IV. ¿QUÉ LENGUAJE HABLAS?
| El pájaro ha venido/ a dar la luz: de cada trino suyo/ nace el agua. Y entre agua y luz que el aire desarrollan/ ya está la primavera inaugurada,/ ya sabe la semilla que ha crecido,/ la raíz se retrata en la corola,/ se abren por fin los párpados del polen. Todo lo hizo un pájaro sencillo/ desde una rama verde. |
"La primavera", PABLO NERUDA |
EN 1921 el alemán Otto Loewi realizó un experimento que iba a
tener consecuencias muy interesantes e importantes para el desarrollo futuro
del conocimiento del lenguaje que las neuronas hablan y lo que se dicen entre
sí. Loewi trabajaba sobre lo mecanismos de contracción del corazón de la rana,
y con este objetivo, utilizaba un modelo experimental que se ha usado mucho
en fisiología cardiaca: mantener el corazón de la rana fuera del cuerpo de ésta,
aislado de todos los otros órganos en una caja de Petri o pequeño vaso de vidrio.
El corazón está bañado en una solución que contiene las sales más abundantes
del plasma y de los líquidos que se encuentran normalmente fuera de las células
en todos los tejidos animales, por ejemplo el cloruro de sodio, que es la sal
más abundante.
En estas condiciones, si hay suficiente oxígeno en la solución, el corazón de la rana puede latir durante dos o tres horas, debido al automatismo característico del músculo cardiaco —recordemos que el corazón no es otra cosa que un músculo con ciertas propiedades especiales, a pesar de que sigue siendo considerado el órgano con el que amamos y sentimos las emociones—. Gracias a esta propiedad del automatismo, el corazón es capaz de contraerse casi como si estuviera aún dentro del organismo y siguiera conectado por medio de las arterias y venas al resto del mismo. Sin embargo, esto no quiere decir que el corazón dentro del cuerpo funcione independientemente; al contrario, como cualquier otro órgano del cuerpo, el corazón está sujeto a una regulación muy importante ejercida por el cerebro, precisamente por aquella parte del sistema nervioso que en el capítulo I denominamos autónoma porque funciona sin la participación de la voluntad. Es mediante esta regulación autónoma que no necesitamos pensar para ordenarle al corazón que acelere su frecuencia de latidos cuando los órganos requieren de más sangre, por ejemplo, cuando corremos o tenemos alguna experiencia que nos produce enojo o un estado de alerta. En estos casos, la frecuencia de latidos del corazón aumenta por acción de la parte autónoma del cerebro, a través de los axones de las neuronas localizadas en ciertos núcleos celulares del tallo cerebral. Sin embargo, otros axones de neuronas diferentes, que llegan al corazón por otro nervio, llevan una información opuesta: en lugar de acelerar la frecuencia cardiaca, la disminuyen. De este modo el sistema nervioso autónomo controla la frecuencia cardiaca, aumentándola o disminuyéndola según lo requiera el organismo.
SUSTANCIAS QUÍMICAS QUE CAMBIAN LA FRECUENCIA CARDIACA
Regresemos ahora a Otto Loewi y sus experimentos del corazón aislado de la rana. A Loewi se le ocurrió que el nervio que causaba una disminución de la frecuencia de latidos del corazón quizá lo hacía mediante una sustancia química, la cual sería liberada por el nervio cuando éste se estimulaba. Como es posible aislar al corazón junto con su nervio, Loewi pensó que si su idea era correcta, debería ser posible demostrarla mediante el siguiente experimento: estimular el nervio encargado de disminuir la frecuencia cardiaca en un corazón aislado, tomar un poco de la solución en que ese corazón está sumergido, y agregarla a otro corazón aislado. Si realmente el nervio ejerciera su acción mediante la liberación de una sustancia, ésta debería estar presente en la primera solución y por consiguiente al agregarla al otro corazón de velocidad normal, éste debería disminuir su frecuencia de latidos.
Loewi refiere que esta idea se le ocurrió por primera vez en un sueño que al despertar tenía muy claro, pero que, como a todos nos has ocurrido muchas veces, le fue imposible recordar después por más esfuerzos que hizo. Sin embargo, afortunadamente para el futuro de la investigación sobre el sistema nervioso, pocos días después volvió a tener el mismo sueño. En esta ocasión Loewi no quiso correr riesgos, y al recordar el sueño lo apuntó inmediatamente. La idea le pareció tan atractiva, que sin pensarlo mucho, muy temprano al día siguiente, se precipitó a su laboratorio a hacer el experimento, el cual, como ya habrá supuesto el lector de estas líneas, resultó como él esperaba: al agregar la solución del corazón cuyo nervio había estimulado, al segundo corazón, éste inmediatamente disminuyó también su frecuencia de contracción. Fue verdaderamente una fortuna que Loewi no esperara mucho tiempo para hacer el experimento en la segunda ocasión que soñó la idea, pues, como él mismo confiesa, si lo hubiera pensado un poco más quizá nunca se hubiera animado a hacerlo, pues las probabilidades de éxito eran muy bajas considerando una serie de factores, como el hecho de que la hipotética sustancia liberada del nervio seguramente estaría en cantidades extraordinariamente pequeñas, y además al liberarse al medio se diluiría enormemente por el relativamente gran volumen de éste, y en esas condiciones era muy improbable que pudiera observarse el efecto esperado en otro corazón. Así pues, debemos a la prisa de Loewi el que no haya pensado de antemano tantas posibles objeciones experimentales y se haya animado a hacer lo que hizo. En efecto, como a pesar de todo el experimento resultó positivo —veremos en este capítulo la trascendencia del hallazgo— se descubrió que las sustancias químicas eran capaces de liberarse de los axones y actuar sobre otros órganos, en este caso el corazón.
La historia del descubrimiento de Loewi nos da la entrada para unir los dos capítulos precedentes y tratar de entender cómo se comunican las neuronas. Por una parte, en el capítulo II revisamos cómo los mecanismos de comunicación entre las moléculas se fueron haciendo más y más complejos en el curso de la evolución biológica, de tal manera que, al aparecer las macromoléculas, especialmente las proteínas, la manera de cómo las moléculas se pueden reconocer entre sí cambió profundamente. Vimos cómo este cambio implicó una especificidad en dicho reconocimiento, de modo que sólo las moléculas con formas que tuvieran afinidad entre sí pueden unirse, combinarse o interactuar, y cómo todo esto se debe a la información contenida en la estructura de las proteínas. Por otra parte, en el capítulo III revisamos cómo es una neurona. Vimos que su forma particular incluye, además del soma, las dendritas o parte receptora de la información, y el axón o parte emisora de la información. El resto de este capítulo lo dedicaremos a ver la relación de estos dos capítulos entre sí y con el descubrimiento de Otto Loewi.
LA INFORMACIÓN QUE UTILIZAN LAS NEURONAS ES LA ELECTRICIDAD
Empecemos por ser un poco más precisos en lo que entendemos por la información
que es recibida por las dendritas y emitida por el axón. Para Aristóteles, la
información —aunque él no la llamaba así sino el "espíritu animal" o "animado"
(animal por referirse al ánima o alma— residía en la sangre, la cual era
impulsada por el corazón, asiento del alma. René Descartes, en la primera mitad
del siglo XVII,sin dar una explicación más exacta, amplió el concepto
aristotélico de un sistema hidráulico en el que se movía el "espíritu animado".
Al considerar ciertos reflejos, como los que ocurren cuando retiramos sin darnos
cuenta un brazo o una pierna del fuego cuando sentimos un calor demasiado intenso,
Descartes postuló que el fuego causaría un desplazamiento de la piel, el cual
sería transmitido por una "cuerda" —los nervios sensoriales que parten
de la piel— hasta el cerebro, en el cual se abriría como consecuencia un
poro o agujero que permitiría la comunicación entre el nervio y el ventrículo
cerebral. Por este poro pasaría entonces el espíritu animado hacia el interior
de los nervios —que él postulaba sería hueco— , de modo que al llegar
el espíritu a los músculos, a través de los nervios, éstos se inflarían y como
primera consecuencia se contraerían y entonces, la pierna o el brazo se retiraría.
(Figura 23.) Estos conceptos de Descartes ciertamente corrigen en gran medida
la equivocación aristotélica, ya que le dan a los nervios la capacidad de conducir
la información, en vez de adjudicar este papel a la sangre, pero conservan dos
errores muy importantes: primero, la idea del espíritu animado como la información
que es enviada, inicialmente de la piel al cerebro y después del cerebro al
músculo, y segundo, conceden a los ventrículos cerebrales —que son verdaderos
huecos que existen en el interior del cerebro, en donde se encuentra el líquido
cefalorraquideo— un papel importante en este proceso, ignorando al propio
tejido cerebral. Asimismo, la naturaleza del espíritu animado, lo que en este
libro llamamos información, continúa completamente indefinida, pues sólo se
le considera como la manifestación del alma. Sin embargo, otro concepto importante
—y correcto— de Descartes es que en un reflejo, como lo hemos ya mencionado
en el primer capítulo para el caso de reflejo rotuliano, la información parte
de un órgano receptor y regresa a un músculo, el cual responde de manera automática
a dicha información.
Tuvo que transcurrir un siglo a partir de Descartes, es decir, llegar casi
a fines del siglo XVIII, para que se empezara a conocer la identidad
de la información transmitida por los nervios.
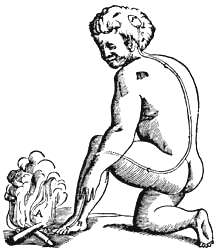
Figura 23. En este dibujo de Descartes, publicado en 1637, el filósofo y naturalista francés ilustra su idea del reflejo de retirar la pierna del fuego. Como se menciona en el texto, aunque el concepto de reflejo es en esencia correcto, la naturaleza de la señal que viaja por los nervios y de los nervios mismos y el cerebro, estaba muy lejos de ser correcta.
El inicio de este descubrimiento fue un hallazgo del italiano Luigi Galvani quien realizando experimentos de electricidad observó que si pasaba una corriente eléctrica de un metal a otro, colocando las patas de una rana entre los dos metales, los músculos de esas patas se contraían, Galvani demostró así que los músculos respondían con una contracción, a la excitación por medios eléctricos. Y fue más allá en sus investigaciones sobre esta propiedad del músculo que llamó "electricidad animal", realizando otro experimento que, además de interesante en sí mismo, planteó por primera vez de una manera directa la posibilidad de que los nervios estimulen los músculos por medio de corrientes eléctricas. Este experimento consistió en formar un circuito eléctrico mediante la inserción de un gancho de cobre en la médula espinal de una rana, de tal manera que este gancho, a través de la médula espinal, se conectaba con otro metal. En estas condiciones, durante una tormenta eléctrica o mediante la estimulación de la médula espinal con una máquina electrostática (una máquina de aquella época capaz de generar chispas) la pata de la rana se movía por la contracción rápida de sus músculos. Posteriormente, Galvani demostró que lo mismo ocurría cuando, en vez de la médula espinal, eran los nervios que llegaban a los músculos de la pata los que eran estimulados eléctricamente. Para Galvani estos hallazgos demostraban que los tejidos de la rana eran capaces de generar electricidad, la cual era probablemente de naturaleza diferente a la de la electricidad física, es decir, habría una electricidad animal y otra electricidad no animal. Sin embargo, Alessandro Volta, quienes años después construiría la primera batería artificial, la pila voltaica, demostró que en los experimentos de Galvani la electricidad no se originaba en los tejidos directamente, sino en los metales con que estaban en contacto, y lo que realmente hacían los nervios era conducir esa electricidad hasta el músculo, provocando que éste se contrajera.
A partir de los experimentos que acabamos de relatar se fue afinando y consolidando
la idea de aquello que Aristóteles bautizó como espíritu animado y Descartes
consideró como transmitido por los nervios para "inflar" los músculos, era en
realidad electricidad. Es curioso que desde dos siglos antes, en el siglo XVI,
se conocía un fenómeno que indicaba una importante relación entre la vida de
ciertos animales y la electricidad. En efecto, las descargas eléctricas producidas
por las anguilas y otros peces eléctricos eran conocidas desde esa época. En
el siglo XIX, Faraday, en sus extensos estudios sobre electricidad,
realizó experimentos sobre la naturaleza de la electricidad generada por estos
peces, y concluyó que era exactamente igual al generado por cualquier otro método,
pila o relámpagos y, además, postuló que había una relación entre el sistema
nervioso de estos peces y su órgano eléctrico.
Muchos estudios posteriores, ya en nuestro siglo, han confirmado de manera muy precisa que, en efecto, la naturaleza de ese flujo que viaja por los nervios y hace contraer los músculos es eléctrica. Así, como los nervios no son otra cosa que un conjunto de axones de las neuronas sensitivas que llevan la información hacia la médula espinal y el cerebro, y de las motoneuronas que la llevan hasta los músculos, podemos concluir que la información de que hemos venido hablando repetidamente, la cual cada neurona es capaz de recibir y enviar, es información de naturaleza eléctrica; que cada axón, por consiguiente, genera en su sitio de origen una corriente eléctrica y que es capaz de conducirla a todo su largo, sin perder intensidad, hasta el sitio donde termina y establece comunicación ya sea con un músculo, una glándula endocrina, el corazón o, si se trata de una neurona del interior de la médula espinal o del cerebro, con otras neuronas. Ésta es la información que buscaba Aristóteles y que Descartes determinó que viajaba por los nervios huecos: la electricidad. Éste es, pues, el lenguaje de las neuronas.
LA COMUNICACIÓN ENTRE LAS NEURONAS ES QUÍMICA
Es éste un punto clave de nuestro relato a propósito del lenguaje que hablan las neuronas. Cada neurona habla, efectivamente, en un lenguaje de electricidad. Pero, ¿qué sucede cuando la corriente eléctrica, palabra de este lenguaje, llega al final del axón, al sitio terminal en que debe comunicarse con la siguiente neurona?
Para contestar esta pregunta debemos primero recordar lo que hemos dicho en el capítulo anterior respecto a la monumental obra de Ramón y Cajal, confirmada después de manera precisa por la microscopía electrónica: las neuronas no se continúan una con otra; sus membranas no son comunes, sino que cada neurona está perfectamente limitada por la membrana que la rodea por todas partes, incluyendo naturalmente a las dendritas y al axón; las neuronas no son continuas una con otra sino contiguas, cada una es una entidad independiente. ¿Cómo es que una neurona puede entonces comunicarse con un músculo o con otra neurona? ¿Qué sucede cuando la corriente eléctrica que viaja a lo largo del axón llega al final de éste? La primera respuesta que podría uno pensar es que, si bien la membrana de la neurona no se continúa con la de la neurona siguiente, sus respectivas membranas podrían estar tan cerca una de la otra que la corriente eléctrica, al llegar al extremo del axón, sería capaz de "brincar" el pequeñísimo espacio que existe entre las dos neuronas. De hecho, es bien conocido que si nosotros acercamos dos cables eléctricos casi hasta tocarse y hacemos pasar una corriente eléctrica por uno de ellos, esa corriente puede transmitirse al otro cable. Así, si las membranas de las neuronas que se van a comunicar, o la de la neurona motora con la membrana de la célula muscular, estuvieran suficientemente cerca, es perfectamente posible que la información o corriente eléctrica de excitación que viaja a lo largo del axón, simplemente siga su camino saltando ese pequeñísimo espacio, de tal manera que la segunda neurona respondiera como si fuera una continuación de la primera neurona.
Siguiendo esta línea de pensamiento, veamos qué nos dicen las observaciones
realizadas con el microscopio electrónico respecto a las distancias existentes
entre la membrana del extremo terminal del axón y la membrana de la neurona
con la que éste se comunica. Dijimos microscopio electrónico, porque estas distancias
son de dimensiones tan pequeñas que ni el más potente microscopio de luz puede
dar una imagen exacta de estos arreglos entre membranas en los sitios de comunicación
entre las neuronas. (Figuras 24 y 25.) Sólo con el microscopio electrónico se
logran los aumentos necesarios y además de los aumentos, el poder de resolución
para poder distinguir una membrana de otra. Pues bien, las fotografías de microscopia
electrónica enseñan (ver figura 26) que efectivamente hay algunos sitios en
los que las membranas de las neuronas involucradas en la comunicación están
directamente adosadas una a la otra, dejando entre ellas un espacio extraordinariamente
pequeño de aproximadamente 2 nanómetros (un nanómetro es la millonésima parte
de un milímetro). ¿Es este espacio suficientemente pequeño para que pueda pasar
el lenguaje de la corriente eléctrica de una neurona a la otra? Un gran número
de estudios fisiológicos, por medio de electrodos colocados en las dos neuronas
que se están comunicando y que muestran este tipo de adosamiento entre las membranas,
indica que sí: efectivamente, cuando la membrana de la terminal axoníca está
tan cerca de la membrana de la otra neurona, la corriente eléctrica puede pasar
y por consiguiente, la segunda neurona al recibir esta información de la primera,
se excita eléctricamente y puede, a su vez, transmitir esa excitación a las
neuronas con las que se comunica.
 |
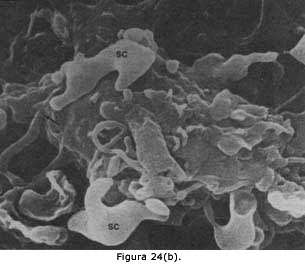 |
Figura 24. El microscopio electrónico "de barrido ", llamado
así porque los electrones barren la preparación en vez de atravesarla, como
sucede en el "de transmisión", permite ver claramente las terminales sinápticas
llegando a las neuronas. En la micrografía de la figura 24(a) se ven por lo
menos cuatro neuronas, la más grande con sus prolongaciones principales y
en el dibujo se observa un esquema de lo que la micrografía muestra. Es evidente
el gran número de terminales que una neurona puede recibir, sobre todo si
consideramos que, por razones de la preparación de la muestra para la microscopía,
sólo algunas de ellas permanecen adheridas y por ello no se ven los cientos
de terminales que llegan a las dendritas (compárese la extensión de la dendritas
en esta fotografía con la observada en microscopía de luz con neurona teñida
en su totalidad. Figuras 19 y 20). En la micrografía electrónica de barrido,
figura 24(b), se observa una neurona que también tiene sobre su superficie
algunas terminales ( la flecha señala un axón terminado en el cuerpo de la
neurona), y además se observan dos células (SC) que no son neuronas sino astrocitos,
cuya función se menciona en la última parte del Capítulo III.
Figura 25. Esta micrografía fue tomada en un microscopio de luz —equivale
a la figura 24 con microscopio electrónico de barrido—. en este caso la
neurona se encuentra cortada y es imposible verla en volumen, sin embargo, muchas
observaciones son visibles las terminales axónicas que la rodean. Los detalles
estructurales de estas terminales y de la zona de contacto con la membrana de
la neurona sólo son visibles con el microscopio electrónico de transmisión (ver
figura 26).
Muchas observaciones con el microscopio electrónico han demostrado de manera
contundente que en la gran mayoría de los sitios de comunicación entre las neuronas
las membranas de éstas tienen ciertas características especiales y están separadas
por un espacio aproximadamente 10 veces mayor que el que acabamos de describir,
es decir de aproximadamente 20 nanómetros. Estos sitios precisos de comunicación
entre las neuronas se llaman sinapsis y por ello, el espacio entre membranas
recibe el nombre de espacio sináptico el cual es demasiado grande para
que la corriente eléctrica lo pueda brincar. Lo mismo ocurre en los sitios en
que las terminales de los axones de las neuronas motoras se comunican con los
músculos: el espacio sináptico entre la membrana de la terminal del axón y la
membrana de la célula muscular es demasiado ancho para que la corriente eléctrica
lo pueda saltar. La pregunta que ahora debemos hacernos es entonces obvia: si
el lenguaje del axón es el de corriente eléctrica pero en los sitios en que
el axón termina y se debe comunicar con un músculo o con la siguiente neurona,
ese lenguaje ya no puede ser entendido y si además, lejos de ser excepción
esto es lo que ocurre en la inmensa mayoría de los sitios en que las neuronas
se comunican dentro del cerebro, ¿cómo es que tal comunicación se lleva a cabo?
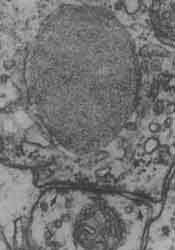
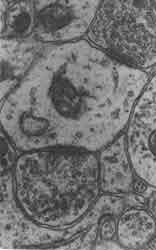
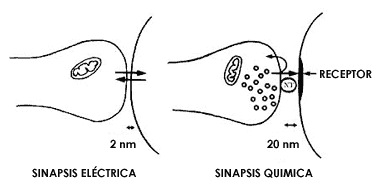
Figura 26. Los detalles arquitectónicos y la estructura fina de los sitios de comunicación entre las neuronas, llamados sinapsis, se observan en estas micrografías electrónicas de gran aumento (un centímetro en las fotografías equivale aproximadamente a dos diezmilésimas de milímetro). En la figura 26(a) se observa una sinapsis eléctrica en la que las membranas de las células que se comunican están muy cerca una de otra; como se ilustra en el esquema izquierdo de la figura 26(c) y se discute en el texto, esta cercanía permite a la corriente eléctrica brincar de una neurona a la siguiente. En contraste, la micrografía de la figura 26(b) muestra que en las sinapsis químicas el espacio entre las membranas es demasiado para que la corriente eléctrica lo pueda saltar, y se requiere por tanto de un transmisor (NT) químico ( de aquí el nombre de sinapsis química). En la foto se observa una dendrita cortada transversalmente, sobre la cual dos terminales axónicas una a la izquierda y otra abajo a la derecha, establecen sinapsis. Las terminales se reconocen porque poseen en su interior pequeñas vesículas esféricas que se agrupan cerca de la membrana en el sitio de la sinapsis. Las sinapsis químicas son, por mucho, las más abundantes e importantes en el cerebro de los mamíferos. Las fotografías han sido tomadas de C. S. Raine, Basic Neurochemistry, Little Brown and Co.
El último cuestionamiento corresponde a un lenguaje eléctrico que ya no puede ser transmitido. Podemos decir entonces que ese lenguaje no es entendido, y que por lo tanto debe ser traducido. La pregunta anterior se convierte así en: ¿cuáles son los mecanismos de traducción del lenguaje eléctrico de los axones a otro lenguaje que pueda ser entendido por la neurona siguiente o por el músculo, a través de ese espacio de 20 nanómetros? La respuesta a esta pregunta tan fundamental nos la da en principios los experimentos de Otto Loewi que hemos descrito más arriba. Ese mecanismo de traducción consiste en que, como efecto directo de la llegada del impulso eléctrico hasta la terminal del axón, ésta libera una sustancia química a través de su membrana, la cual sí puede cruzar el espacio de 20 millonésimas de milímetro que la separa de la siguiente neurona y así llegar a ella. (Figura 27.) Y ¿qué sucede cuando esa molécula alcanza a la siguiente neurona? El lector atento del capítulo II de este libro tendrá sin duda la respuesta en la punta de la lengua, pues en ese capítulo vimos cómo las moléculas pueden comunicarse entre sí de acuerdo a sus afinidades, mediante un reconocimiento específico de una molécula hacia otra. Además revisamos en ese capítulo II cómo algunas de estas macromoléculas capaces de reconocer a otras están colocadas en el seno mismo de la membrana, embebidas en ellas, de modo tal que pueden reconocer a otras moléculas que se acerquen y que posean las características de afinidad para que puedan ser reconocidas. Inclusive dijimos que esas moléculas de la membrana reciben el nombre de moléculas receptoras, justamente porque eran capaces de recibir, de aceptar, a otra molécula con las características adecuadas para ser reconocida. Mencionamos también que un ejemplo muy claro de las moléculas que viajan desde grandes distancias y son reconocidas por receptores específicos que existen en muchas de las células del organismo eran las hormonas, las cuales podían así ser definidas como mensajeros que llevaban información desde el sitio en que eran producidas hasta todas esas otras células del organismo. Pues bien, consideremos ahora a estas otras moléculas, que son liberadas, es decir, secretadas, por las terminales de los axones en los sitios de comunicación. Aunque ciertamente no viajan tanto como las hormonas, ni son secretadas hacia la sangre para ser ahí transportadas, el hecho es que sí recorren la distancia de 20 nanómetros que separa las dos membranas de las células que se están comunicando, y al igual que las hormonas requieren de una molécula receptora en la membrana de la segunda neurona para ser reconocidas como mensajeros y transmitir la información de la célula de las cuales son liberadas. Como estos mensajeros se forman en las neuronas, que hemos definido como las células constituyentes del sistema nervioso que determinan su funcionamiento, estas moléculas mensajeras reciben el nombre de neurotransmisores.
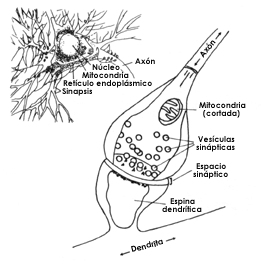
Figura 27. Este esquema de la sinapsis química ilustra las vesículas sinápticas contenidas en el interior de la terminal axónica, así como el espacio sináptico entre la membrana de dicha terminal y la membrana de la siguiente neurona; en este caso la sinapsis se establece con una de las llamadas espinas dendríticas, que son pequeñas salientes de las dendritas más gruesas (véase la figura 19 y el primer dibujo de la 20). Se observa también en el interior de la terminal una mitocondria; este importante organelo celular es el encargado de aprovechar la energia obtenida de los alimentos, de modo que mediante una serie de reacciones químicas, la energía es almacenada en una molécula especial y puede ser usada para las distintas funciones celulares. En el dibujo superior se muestra una neurona con el cuerpo cortado en parte para dejar ver su interior, que contiene un núcleo, mitocondrias y el llamado sistema retículo endoplásmico, que es el sitio en el que todas las células sintetizan sus proteínas. En el exterior de la neurona se observan numerosas terminales axónicas haciendo sinapsis con el cuerpo o las dendritas de la neurona, como en la figura 24.
NEUROTRANSMISORES Y RECEPTORES
Ya estamos en condiciones de definir el mecanismo de traducción que usan las neuronas para comunicarse: mediante la liberación de un neurotransmisor, y la combinación de éste con una molécula receptora localizada en la membrana de la segunda neurona, traducen el lenguaje eléctrico del axón a un lenguaje químico. Sin embargo, tenemos otra interesante pregunta todavía frente a nosotros: ¿qué hace esa segunda neurona una vez que ha reconocido y aceptado al neurotransmisor sobre su membrana, mediante el receptor correspondiente? Para contestar esta nueva y muy importante pregunta tenemos que hacer otra consideración respecto al lenguaje eléctrico de las neuronas, que es la siguiente. En términos de excitación eléctrica, estrictamente hablando, una neurona puede hacer solamente dos cosas: a) emitir, como hemos revisado, una señal eléctrica a lo largo de su axón, o b) no emitirla. Si emite la señal, una vez que ésta se ha iniciado en el sitio de origen del axón, la corriente eléctrica será indefectiblemente conducida hasta todas las terminales de dicho axón, de la misma manera que una vez prendida la mecha de un polvorín la chispa continuará su avance a lo largo de la mecha hasta su destino final. En otras palabras, una vez que la neurona ha disparado la corriente eléctrica, ésta llegará a su destino sin que la neurona pueda hacer algo para impedirlo. Pues bien, si esto es lo que sabe hacer una neurona, es inevitable concluir que cuando un neurotransmisor se combina con su receptor, algo debe pasar en la membrana de la neurona que reconoció y aceptó al neurotransmisor para que a su vez dispare una señal eléctrica desde el origen de su axón, y pueda ésta ser conducida a lo largo de dicho axón. Nos interesa, pues, saber qué es ese "algo" que sucede en la membrana.
Lo que sucede en la membrana es el mecanismo mediante el cual el receptor, en el momento de reconocer al transmisor, es capaz de excitar a toda la membrana de la neurona a la cual pertenece, para que ésta dispare sus señales. Ese mecanismo es muy interesante. Como en el medio externo a las neuronas hay átomos que tienen carga positiva, especialmente sodio —el cual ya comentábamos en el capítulo II que se encuentra cargado cuando está en forma de la sal cloruro de sodio—, lo que sucede cuando el transmisor se combina con el receptor es que se abren unos pequeñísimos canales en la membrana, y por estos canales penetran rápidamente los átomos de sodio. Dado que el sodio tiene carga positiva, su entrada genera cambios en las características eléctricas de la membrana, y la consecuencia de estos cambios es que la neurona se excita al grado de generar en su axón los impulsos eléctricos de que hemos venido hablando. De esta manera, mediante una íntima relación de las moléculas receptoras con los canales que permiten el paso del sodio, se lleva a cabo una nueva traducción del lenguaje, podríamos decir que en reversa: inicialmente el lenguaje eléctrico se convirtió en un lenguaje químico mediante la liberación del neurotransmisor en las terminales axónicas mientras que ahora, como resultado de la combinación del transmisor con el receptor, a través de la apertura de canales para sodio el lenguaje químico es reconvertido en lenguaje eléctrico, cerrándose el mecanismo de comunicación entre neuronas.
Este mecanismo de comunicación eléctrico-químico-eléctrico que hemos descrito, ocurre de manera muy similar entre dos neuronas y entre una neurona y el músculo al que la neurona llega para que éste se contraiga. Es así como podemos mover cada uno de nuestros músculos, mediante la acción de una sustancia que las terminales nerviosas liberan en el sitio en que las motoneuronas se comunican con ellos. Esto quiere decir que la membrana de las células musculares también tiene moléculas receptoras, y además que el músculo también tiene un lenguaje eléctrico que le permite contraerse.
Debemos ahora introducir una complicación más en nuestro estudio de la comunicación entre las neuronas. Recordemos el tantas veces mencionado experimento de Loewi. Decíamos que lo que hacía la sustancia que parecía liberarse de los axones que llegan al corazón no era aumentar la frecuencia de los latidos, sino disminuirla, frenarla. Pensemos un poco en lo que esto quiere decir. Antes describimos un mecanismo que permitía a la segunda neurona disparar impulsos, es decir, excitarse, y dijimos también que al músculo le sucedía algo parecido, sólo que en este caso en lugar de responder a la excitación con el envío de señales eléctricas lo hacía contrayéndose. Y ahora, al recordar el experimento de Loewi, vemos con sorpresa que en vez de que el músculo cardiaco se contraiga más al recibir el neurotransmisor, se contrae menos. ¿Cual es la conclusión?. Inevitablemente es necesario concluir que ese transmisor en vez de excitar al corazón lo inhibe. Traslademos ahora esta conclusión a las neuronas, mediante una pregunta, ¿existen neurotransmisores que al ser liberados y combinarse con los receptores de otras neuronas las inhiban en vez de excitarse? Y si la respuesta es afirmativa, ¿cómo es que esto sucede? Y como tercera pregunta inmediata, de evidente trascendencia, ¿qué importancia tendría para el funcionamiento del cerebro el que las neuronas puedan inhibirse en vez de excitarse, es decir frenar o disminuir la capacidad de disparar señales eléctricas a través de su axón, en vez de emitirlas?
Contestemos a estas preguntas. La respuesta a la primera pregunta es inmediata y muy simple: sí existen neurotransrnisores que al combinarse con sus receptores en la siguiente neurona, causan una inhibición y no una excitación de ella, es decir, disminuyen la probabilidad de que esa neurona dispare señales eléctricas por su axón. Es más, de una vez vale la pena decirlo, las neuronas que tienen esta acción inhibidora sobre otras neuronas son mucho más numerosas que aquellas de cuya acción resulta una excitación.
La respuesta a la segunda pregunta es un poco más complicada, pero no muy difícil después de lo que hemos dicho respecto a los mecanismos de excitación. Si en ellos entra un átomo cargado positivamente, como el receptor está acoplado a canales para átomos cargados negativamente, sucede un cambio en el mecanismo de excitación por inhibición. Así, cuando la comunicación entre las neuronas es de naturaleza inhibidora, se abren canales para átomos cargados negativamente, los cuales al penetrar al interior de la neurona la inhiben. (Figura 28.)
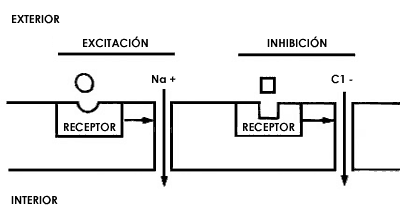
Figura 28. La proteína que actúa como receptor en la membrana de las neuronas, al reconocer y aceptar a un determinado neurotransmisor por su parte externa, que ve hacia el exterior de la célula, sufre un cambio en su estructura tridimencional el cual produce una modificación en otras proteínas. Estas últimas son canales para sodio (Na+) o para cloro(Cl-), por lo que al modificarse por efecto de la combinacion transmisor-receptor dejan pasar estos átomos. Debido a que, como se describió en el texto, los átomos de sodio y de cloro tienen una carga positiva o negativa, respectivamente (véase la figura 9), al entrar, cambian las propiedades eléctricas de la membrana: cuando entra el sodio hay excitación de la neurona y cuando entra el cloro hay inhibición. El transmisor que produce excitación es distinto al que produce inhibición, y naturalmente también sus correspondientes receptores son diferentes, pues reconocen distintas moléculas.
Evidentemente debe haber una relación entre las cargas eléctricas de los átomos que entran a través de los canales abiertos por los transmisores y la corriente eléctrica que finalmente va a fluir a lo largo de todo el axón. Esta relación está dada por la membrana de la neurona, la cual es capaz de guardar o almacenar cargas eléctricas. Debido a esta propiedad, al entrar los átomos con carga positiva —en el caso de la excitación— o negativa —en el caso de la inhibición—, la membrana modifica su capacidad de almacenar esas cargas. El resultado de este cambio puede ser, por lo tanto, que aumente la diferencia de carga contenida en la membrana o que disminuya. Debido a las características especiales de la membrana y a la composición de los líquidos que se encuentran en el interior y en el exterior de la neurona, cuando disminuye la diferencia en la carga eléctrica de la membrana, por la entrada de átomos con carga positiva, la neurona se excita. Lo contrario sucede cuando esta diferencia de carga aumenta por la entrada de átomos con carga negativa: la neurona se inhibe.
Hasta ahora hemos visto cómo las dendritas y el axón reciben y envían la información eléctrica, respectivamente; cómo esa información eléctrica es traducida a un lenguaje químico para que por medio de los neurotransmisores se establezca la comunicación entre dos neuronas; y cómo esa comunicación puede ser de naturaleza excitadora o inhibidora, de acuerdo con los mecanismos de traducción en reversa, es decir, del lenguaje químico al eléctrico, por la apertura de canales para átomos que poseen carga eléctrica positiva o negativa. Pero aún no sabemos cómo la célula maneja esta información inhibidora o excitadora.
Para revisar este importantísimo aspecto de la función de las neuronas es necesario recordar brevemente la arquitectura de las comunicaciones entre las neuronas. Como anotamos en el capítulo anterior, cada neurona recibe una enorme cantidad de terminales nerviosas, cada una de ellas proveniente de otra neurona. Dijimos inclusive que ese número de conexiones podía ser de muchos miles sobre una sola neurona. Ahora nuestro objetivo es entender cómo la neurona maneja toda esa información que le llega simultáneamente tomando en cuenta que esa información es de naturaleza distinta, pues muchas de esas señales serán inhibidoras mientras que otras serán excitadoras. Es aquí que el soma de la neurona adquiere una gran importancia funcional, pues el soma es el maestro capaz de analizar toda esa información. En efecto, recordemos que lo único que podrá hacer la neurona como respuesta a toda la información que recibees enviar o no, la corriente eléctrica que viajará a lo largo de su axón, para a su vez comunicarse con otras neuronas. Así, lo que el soma debe hacer con toda la información excitadora o inhibidora que recibe en cada momento es integrarla, es decir, sumarla algebraicamente para obtener un resultado final. (Figura 29.) Esta suma algebraica implica que una excitación se opondrá a una inhibición y viceversa, pues son claramente de signo contrario, de manera que sucederá lo siguiente: supongamos que una neurona recibe 7 señales excitatorias simultáneamente y ninguna inhibidora. Estas 7 señales excitadoras serán suficientes para que el soma inicie el envío de la corriente eléctrica por su axón: la neurona ha sido excitada. Pero, ¿qué sucede si simultáneamente con la llegada de estos 7 estímulos excitadores llegan 3 inhibidores? La respuesta es clara: esta inhibición neutralizará a los estímulos excitadores, de manera que ahora la neurona no podrá ya enviar su señal a través de su axón: la neurona ha sido inhibida.
Lo que acabamos de revisar implica que la membrana de las neuronas es un verdadero mosaico de receptores, pues frente a cada membrana de cada una de los cientos o miles de terminales axónicas que lleguen a ella habrá un receptor, que además, por los mecanismos de especificidad en el reconocimiento entre moléculas que hemos revisado en el Capítulo II, será sólo para la molécula neurotansmisora que se libere desde la terminal que le corresponde y para ninguna otra molécula que pueda estar cerca en el medio extracelular. Así, los mecanismos de comunicación entre las neuronas resultan de una sofisticación y complejidad verdaderamente asombrosas. No solamente cada célula está anatómicamente comunicada exclusivamente con las otras neuronas que forman parte de la vía o circuito a que ella pertenece, sino que además en cada uno de los sitios de comunicación hay una especificidad absoluta respecto tanto a la molécula neurotransmisora que se está liberando desde la terminal, como a la molécula receptora que está en la membrana de la neurona que recibe, precisamente frente a la terminal de la otra neurona. Es por esto que hablamos de un mosaico en las membranas neuronales, un riquísimo mosaico en cada neurona, comparable en riqueza y complejidad a los vetustos, extraordinarios mosaicos bizantinos de San Marcos de Venecia, o de las tantísimas iglesias y monasterios bizantinos en Grecia y en Turquía.
LAS SINAPSIS EXCITADORAS E INHIBIDORAS EN LOS CIRCUITOS NEURONALES
Detengámonos ahora un momento para analizar las implicaciones y consecuencias que tiene sobre el funcionamiento de grupos de neuronas el hecho de que exista la inhibición. Pensemos para esto en un motor cualquiera, por ejemplo el de un automóvil. ¿Qué sucedería si no hubiera manera de detenerlo, de frenarlo una vez en marcha? ¿Qué tan eficiente sería su funcionamiento si sólo fuera posible acelerar o no acelerar? ¿Qué ventaja tiene, además, el hecho de poner un freno más constante cuando en una bajada muy pronunciada colocamos la palanca de velocidades en segunda o primera sin necesidad de mantener presionado el freno para que los mismos engranes impidan que siga acelerándose por simple efecto de gravedad? Probablemente no sea necesario ser más explícito en contestar estas preguntas. Es obvio que contar con un mecanismo capaz de frenar de manera continua, además del freno momentáneo, es lo que permite tener un control del movimiento de un vehículo, particularmente cuando su velocidad tiende a aumentar en forma espontánea, como en el caso de una bajada. La falta o la pérdida de un mecanismo frenador resultaría evidentemente en la pérdida del control de vehículo, con resultados catastróficos.
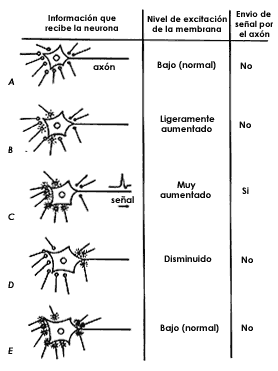
Figura 29. Cada neurona es capaz de integrar, sumando algebraicamente, la información que recibe de todas las terminales que llegan a ella y que en un momento dado están liberando su correspondiente neurotransmisor. El resultado de esta suma algebraica es que la neurona enviará información o no a lo largo de su axón, en forma de una señal eléctrica. Esta neurona recibe 7 sinapsis excitadoras (terminales blancas) y 3 sinapsis inhibidoras (terminales negras). A: Cuando ninguna de ellas está activa, liberando transmisor, la neurona está en reposo, el nivel de excitación de su membrana es normalmente bajo y naturalmente no hay envío de señales. B: Si 3 de las7 sinapsis excitadoras están activas, el nivel de excitación de la membrana está aumentando, pero no lo suficiente para que la neurona dispare la señal. C: Si las 7 terminales excitadoras están "encendidas", el nivel de excitación de la membrana será suficientemente alto para que dispare la señal a lo largo de su axón: se habrá alcanzado el umbral de disparo de esa neurona. D: Si solamente están activas las terminales inhibidoras, el nivel de excitación de la membrana estará obviamente disminuido, y evidentemente no habrá envío de señal. E: Si las 7 terminales excitadoras están activas, pero al mismo tiempo las 3 inhibidoras también están encendidas, el nivel de excitación será bajo y por lo consiguiente la neurona no disparará. Nótese que en esta última condición la neurona ha sido inhibida, puesto que no puede disparar a pesar de estar recibiendo el mismo número de estímulos excitadores que, en ausencia de inhibición, era capaz de causar el envío de señales.
Apliquemos estas ideas al control de la actividad neuronal. Por una parte, cuando un grupo de neuronas ha sido excitado hasta el punto de enviar señales a lo largo de su axón, sería posible inhibir su actividad mediante la llegada de estímulos inhibidores, sin necesidad de disminuir la excitación que le está llegando, y que fue determinante para que empezara a disparar. Por otro lado, si para mantener excitada una neurona es necesario que reciba digamos los 7 estímulos excitadores que hemos mencionado, bastaría con inhibir uno de ellos, es decir, inhibir a la neurona que está enviando esa excitación, para obtener un cese del envío de señales por la neurona en cuestión. Pero, además, supongamos ahora que hay neuronas qué tienen un ritmo endógeno de disparo, es decir que, de manera similar a las células del corazón de la rana, tienen la propiedad de estar en un estado de excitación espontánea, de tal manera que para mantenerla funcionando eficientemente no hay que excitarla, sino más bien controlarla mediante una inhibición: con menor control inhibidor se suelta demasiado, mientras que si se aumenta esa inhibición disminuye su actividad. En este caso estamos hablando de una actividad que depende totalmente del grado de inhibición que se ejerza para que sea eficiente, y prácticamente no es necesario ningún mecanismo excitador.
Probablemente el lector no necesite más para entender y convencerse de la importancia enorme que tienen los mecanismos inhibidores en el funcionamiento de las neuronas, particularmente cuando consideramos no una sola, sino un conjunto de ellas arregladas en circuitos o vías específicas que a su vez determinan cierta función particular del sistema nervioso.
Este tipo de repercusiones sobre circuito será el tema de nuestro siguiente capítulo. Sin embargo, por el momento y antes de pasar a él, necesitamos dedicar un espacio a las interesantes consecuencias que tiene sobre el funcionamiento de las neuronas, el hecho de que la comunicación entre ellas sea la naturaleza química y no eléctrica.
ALGUNAS VENTAJAS Y CONSECUENCIAS DE LA COMUNICACIÓN QUÍMICA
Si la comunicación fuera eléctrica, sólo pasando la corriente de una neurona a otra, como si fuera una continuación del axón, es claro que no podría haber inhibición. Solamente mediante el procedimiento de los neurotransmisores que al acoplarse con su receptor abren canales para átomos con carga negativa, es posible que las neuronas se inhiban. Esta es una primera y trascendental consecuencia de la existencia de los mecanismos químicos de la comunicación entre las células de la mente, pero no es la única. Recordemos que se trata de corrientes que fluyen como en un cable, con una comunicación eléctrica esas corrientes podrían pasar de una neurona a otra y viceversa, es decir, la comunicación entre dos neuronas sería bidireccional, lo cual podría tener una serie de consecuencia indeseables en términos del control de la información a lo largo de determinados circuitos. Por el contrario, mediante la existencia de los mecanismos químicos que requieren que un neurotransmisor se libere de la terminal axónica, se establece con precisión que la comunicación ocurra unidireccionalmente, es decir, sólo de la terminal axónica a la neurona siguiente, pero no a la inversa.
Además de las ventajas que mencionamos en el párrafo anterior, la existencia de mecanismos químicos en la comunicación interneuronal representa una ventaja adicional enorme: permite una serie de posibilidades de regulación o control de la misma transmisión de la información, que no podrían existir con una transmisión eléctrica. Por ejemplo, a través de la proteínas que mencionamos en el Capítulo II como responsables de que ocurran las reacciones intracelulares a una velocidad apropiada, es posible regular la velocidad de síntesis y de destrucción química de las moléculas que funcionan como neurotransmisores, determinando así en un momento dado qué tanto puede funcionar la comunicación en que participa ese transmisor. Además, también es posible para la neurona controlar la liberación del transmisor desde la terminal axónica, con lo cual también se puede regular la comunicación. Pero esto no es todo. Como la comunicación depende también de la combinación y la interacción del neurotransmisor con su receptor, imaginemos qué sucedería si la célula que recibe la información fuera capaz de alterar la estructura o la disposición de esa molécula receptora, de modo que se haga más sensible al transmisor, es decir que con menor cantidad de éste se produzca la apertura del canal correspondiente; o por el contrario, que el receptor pierda sensibilidad a su transmisor.
Es claro que mediante cualquiera de los mecanismos señalados en el párrafo anterior, o de la combinación de dos o más de ellos, es posible modificar la comunicación interneuronal. Dicho de otro modo, la existencia de los mecanismos químicos de la transmisión de información permite que dicha transmisión sea modificable, maleable, plástica. ¿Y no es precisamente ésta una de las propiedades más sorprendentes del cerebro, si pensamos en su capacidad de memorizar, de dar marcha atrás, de reconsiderar en función de nuevos datos, de aplicar los conocimientos o las experiencias previas a nuevos actos, de recordar, de asociar eventos, de recapitular, en una palabra de aprender? Desde esta perspectiva, el hecho de que el funcionamiento de la comunicación interneuronal sea química hace pensar que quizá muchos de los mecanismos de aprendizaje tengan su explicación en esta plasticidad de la comunicación, en el nivel interneuronal, ejercida mediante alguno, o varios, de los mecanismos que hemos mencionado.
Si consideramos las características y las ventajas que los mecanismos químicos conceden al lenguaje con el que se entienden las neuronas, tendremos que concluir necesariamente que eso se debe a una sorprendente sofisticación de la comunicación intermolecular que revisamos en el Capítulo II. En efecto, es gracias a la afinidad entre las moléculas, en este caso determinada por la increíble capacidad de las proteínas receptoras de reconocer específicamente a las moléculas neurotransmisoras, que fue posible para la naturaleza desarrollar los mecanismos de comunicación entre las neuronas. Es también éste el lenguaje químico que permite contraerse a los músculos —todos, desde los que usamos para expresarnos, hasta los de nuestras vísceras, incluyendo el corazón de cuyos latidos depende el viaje de la sangre hasta el más recóndito sitio en que una célula del organismo hace lo que tiene que hacer en el concierto del organismo completo—. Y es así como el lenguaje de las células de la mente se manifiesta y permite a su vez la manifestación de la comunicación entre los organismos, particularmente entre los seres humanos, a través de los lenguajes hablado y escrito o bien mediante el otro lenguaje, el corporal de las emociones y sentimientos plasmados en actitudes, entregas, amores y desamores.
Es muy poco lo que sabemos respecto al funcionamiento de grandes grupos de neuronas trabajando sincrónicamente para dar lugar a nuestros sentimientos, placeres, actos inteligentes, pensamientos, reflexiones, creaciones, conciencias, remordimientos, arrepentimientos, dudas, odios, iras, pasiones, tristezas y alegrías. En donde empezamos a tener una idea un poco más clara es en el papel de algunos núcleos de neuronas en la regulación de nuestra actividad muscular. Por esta razón en el siguiente capítulo ejemplificaremos el funcionamiento de grupos de neuronas, con su excitación, inhibición y regulaciones implícitas en el control muscular. Veremos también cómo es posible alterar los mecanismos químicos de la comunicación mediante el uso de una serie de sustancias y de cómo esto puede ser —en la actualidad ya de hecho lo es— de enorme utilidad para entender mejor cómo funcionan las neuronas y también para poder desarrollar fármacos que resulten útiles en el tratamiento de muchos padecimientos.