, transmisibles inclusive después de la muerte! ¡La comunicación traspasando los límites de lo que dura la vida del cerebro!
Pero también el lenguaje escrito, con todas sus posteriores extensiones debidas a los adelantos tecnológicos, como las máquinas de escribir, grabadoras, computadoras, impresoras, rotativas, etc., es obra de las neuronas agrupadas en esa masa que tenemos dentro de nuestros cráneos y que llamamos cerebro. Es conveniente ahora, por lo tanto, regresar a las neuronas. Esas células que son la cumbre de la evolución biológica, los elementos vivos más sensibles para percibir, transmitir, comunicar, coordinar y organizar. Veamos, pues, de cerca a las neuronas. Conozcamos cómo son, qué forma tienen, y cómo están organizadas dentro del cerebro.
¡Saludemos a las neuronas!