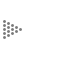V. ¿QUIÉN ERES?
| Ante la cal de una pared que nada nos veda imaginar como infinita un hombre se ha sentado y premedita trazar con rigurosa pincelada en la blanca pared el mundo entero: puertas, balanzas, tártaros, jacintos, ángeles, bibliotecas, laberintos, anclas, Uxmal, el infinito, el cero. Puebla de formas la pared. La suerte, que de curiosos dones no es avara, le permite dar fin a su porfía. En el preciso instante de la muerte descubre que esa vasta algarabía de líneas es la imagen de su cara. |
"La suma", JORGE LUIS BORGES |
AQUEL hombre trabajaba con ahínco. Era un obrero responsable, tranquilo, de modales austeros y suaves a quien le era fácil tener amigos y llevarse bien con sus compañeros y jefes. Este modo de ser, y sus deseos de trabajar y progresar, lo habían hecho —a los 25 años— uno de los jefes de cuadrilla que construían la línea del ferrocarril en el estado de Vermont, entre Rutland y Burlington, en los Estados Unidos. Su nombre era Phineas Gage.
Ese día, el 13 de septiembre de 1848, era necesario hacer volar una gran roca que se atravesaba en el camino de la vía, y Phineas decidió que ésa era una tarea que él personalmente debía realizar. Primero hizo una profunda perforación en la roca y la llenó de pólvora. Tomó entonces una barreta de fierro para apisonar la pólvora en el fondo de la perforación, antes de cubrirla con arena y provocar la explosión que volaría la roca para dar paso a la vía del tren. Y entonces ocurrió el accidente. Al apisonar la pólvora, la barreta se frotó contra una de las paredes de la perforación y provocó una chispa, la cual hizo explotar la dinamita. Con la fuerza de la explosión, la barreta de más de un metro de largo, 3 centímetros de diámetro y 6 kilogramos de peso salió disparada con enorme fuerza, convirtiéndose en un poderoso proyectil que encontró en su camino la cabeza de Phineas Gage. La barreta penetró por la parte superior de la órbita del ojo izquierdo, y en una fracción de segundo perforó el cerebro y salió por la parte superior del cráneo cerca de la frente de Phineas, y aterrizó a unos cuarenta metros de la roca.
Phineas Gage no murió a raíz de este terrible accidente. Los dos médicos que una media hora después lo atendieron en el pequeño pueblo situado a un kilómetro del sitio del accidente no podían creer que Phineas estuviera no solamente vivo, sino también consciente. Esa misma noche, unas 6 horas después, Phineas podía hablar y recordar con claridad todo lo que había sucedido, e inclusive decía que en pocos días estaría de nuevo trabajando. A pesar de que no había duda que la barreta había pasado a través de la parte más anterior de su cerebro, separando una parte del mismo del resto, su percepción del tiempo, sus sentidos y su lenguaje no habían sido afectados.
El estado de Phineas en los días siguientes fue muy grave. Además de la sangre que había perdido, la herida se infectó y estuvo muy cerca de morir. Sin embargo, su fuerte naturaleza y las altas dosis de los antisépticos de entonces —en 1848 no había antibióticos— le salvaron y dos meses después del accidente se sentía listo para empezar una nueva vida. La pregunta para nosotros es ¿cómo iba a ser esa nueva vida?, ¿en qué se iba a afectar —si es que algún cambio habría— la vida de un hombre que había perdido una parte de su cerebro, al ser esta parte desconectada del resto por el paso de la barreta? La respuesta a estas preguntas nos la da el testimonio de uno de los médicos que lo atendieron y que lo vio de nuevo años después. Este médico escribió:
Aunque su salud física parece haberse recuperado,
el equilibrio entre sus facultades intelectuales y sus tendencias más
animales parece haberse destruido. Es irreverente, agresivo, dado a
exclamaciones profanas y vulgares (lo cual no era su costumbre), manifestando
muy poca deferencia hacia sus compañeros, impaciente para realizar sus
deseos, obstinado y al mismo tiempo caprichoso y vacilante, haciendo
muchos planes para el futuro que más tardan en ser pensados que en ser
abandonados... En este aspecto su mente cambió radicalmente, tanto que
sus amigos y conocidos dicen de él que "ya no es Gage". |
Este cambio fue tan notorio que sus patrones no quisieron recontratarlo y Phineas se dedicó a vagar por el territorio de Estados Unidos y por Sudamérica, cargando consigo la barreta y exhibiéndose en ferias como un milagro viviente. Finalmente murió en San Francisco, y tanto la barreta como el cráneo de Phineas se recuperaron y en la actualidad pueden verse en el museo de la Escuela de Medicina de Harvard, en Massachusetts.
ZONAS DEL CEREBRO: ¿DISTINTAS FUNCIONES?
¿Qué interpretación puede darse a este caso tan excepcional de la vida real que parece inventado, como el del individuo que perdió todos sus sentidos y la capacidad de mover los músculos relatado al inicio de este libro? En cierto modo este caso verdadero es la contrapartida del narrado en el capítulo I: un individuo que conserva perfectamente todos sus sentidos y la capacidad íntegra de sus movimientos musculares, pero cambia su personalidad, ya no es el mismo, debido a una lesión del cerebro en la que el lóbulo frontal (llamado así por estar colocado en la parte anterior del cráneo sobre las órbitas de los ojos) fue separado de todas las otras partes del cerebro. Una conclusión apresurada pero razonable sería que ese lóbulo del cerebro es la región que determina la personalidad y el modo de ser de cada individuo, por lo cual al ser separado o destruido, dicha personalidad cambia radicalmente. Con esta idea en mente, un siglo después de este accidente, entre 1935 y 1950, se realizaron varios miles de operaciones quirúrgicas conocidas con el nombre de lobotomía frontal, en las cuales los neurocirujanos hacían prácticamente lo mismo que la barreta, es decir, desconectar el lóbulo frontal del resto del cerebro, sólo que en una sala de operaciones y con toda la asepsia necesaria para evitar infecciones. Paradójicamente, el objetivo de estas operaciones era precisamente cambiar la conducta de ciertos pacientes agresivos o con alteraciones mentales, para volverlos más "normales". El portugués Egas Moniz, un neuropsiquiatra, concibió esta idea al conocer los resultados de una lobotomía frontal realizada en un chimpacé que mostraba una conducta muy temperamental y difícil de manejar cuando algo no le salía bien al rebuscar dónde habían ocultado sus alimentos, conducta que desapareció después de la lobotomía. Sobre esta endeble base experimental, Moniz y sus colaboradores, y después otros neurocirujanos, realizaron muchas lobotomías frontales con resultados según ellos muy alentadores. Sin embargo, esta operación carente de fundamento, prácticamente ya no se realiza. Si se piensa en la historia de Phineas Gage, ciertamente no es fácil encontrar una justificación apropiada para que a tantas personas se les haya sometido a tan riesgosa mutilación cerebral.
Independientemente de esta equivocada concepción de los padecimientos mentales
y de la participación de los lóbulos frontales en ellos, el accidente de Phineas
Gage hizo ver que algunas regiones del cerebro podrían estar relacionadas con
cierta función particular, es decir, que parecería haber una especialización
interna en el cerebro mismo. Esta idea llevó a exageraciones casi irracionales
por parte de los llamados frenólogos de la segunda mitad del siglo XIX,
quienes en su afán de asignar funciones mentales específicas para cada zona
cerebral, llegaron a hablar de regiones que determinarían hasta las actitudes
políticas, como el "republicanismo". Por si fuera poco, los frenólogos afirmaban
que era posible determinar las tendencias, el modo de ser y las características
mentales de las personas analizando las protuberancias del cráneo y de la cara,
pues éstas reflejarían cuáles de las áreas cerebrales eran más grandes
o más importantes. Se relata, por ejemplo, el caso de una mujer particularmente
bien dotada para las relaciones morosas que tenía unas protuberancias craneales
excepcionalmente grandes detrás de los oídos, lo cual dio pie a los frenólogos
para postular que este tipo de protuberancias indicaba una habilidad especial
para el amor físico.
La frenología fue muriendo lentamente de muerte natural por carencia de evidencias científicas que la sustentaran, a pesar de que a principios de este siglo se llegaron a diseñar aparatos más o menos sofisticados para determinar qué partes del cráneo eran más prominentes y así establecer el modo de ser de las personas . Sin embargo, la muerte de la frenología causada por su misma exageración no destruyó la idea de que el cerebro está organizado en regiones diferentes, las cuales probablemente tengan que ver con ciertas funciones específicas. Experimentos en humanos llevados a cabo alrededor de 1950 por Jackson y Penfield demostraron que si se estimulaba la corteza cerebral eléctricamente era posible observar la contracción de un músculo o grupo de músculos preciso, dependiendo del sitio de la corteza que era estimulado. Se descubrió así que existe una zona especial de la corteza cerebral donde se localizan las motoneuronas responsables del movimiento de los músculos de las extremidades, los dedos, el tronco, la lengua, la cara, etc. Además, lo que fue todavía más interesante: se descubrió que la zona de la corteza que controlaba la actividad de los músculos con mayor movilidad o mejor control, como son los del pulgar o los de la lengua, tenía una superficie mucho más extensa que la de aquellos músculos de movilidad limitada y de importancia menor en las funciones motoras, como son los músculos del tronco, incluyendo el tórax y el abdomen. Se construyó así el llamado homúnculo motor que se puede observar en la Figura 30, en el cual el tamaño de cada región del cuerpo representa comparativamente qué tan grande es el área de la corteza cerebral que controla los músculos de dicha región corporal.
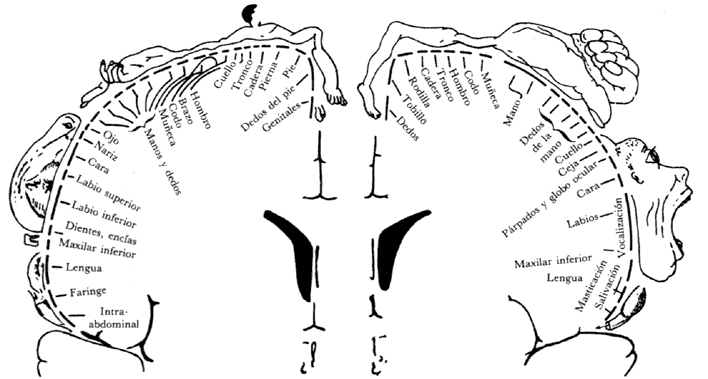
Figura 30. La llamada corteza cerebral motora (dibujo del lado derecho) en el lóbulo parietal de los hemisferios cerebrales, controla los movimientos de los músculos de lado contrario del cuerpo. Lo más interesante es que la superficie de la corteza que controla los músculos de movilidad más amplia y al mismo tiempo más fina y precisa, como los de la cara, boca, manos, pulgar y lengua, es mucho mayor que aquella del tronco, muslo y piernas. Esto es lo que está representado por el "homúnculo" dibujado en el extremo derecho. Lo mismo sucede con la corteza sensitiva (dibujo del lado izquierdo), a la cual llegan los estímulos táctiles de todo el cuerpo. En este caso, la superficie de la corteza más extensa corresponde a los sitios del cuerpo que poseen sensibilidad más exquisita, como labios, lengua, faringe, dedos de mano y pies, así como órganos genitales.
En experimentos paralelos, en los cuales se tocaban levemente con un alfiler o con objetos romos distintas partes del cuerpo y simultáneamente se registraba la actividad eléctrica de distintas zonas de la corteza cerebral, se descubrió que existían regiones en dicha corteza que recibían específicamente los estímulos sensoriales de la piel de cada zona del cuerpo, y de nuevo había una correlación entre la capacidad de sentir más finamente y la extensión de la corteza que recibía tales estímulos. Por ejemplo la lengua, que es uno de los órganos con más fina sensibilidad que el hombre posee (como es de experiencia de todos, basta el más fino cabello o pelusilla en la lengua para sentirla, lo cual no sucede ni siquiera con la yema de los dedos), así como los órganos sexuales, tienen un área muy extensa en la corteza que se encarga de recibir y procesar la llegada de estos estímulos sensoriales; en contraste, otras regiones como el tórax tienen una representación pequeña en la corteza sensitiva. La zona de la corteza sensitiva es diferente de aquélla de la corteza motora, estando una por detrás de la otra, situadas en la parte posterior del lóbulo frontal del cerebro.
CENTROS NEURONALES EN EL INTERIOR DEL CEREBRO
Además de la corteza cerebral, que como su nombre indica es la capa superficial que recubre los hemisferios cerebrales y que tiene muchos surcos que aumentan considerablemente su superficie, en el interior del cerebro existen grupos de neuronas bien definidos, separados de otros grupos por las fibras nerviosas constituidas por los grandes conjuntos de axones que van de un lado a otro del cerebro, y es lo que se denomina "materia blanca", en contraposición con los grupos de neuronas, que constituyen la llamada "materia gris". (Figura 31.) Algunos de estos grupos o núcleos neuronales del interior del cerebro también se han relacionado con aspectos específicos del funcionamiento del cerebro, por ejemplo el control de los movimientos musculares, particularmente de los movimientos finos, o inclusive de otras funciones que podríamos llamar más relacionadas con las actividades mentales. Por ejemplo, experimentos realizados en ratas a las cuales se les ha implantado un electrodo (varilla muy delgada de un metal que es capaz de conducir la electricidad) en una parte del interior del cerebro conocida como "sistema límbico", han demostrado claramente que si el electrodo está en cierto sitio particular, y éste se estimula por medio del paso de una pequeñísima corriente eléctrica, el animal muestra signos claros de ira, atacando prácticamente cualquier objeto que se acerque, vivo o no; en cambio si se estimula otra zona cercana, lo que sucede es que el animal se queda exageradamente tranquilo y no responde a estímulos que normalmente provocarían en él una reacción de defensa o de ataque.
Al ser estimuladas otras zonas del cerebro se producen, aparentemente, una muy intensa sensación placentera. Esto ha podido demostrarse en ratas a las que se les ha colocado un electrodo en una región del sistema límbico, diferente a las mencionadas arriba, de tal manera que el cable del electrodo esté conectado a un dispositivo que permite que la propia rata se estimule a sí misma en esa región del cerebro, si presiona una palanca con las patas. Lo que se ha observado en estos casos es que una vez que la rata se ha autoestimulado una vez, empieza a presionar la palanca en forma continua durante mucho tiempo, llegando inclusive al grado de dejar de comer por no interrumpir la autoestimulación. Aunque es claro que no podemos saber qué es lo que la rata siente al autoestimularse, es evidente que la sensación que obtiene debe ser extraordinariamente placentera, pues de otro modo no se concibe que continúe haciéndolo tan repetidamente hasta el grado de dejar de comer. De manera análoga pero contraria, al ser estimuladas otras regiones del sistema límbico producen una sensación que seguramente es muy desagradable, pues la rata una vez que ha apretado la palanca para autoestimularse no vuelve jamás a acercarse a ella. Se habla así de centros del cerebro relacionados con el placer o con el desagrado.
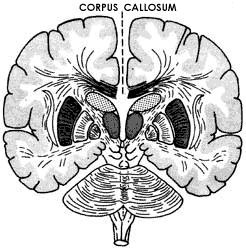
Figura 31. La materia gris cerebral está constituida por la corteza que recubre los dos hemisferios cerebrales y por los grupos de neuronas que están situados en su interior, mientras que la materia blanca está formada por los conjuntos de fibras nerviosas (axones) que viajan de una parte a otra del cerebro y a lo largo de la médula espinal. En este corte del cerebro se observa la corteza y algunos de los núcleos neuronales del interior. El corpus callosum o cuerpo calloso, es un gran conjunto de fibras que comunica los hemisferios cerebrales entre sí. En este dibujo se ve cortado, como se ha hecho en monos con fines experimentales y en humanos con fines terapéuticos (véase el capítulo VI).
Hay todavía interesantes ejemplos de centros cerebrales aparentemente responsables de otras funciones específicas. Una lesión o la destrucción de una zona particular conocida con el nombre de "hipotálamo", en la base del cerebro, hace que una rata empiece a comer descontrolada y vorazmente, sin saciarse aparentemente y volviéndose, en consecuencia, extraordinariamente obesa, por lo que se ha concluido que dicha zona corresponde al centro de la saciedad. De manera similar, se han descubierto otros centros como el de la sexualidad. Cuando éste se estimula, el animal se vuelve hiperactivo sexualmente, y si es macho tenderá a montar no sólo a las hembras que se encuentren cerca, sino también a los machos o inclusive a otras especies animales diferentes a la suya (estos experimentos fueron hechos en gatos); más todavía, un animal así estimulado tenderá a mostrar una conducta sexual aún frente a ciertos objetos inanimados, por ejemplo animales de peluche.
CIRCUITOS NEURONALES Y LA FUNCIÓN DE LAS REGIONES CEREBRALES
Los ejemplos que acabamos de citar en los párrafos anteriores, indican con claridad que, sin llegar a las absurdas exageraciones de los frenólogos, es claro que muchas regiones del cerebro tienen que ver específicamente con una función más o menos bien definida, tanto en lo que se refiere a sus aspectos motores como en aquellos más relacionados con las funciones mentales. Podríamos todavía mencionar otras áreas con una función muy precisa, como es la región del lenguaje oral, localizada en una región pequeña del lóbulo temporal izquierdo. Es ésta la región que cuando se destruye ocasiona la pérdida de la capacidad para expresarse verbalmente.
¿Qué representan en términos de conexiones neuronales los distintos grupos de neuronas que parecen estar relacionados con funciones específicas? Si hemos insistido en los capítulos anteriores que las funciones del sistema nervioso dependen básicamente de la comunicación entre las células nerviosas, ¿cuál es la participación de los mecanismos de comunicación interneuronal mediante moléculas neurotransmisoras en la realización de las funciones en que participan los diferentes grupos de neuronas? Podremos dar sólo algunas respuestas parciales a esta importante e interesante pregunta. En primer lugar, es claro que si un núcleo neuronal como los que hemos mencionado se aísla de los demás, difícilmente podría producir las acciones descritas. Es sólo mediante las conexiones de cada uno de esos núcleos con otras regiones del cerebro que su función se puede manifestar. Y si ahora pensamos cómo tales conexiones entre los grupos neuronales se establecen, recordando la forma y organización de las neuronas, será fácil concluir que los axones de las neuronas de un determinado núcleo se dirigirán precisamente a los otros núcleos que tengan que ver también con la actividad o función de que se trate, y éstos a su vez harán lo mismo con otros, tal como ya habíamos mencionado en el capítulo anterior. El resultado de estas comunicaciones entre grupos neuronales es la formación de circuitos o redes de neuronas que en conjunto llevan a cabo un determinado aspecto de las funciones cerebrales.
Veamos ahora un poco más de cerca que papel tienen los neurotransmisores excitadores e inhibidores en el funcionamiento de los circuitos. Hemos visto en capítulos anteriores que las neuronas se comunican entre sí mediante mecanismos químicos que implican el uso de moléculas mensajeras que llevan la información de excitación o de inhibición a la siguiente neurona. Cuando se trata de circuitos formados por muchas neuronas es claro que el funcionamiento del circuito neuronal completo dependerá en último análisis de la manera en que los diferentes neurotransmisores excitadores e inhibidores que participan en cada una de las conexiones del circuito esté funcionando en un momento dado.
Teniendo en mente lo anterior, podemos imaginar el funcionamiento del circuito si pensamos que éste opera mediante la integración de las entradas y las salidas. Por ejemplo, si al circuito en cuestión llegan varias entradas provenientes de neuronas de tres o cuatro sitios o regiones distintas del cerebro y todos estos axones son excitadores, esa información tendrá que ser procesada como una suma de estímulos que llegan de distintos sitios, de tal modo que la salida del circuito estará activada y éste enviará su señal hacia otro u otros circuitos o núcleos neuronales.
Hasta aquí no habría gran diferencia entre una sola neurona y un grupo de ellas en un circuito, excepto en que se estarían sumando las entradas, mediante el proceso que se conoce como suma espacial o topográfica, ya que dichas entradas provienen de regiones diferentes. Sin embargo, si recordamos la existencia de las neuronas inhibidoras y nos imaginamos que dentro de la organización del circuito puede haber una o varias neuronas de este tipo, las cuales pueden inhibir a muchas de las neuronas excitadoras del propio circuito, el panorama cambiará de manera importante. En efecto, por más que puedan llegar al circuito muchos estímulos excitadores provenientes de otros núcleos neuronales, tales excitaciones serán matizadas o controladas por las neuronas inhibidoras que forman parte del mismo circuito. Como se observa en la Figura 32, esta posibilidad de organización permite que un determinado circuito tenga muy interesantes posibilidades de autorregulación. Así, cabe la posibilidad de que una neurona cuyo axón emerja del circuito tenga una colateral que se comunique con una neurona inhibidora, la cual a su vez va a inhibir a otra u otras neuronas excitadoras del propio circuito. De esta manera, las posibilidades de procesamiento o manejo de la información que llega a cada circuito neuronal son múltiples y pueden graduarse de acuerdo a las necesidades o condiciones de funcionamiento del circuito. El ejemplo de la figura 32 nos ayudará a entender este mecanismo. El axón de la motoneurona que emerge de la médula espinal para dirigirse a un músculo y excitarlo —en este caso hacer que se contraiga—, emite una colateral cuando todavía está dentro de la médula, la cual se comunica con una interneurona de axón corto, llamada por esta razón interneurona, la cual es inhibidora de la propia motoneurona que había emitido la colateral. Es claro que el resultado de la existencia de este tipo de arreglos entre neuronas excitadoras e inhibidoras lleva a la posibilidad de controlar a una neurona excitadora, de tal manera que ésta tenga limitada su acción y sea más difícil caer en un estado de hiperexcitación que evidentemente sería peligroso para el músculo y por consiguiente para el organismo en su conjunto.
Figura 32. Los mecanismos de inhibición permiten la regulación de la contracción de los músculos agonistas y antagonistas. Cuando el músculo flexor es excitado, como en la figura 3, la neurona sensitiva excita a la motoneurona que hará contraer a ese músculo, pero al mismo tiempo, mediante una colateral, excita a una neurona inhibidora (en negro) de la médula espinal, la cual inhibirá la motoneurona del músculo antagonista extensor. Además el axón de la motoneurona del músculo flexor emite una colateral que excita a otra neurona inhibidora (también de negro) que a su vez inhibirá asimismo la motoneurona del músculo antagonista y a la propia motoneurona del flexor y de su agonista, proporcionando así un mecanismo regulador que permite una contracción más controlada de todos los músculos que participan en la movilidad de la articulación.
Vayamos ahora un poco más allá, intentando ver qué sucede cuando la salida de un circuito afecta a otro circuito. Regresemos a nuestro ejemplo de la motoneurona que sale de la médula espinal y cuyo colateral del axón excita a una interneurona inhibidora. ¿Qué pasaría si el axón de esta última neurona inhibiera la motoneurona de otro músculo? A primera vista parecería que este tipo de arreglo —ya descrito en el capítulo III—, no podría tener ninguna utilidad, ya que daría la impresión de interferir con otro sistema neuronal independiente, lo cual no suena razonable. Sin embargo, si pensamos que esta segunda neurona que sería inhibida como consecuencia de la excitación de la motoneurona inicial, podría ser la que se encargara de excitar precisamente al músculo antagonista que resultará excitado por la primera neurona del sistema, nos daríamos cuenta que éste sería un mecanismo muy útil y eficiente para permitir que el músculo se contraiga. En efecto, mediante este arreglo de excitación e inhibición se excita un músculo al mismo tiempo que se inhibe a su antagonista, con lo cual la oposición entre ambos queda anulada (véase la Figura 32).
Los ejemplos anteriores dan una idea clara de cómo dentro de un mismo circuito los mecanismos de inhibición permiten un control fino de su funcionamiento global; además, estos mismos mecanismos por inhibición son capaces de regular el funcionamiento de un circuito neuronal como resultado de la actividad de otro circuito.
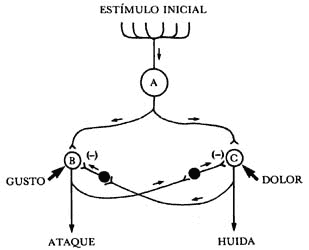
Figura 33. Los circuitos neuronales con neuronas inhibidoras intercaladas proporcionan eficientes mecanismos reguladores de las respuestas a estímulos externos. En este esquema, un estímulo inicial captado por la neurona A puede causar inicialmente una respuesta de ataque a causa del estímulo, mediante la excitación de la neurona B. Si como consecuencia del ataque se produce una sensación de "gusto", se reforzará dicha respuesta de ataque, mientras que simultáneamente se inhibe la posible respuesta de huida mediante la neurona inhibidora que actúa sobre la neurona C. Por el contrario, si la consecuencia de ataque es dolor, registrado por la neurona C, ésta generará una respuesta de huida al tiempo que mediante la colateral de su axón y la correspondiente neurona inhibidora, inhibe la neurona B. Las neuronas inhibidoras son las negras, y su acción esta señalada en la sinapsis correspondiente con el signo (-). Todas las otras neuronas y sus correspondientes sinapsis son excitadoras. Las flechas señalan la dirección de la información a lo largo de los axones.
Imaginemos ahora el circuito hipotético de solamente cinco neuronas que se observa en la Figura 33. En este circuito se ha sobresimplificado muchas cosas, entre ellas los sistemas de llegada de estímulos dolorosos ("dolor") o de gusto ("gusto"), así como las salidas, marcada como "ataque" o "huida". Haciendo una analogía con los circuitos que controlan la contracción de los músculos y sus antagonistas —en los que ya vimos que si no se inhibe el músculo antagónico es difícil que se contraiga aquél que se quiere contraer— de manera similar, si no se inhibe el circuito que determina la huida, no se podrá poner en marcha el circuito que origina el ataque. Nuevamente, y siguiendo con la analogía de los mecanismos, es necesario que exista una neurona inhibidora capaz de inhibir el circuito antagónico a aquél que se desea echar a andar. Naturalmente, para que esta neurona inhibidora realice su función acoplada entre los dos circuitos, se requiere que la colateral de cada uno de ellos excite las neuronas inhibidoras del mecanismo contrario al mismo tiempo que se activa o excita el que debe predominar (véase la Figura).
Es claro que la existencia de neuronas inhibidoras dentro de los circuitos neuronales conlleva la posibilidad de regular, controlar o modular el funcionamiento de ellos y, lo que es quizá más importante, permite la posibilidad también de modular la actividad de otros circuitos neuronales opuestos, al tiempo que se activa el requerido.
NEUROTRANSMISORES Y FUNCIONES CEREBRALES: EXPERIMENTOS CON DROGAS
Regresemos a los mecanismos químicos de la comunicación entre las neuronas, revisados en el capítulo anterior, pero ahora tratemos de analizarlos en términos de circuitos neuronales. Para este propósito describiremos algunos experimentos que permiten algunas conclusiones interesantes respecto a cómo funcionan algunos de los circuitos neuronales que tienen que ver con el control de los movimientos voluntarios. Además, en el animal íntegro, estos experimentos permiten ver con cierta claridad el papel de algunas neuronas inhibidoras en el funcionamiento de los circuitos mencionados.
Hagamos inicialmente algunas consideraciones respecto a cómo podría ser estudiada la comunicación interneuronal y además, cómo sería posible distinguir entre la actividad de la neuronas excitadoras y las inhibidoras en el animal vivo. Pregunta difícil para adentrarnos en su respuesta y así poder referirnos a los experimentos anunciados; damos por sentado que estamos hablando de comunicación química y no eléctrica, entre neuronas; comunicación que es la más importante y abundante en cerebro y médula espinal de vertebrados, particularmente de los mamíferos a los cuales el hombre pertenece. Recordemos que por definición la comunicación química implica que la neurona a comunicar con la siguiente, libere un neurotransmisor al espacio que la separa de ella y que dicho neurotransmisor se combine específicamente con la molécula receptora localizada en la membrana de la segunda neurona. Considerando lo anterior es fácil visualizar que mediante manipulaciones de la función de los neurotransmisores sea posible alterar experimentalmente el funcionamiento de alguno o algunos de los pasos en la comunicación interneuronal. Por ejemplo, si cada neurotransmisor debe ser sintetizado, como hemos dicho, por la terminal del axón en el sitio en que éste se comunica con la otra neurona, ¿habrá algún modo de impedir que se forme dicho transmisor sin afectar la síntesis de otros transmisores? Debido a la manera como se llevan a cabo las reacciones bioquímicas dentro de las células, las cuales requieren, como hemos dicho en el capítulo II, que ciertas proteínas con propiedades catalíticas participen, la respuesta es: sí, mediante el uso de ciertas drogas o fármacos, se puede "engañar" a la proteína que sirve para formar el neurotransmisor, y así impedir que éste se sintetice. Como este tipo de proteínas posee esa extraordinaria propiedad de la especificidad o selectividad, sobre la cual insistimos mucho en el capítulo II, es posible usar drogas que impidan la síntesis de un neurotransmisor sólo, sin afectar a otros neurotransmisores.
Con esta información, imaginemos ahora que disponemos de una droga capaz de impedir la síntesis de un neurotransmisor inhibidor, pero no sabemos qué es lo que éste hace primordialmente en los complicados circuitos neuronales del cerebro. Una manera de saberlo sería inyectar la droga en cuestión al animal y observar qué efectos tiene sobre su conducta. Desde luego que estamos simplificando el problema y dando por sentado que la droga inyectada efectivamente impide la síntesis del transmisor y que además puede penetrar al cerebro por la sangre cuando se inyecta. Tomemos pues un ratón albino, animales fácilmente manejables y que se usan rutinariamente en los laboratorios de todo el mundo, e inyectémosle nuestra droga. Coloquemos al ratón en una jaula transparente para observarlo, sentémonos en un banco cómodo, con lápiz y papel o una videograbadora si disponemos de ella y veamos qué le pasa al ratón. Recordando que el transmisor cuya síntesis estamos impidiendo con la droga inyectada es un transmisor inhibidor, veremos que al cabo de unos 30 a 45 minutos, el ratón se muestra inquieto, se rasca continuamente la cara, se limpia los bigotes, se acicala la parte más alta de la cabeza, brincotea un poco y después de varios minutos con esta conducta, de pronto corre muy rápidamente golpeándose con las paredes de la jaula durante 5 o 10 segundos, después, pone todos sus músculos muy rígidos en una contracción generalizada de toda la musculatura. Esta contracción total, muy intensa, puede terminar con la vida del ratón si los músculos respiratorios del tórax y el diafragma se ven también afectados, pues no podría seguir respirando. Si esto último no sucede, el ratón se recuperará lentamente, pero unos cuantos minutos más tarde volverá a caer en ese estado de contracción generalizada.
La conducta motora que acabamos de describir es muy parecida a ciertos tipos de convulsiones de tipo epiléptico en el hombre, por lo que puede afirmarse que en el experimento descrito estamos produciendo en el ratón epilepsia experimental. Si recordamos ahora lo que originó el experimento, el hecho de que la droga que inyectamos impide la formación de un transmisor inhibidor, la conclusión de este resultado experimental debe ser que muy probablemente la función de ese transmisor es participar como inhibidor en los circuitos neuronales que tienen que ver con el inicio de los estímulos que finalmente van a producir la contracción de los músculos. Al inyectar la droga e impedir que el transmisor se sintetice por la neurona inhibidora, aquellas neuronas que forman el extraño homúnculo que aparece considerando la superficie de la corteza cerebral destinada a los diferentes músculos—, y por consiguiente éstas disparan en forma no modulada, descontrolada y sobrevienen las convulsiones.
Naturalmente que esta conclusión no se puede considerar como definitiva si no está apoyada en otros hechos experimentales que la confirmen. Entre los varios experimentos que se han diseñado para saber si en verdad la conclusión a que hemos llegado es correcta, se ha inyectado otra droga cuya acción no es impedir la síntesis del transmisor inhibidor, sino antagonizar la combinación de éste con su receptor en la membrana de la neurona motora. El resultado de inyectar esta droga es muy similar al de impedir la síntesis del transmisor, sólo que los efectos son ahora más rápidos. En este caso el ratón tiene convulsiones sólo 5 o 10 minutos después de la inyección y estas convulsiones son muy parecidas a las observadas con la otra droga. La hipótesis de que el neurotransmisor en cuestión es un regulador natural de los circuitos cerebrales que tienen que ver con el inicio de la contracción de los músculos se ve así fuertemente apoyada. Era de esperarse que la acción de esta nueva droga fuera similar a la de la inhibición de la síntesis del transmisor, ya que si éste no puede combinarse con su receptor es como si su acción hubiera disminuido mucho o como si no existiera, pues mientras no actúe sobre la molécula receptora de la siguiente neurona, nada sucede desde el punto de vista de la comunicación entre las dos neuronas. En ambos casos se logra enmudecer la comunicación: la primera neurona no puede hablar cuando se impide que forme su transmisor o bien la segunda no puede oír, porque se ha tapado su receptor. De este modo, mediante este tipo de experimentos, se ha podido conocer que existe un neurotransmisor inhibidor cuya acción es precisamente la de regular, o mantener bajo estricto control, la actividad de las neuronas excitadoras de cuya acción depende la contracción de los músculos. Dichas neuronas se encuentran localizadas en los sitios de la corteza motora, la cual constituye parte fundamental de los circuitos de excitación muscular.
MÁS EXPERIMENTOS CON DROGAS: ACCIÓN SOBRE LA COMUNICACIÓN NEUROMUSCULAR
Los experimentos relatados nos muestran la enorme utilidad del buen uso de las drogas como herramienta para ver qué está sucediendo en ciertos circuitos neuronales y cuáles son los neurotransmisores con los que éstos trabajan. Gracias a drogas capaces de alterar el funcionamiento del cerebro, hemos podido conocer la naturaleza de muchas moléculas neurotransmisores en distintas regiones del sistema nervioso y podemos tener cierta idea de qué función llevan a cabo —inhibidora o excitadora— en dichos circuitos. Mediante su uso podemos no sólo impedir la síntesis de neurotransmisor o la combinación con su receptor en la membrana de la siguiente neurona, sino también evitar que el transmisor se libere desde la terminal axónica o bien aumentar su liberación. En el laboratorio del autor se han utilizado algunas de estas drogas que evitan o aumentan la liberación de los neutransmisores con resultados interesantes para la discusión de este capítulo. ¿Qué sucede cuando a un animal se le inyecta una droga que inhibe la liberación del transmisor? Pensemos, inicialmente, no en el cerebro sino en la comunicación entre las terminales nerviosas de las neuronas motoras sobre los músculos, es decir, en la comunicación neuromuscular. Recordemos que cuando estas neuronas —localizadas en la médula espinal— se excitan, liberan desde sus terminales vecinas al músculo un transmisor excitador que actúa sobre la membrana de las células musculares y como consecuencia el músculo se contrae. Si inyectarnos a un ratón la droga mencionada, a una dosis que pueda llegar hasta los sitios de comunicación entre los nervios y los músculos, la droga impedirá que el transmisor sea liberado y por consiguiente los músculos ya no podrán contraerse por falta de esta comunicación excitadora. El resultado será que el ratón no pueda moverse, caerá en un estado de parálisis con los músculos fláccidos, tal como se ve en la fotografía. (Figura 34.)

Figura 34. Este ratón está totalmente paralizado por la inyección de una sustancia que inhibe la liberación del transmisor que excita los músculos. Este experimento fue realizado en el laboratorio del autor, como se describe en el texto. Ningún ratón no paralizado permite ser sostenido de la piel del cuello, pues rápidamente se voltearía para morder a quien lo intenta.
¿Cómo comprobar que la droga inyectada está efectivamente actuando mediante la inhibición de la liberación del neurotransmisor? Una manera más o menos obvia es inyectar al ratón paralizado otra droga que tenga el efecto contrario, es decir, que estimule la liberación del transmisor. Afortunadamente también existen este tipo de drogas. El experimento siguiente al de la producción de la parálisis con la primera droga fue tomar al animal paralizado e inyectarle la segunda droga. El resultado fue emocionante: a los dos o tres minutos de la segunda inyección, los animales empezaron a mover ligeramente las patas y, en otros dos o tres minutos, eran capaces de caminar casi normalmente. Lo que hicimos en estos experimentos fue entonces antagonizar el efecto paralizante de la primera droga con la segunda, confirmando así que la parálisis se debía a la disminución de la liberación del transmisor, de modo que al incrementar dicha liberación por la segunda droga, los animales podían de nuevo moverse normalmente.
Los experimentos que acabamos de relatar nos muestran con gran claridad que el control de la intensidad de los movimientos musculares se ejerce por la cantidad de transmisor que es liberado en cada momento desde las terminales de los nervios, es decir, de los axones de las neuronas excitadoras cuyo soma se encuentra en la médula espinal. En la vida real existen numerosos ejemplos de la acción de drogas usadas como armas, que matan precisamente por alterar los mecanismos químicos de la comunicación nerviosa. Por ejemplo, algunos gases usados durante la segunda Guerra Mundial actúan impidiendo las reacciones normales de interconversión del neurotransmisor activo sobre los músculos. El curare, veneno ancestralmente usado por algunas tribus del Amazonas, con el cual impregnan la punta de sus flechas, es una droga que se combina con los receptores al transmisor en los músculos y por los tanto éstos no pueden contraerse. Este veneno es extraordinariamente potente y basta una pequeñísima cantidad para que los músculos respiratorios se paralicen sobreviniendo la muerte.
Los venenos de algunas serpientes, así como el de la araña conocida como viuda negra, contienen toxinas que al parecer también actúan sobre los mecanismos de liberación de transmisores, bien porque impiden dicha liberación o porque producen una liberación masiva de ellos. No es difícil imaginar que, si estos venenos llegan hasta el cerebro, afecten el funcionamiento de varios de los circuitos neuronales. Debemos aquí recordar también que muchos de los circuitos neuronales son los que controlan las llamadas funciones involuntarias del organismo a que nos referimos en el capítulo I, tales como la contracción del corazón o la respiración. Es evidente entonces que si estos venenos afectan, como muchas veces ocurre, los mecanismos de transmisión entre las neuronas de este tipo de circuitos, la muerte puede sobrevenir muy rápidamente.
LA REGULACIÓN FINA DE LOS MOVIMIENTOS Y LA ENFERMEDAD DE PARKINSON
Volvamos un poco más sobre los circuitos que controlan los movimientos musculares. Hemos dado ejemplos de experimentos en los que la movilidad muscular se altera en los dos sentidos: en una hiperactividad que resulta en convulsiones, o en una hipoactividad que resulta en parálisis. Estos ejemplos, sin embargo, se refieren sólo a acciones sobre los músculos mismos o sobre los circuitos que en forma directa inician la estimulación que finalmente provoca que el músculo se contraiga. Además de estos circuitos, en el interior del cerebro existen otros que se encargan de establecer una regulación muy fina de los movimientos. Con regulación de movimientos finos quiero decir aquélla que es necesaria para realizar las tareas de todos los días y que precisamente por ser cosas rutinarias no nos percatamos del increíble y exquisito control que requieren. Me refiero por ejemplo a tomar un vaso lleno de agua, llevarlo a los labios y beber el contenido sin derramar una gota, o servirse del cuchillo y tenedor para cortar y comer la carne, o pelar una naranja, o escribir, o caminar a una velocidad perfectamente controlada de acuerdo con la prisa que se tenga, y con el tipo de piso sobre el que se esté, librando los obstáculos que surjan, etc. Prácticamente cualquier actividad en que podamos pensar se podría usar como ejemplo de esta indispensable finísima regulación de nuestros movimientos. Por supuesto ésta es mucho más obvia si consideramos, ya no las actividades diarias que todo el mundo realiza todos los días, sino otras que requieren de gran destreza, entrenamiento, aprendizaje y finura. Los ejecutantes de instrumentos musicales son quizá el mejor ejemplo de esto. Los movimientos de los dedos de un pianista, un guitarrista o un arpista son asombrosos por su precisión, no solamente en cuanto a tocar las teclas debidas o pulsar las cuerdas adecuadas sin equivocarse, sino también por la enorme velocidad con que los movimientos se realizan y la variedad en la intensidad, calidad de la vibración y simultaneidad de las notas. La danza es también otro ejemplo maravilloso ya que en esta actividad son todos los músculos del cuerpo los que participan armónicamente del movimiento, lográndose un control exquisito de ellos. Otras actividades, menos artísticas que las mencionadas pero también interesantes, requieren asimismo de un grado de control muscular extraordinario. Los acróbatas, malabaristas, prestidigitadores, escaladores de rocas, etc., nos dejan asombrados por el control que son capaces de tener sobre sus movimientos musculares.
Como lo pintó en palabras el poeta Luis Rius, refíriéndose a la bailarina Pilar Rioja:
| Podría bailar en un tablado de agua sin que su pie la turbase, sin que lastimara al agua. No en el aire, que al fin es humano el ángel que baila. No, en el aire no podría, pero sí en el agua. |
¿Qué circuitos cerebrales son los que participan en tan fina regulación de los movimientos? En una de las regiones más internas del cerebro se encuentran dos grupos de neuronas conectados entre sí por sus axones, de tal manera que los axones de uno de los grupos llega hasta el otro y a su vez los axones de éste regresan a los del primero. (Figura 35). Este circuito es en gran medida, aunque no únicamente, el responsable del mencionado control de los movimientos musculares. Hace apenas cuatro años se descubrió, en otro de esos hallazgos fortuitos que al seguirse más profundamente proporcionan información de mucho valor, una sustancia muy tóxica que al ser inyectada a monos producía alteraciones del control de los movimientos. Estudios experimentales en varias especies animales demostraron que esta droga tenía estos efectos debido a que producía una lesión permanente de las neuronas de uno de estos dos grupos neuronales que participan en el circuito mencionado. Como resultado de esta lesión, esas neuronas mueren y por consiguiente ya no pueden liberar su neurotransmisor y actuar sobre las neuronas del segundo grupo constituyente del circuito. Los síntomas que mostraban los monos así lesionados eran muy parecidos a los de la enfermedad de Parkinson, la cual se conoce desde hace muchos años en humanos y se caracteriza porque los músculos están rígidos y tienen un temblor fino que, a veces se exacerba precisamente, cuando el paciente trata de realizar algún movimiento delicado. De hecho, se sabe que esta droga es capaz de producir la enfermedad de Parkinson en humanos intoxicados con ella —en realidad así fue como se descubrió, cuando se observó que aparecieron algunos casos de enfermedad de Parkinson en jóvenes menores de 30 años, cuando es bien conocido que esta enfermedad ataca a personas casi siempre mayores de 45 o 50 años—.
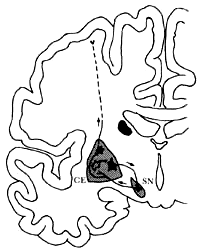
Figura 35. El control y la coordinación de los movimientos musculares se ejercen principalmente mediante la actividad del circuito neuronal mostrado en este esquema de un corte del cerebro. El grupo de neuronas del tallo cerebral conocido como sustancia negra (SN) inhibe a otro grupo de neuronas localizado en el interior del cerebro, llamado cuerpo estriado (CE), el cual a su vez inhibe a la sustancia negra. Esta última se comunica también con neuronas que a través de varias conexiones (no mostradas pero esquematizadas por línea discontinua) actúan sobre las motoneuronas de la corteza cerebral. Las neuronas negras son inhibidoras, y la otra es excitadora. Aquí sólo se muestra el circuito de un hemisferio cerebral, pero en el otro hemisferio también existe. Las alteraciones de los movimientos que se observan en la enfermedad de Parkinson —fundamentalmente rigidez, temblores y falta de movilidad— se deben a que neuronas de la sustancia negra se destruyen. Aún no sabemos la causa de esta destrucción, pero experimentalmente es posible reproducir los síntomas de la enfermedad mediante ciertas drogas que lesionan selectivamente estas neuronas de la sustancia negra. La información que se obtenga de este tipo de experimentos quizá proporcione la clave para entender por qué ocurre el Parkinson.
Unos 10 años antes del descubrimiento de esta droga tan tóxica se sabía que la enfermedad de Parkinson se debe a la destrucción del núcleo neuronal al que nos hemos referido. Este descubrimiento se había hecho mediante un estudio bioquímico sobre cuáles eran los posibles neurotransmisores que se afectaban en distintas regiones del cerebro de pacientes que morían con esta enfermedad. Sin embargo, hasta la fecha no se sabe qué es lo que causa que dichas neuronas mueran, por lo que el descubrimiento de la droga mencionada ha despertado mucho interés entre los investigadores ya que, precisamente por ser tan selectiva en la localización de las neuronas que mata, podría proporcionar la clave de cómo y por que mueren tales neuronas en la enfermedad de Parkinson.
Podemos concluir que la enfermedad de Parkinson es una enfermedad de un circuito neuronal, en la cual ciertas neuronas mueren, mientras que el resto del cerebro está normal. Es por esto que los tratamientos para la enfermedad, intentan sustituir al neurotransmisor que falta —debido a la lesión celular— Nuevamente enfatizamos que para conocer todo esto, primero fue necesario conocer los mecanismos de comunicación interneuronal que nos han ocupado buena parte de este libro. Sin estos conocimientos nunca hubieran podido buscarse los defectos bioquímicos que abrieron la puerta al descubrimiento del defecto en el neurotransmisor del grupo neuronal afectado.
¿Quiere todo esto decir que será posible identificar una lesión o defecto funcional en un circuito específico para cada enfermedad que afecta al sistema nervioso? Contestar esta pregunta no es fácil. En principio, parece posible que algunos de los padecimientos neurológicos que afectan los movimientos musculares, como la epilepsia, sí pudieran tener su explicación en lesiones más o menos específicas de ciertos circuitos cerebrales. Los ejemplos dados al inicio de este capítulo sobre las funciones específicas de varios de ellos podrían hacer pensar que también algunas alteraciones de las funciones mentales propiamente dichas podrían ser el resultado de lesiones restringidas a ciertos grupos neuronales. De hecho, ésta fue la base para las operaciones de lobotomía frontal en pacientes con alteraciones mentales anteriormente mencionados. Sin embargo, cada vez es más claro que no es posible pensar en explicaciones simplistas de este tipo para entender la actividad mental y sus alteraciones patológicas, pues aún estamos lejos de entender estas funciones en términos de circuitos neuronales. Baste por ahora decir que los fenómenos de comunicación interneuronal y particularmente los mecanismos químicos responsables de esta comunicación, tienen sin duda mucho que ver en un gran número de padecimientos neurológicos y mentales. En este sentido, la frase del investigador de finales del siglo pasado, Thudichum, podría ser profética. Thudichum escribió en 1884 en su Tratado sobre la constitución química del cerebro lo siguiente:
Creo que se demostrará que las grandes enfermedades
del cerebro y la médula espinal están relacionadas con cambios químicos
específicos en las neuronas... En resumen, es probable que con la química
muchas alteraciones del cerebro y de la mente, que actualmente son oscuras,
podrán ser definidas con exactitud y ser susceptibles de un tratamiento
preciso, y lo que es ahora el objeto de un ansioso empiricismo se convertirá
en el orgulloso ejercicio de las ciencias exactas. |
Es claro que aún falta mucho tiempo y muchas investigaciones para que estas
palabras de hace 100 años se hagan una realidad. ¿Hasta qué punto esto ocurrirá?
Es muy difícil de decir. En el próximo capítulo cerraremos este discurso en
el punto del futuro de la investigación sobre las células de la mente.