IV. LOS MECANISMOS DE AGRESIÓN
LA CAPACIDAD de un microorganismo para producir enfermedad en el ser humano depende de tres factores primordiales. En primer lugar, el parásito debe propagarse de un huésped a otro; es ésta la propiedad de comunicabilidad. Debe, en segundo término, tener la habilidad de invadir el organismo huésped; ello implica la presencia de un conjunto de factores que determinan la penetración del parásito, la resistencia de éste a los mecanismos de defensa del huésped, así como la posibilidad de sobrevivir y multiplicarse en el seno de los tejidos invadidos. El tercero de los factores requeridos para que un agente produzca enfermedad es su capacidad de dañar los tejidos del organismo que invade. No basta, pues, que un microorganismo produzca daño; debe contar con un medio eficaz para propagarse de un huésped a otro; debe poder colonizar, invadir y ser inmune a la respuesta inmune del huésped; encontrar en él los medios requeridos para su supervivencia y, por último, producir lesión en el huésped.
La notoria capacidad de la Entamoeba histolytica de producir daño extenso
en los tejidos del ser humano durante el transcurso de la amibiasis invasora
ha sido bien conocida por largo tiempo. Recordemos tan sólo las vastas lesiones
encontradas en la colitis amibiana fulminante, en las que casi la totalidad
de la mucosa del intestino grueso se encuentra alterada, o bien los grandes
abscesos hepáticos amibianos, que llegan a destruir buena parte del órgano (Figuras
16 y 17).
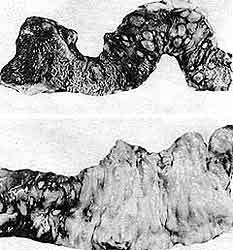
Figura 16. Lesiones intestinales producidas en humanos por la Entamoeba histolytica. (Cortesía del doctor Ruy Pérez Tamayo.)
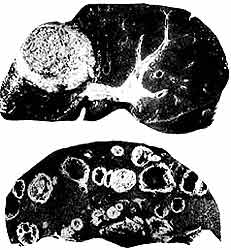
Figura 17. Abscesos hepáticos amibianos. (Cortesía del doctor Ruy Pérez Tamayo.)
Por algo, según vimos en el primer capitulo, Schaudinn decidió llamar a la amiba histolítica, es decir, productora de lisis de tejidos; el gran acierto de la denominación de Schaudinn ha hecho que desde 1903 esta amiba "en su nombre lleve la fama".
Fue esto, sin embargo, lo único que durante largo tiempo hicieron los investigadores en relación al problema de determinar la capacidad patógena de la amiba: adjudicarle un nombre descriptivo.
Tal vez sea el conocimiento de los mecanismos de agresión de la amiba histolítica el campo más explorado de la amibiasis experimental durante los últimos diez años. El tema interesa por igual a los biólogos celulares, quienes indagan las moléculas y los procesos celulares involucrados en el efecto citopático, a los inmunólogos, atraídos por el reto de descubrir los medios de que se valen las amibas patógenas para burlar las defensas del huésped, a los patólogos, que analizan en animales de experimentación o en material de biopsias y necropsias de casos de amibiasis invasora, las complejas interacciones celulares que dan por resultado la necrosis de un tejido invadido por las amibas. Del concurso de todas las observaciones ha emergido en años recientes una idea, cada vez más precisa, de la complejidad del fenómeno de la patogenia de la amibiasis.
Durante casi medio siglo, el conocimiento de la patogenia de la amibiasis se estancó, adormilado por falsos dogmas que satisfacían la escasa curiosidad de contados investigadores interesados en esta "Cenicienta" de las enfermedades parasitarias. Se aceptaba, sin discusión, la capacidad de las amibas de destruir al huésped infectado en la ausencia de respuesta inmune: no hay inflamación, se decía; "las amibas liberan enzimas —no importaba mucho cuáles, ni el hecho de que no hubieran demostrado— capaces de destruir los tejidos que invaden; al mismo tiempo se multiplican y cada generación de amibas resultante renueva la necesidad nunca satisfecha de alimentarse a expensas del huésped".
¿Cómo era posible que un microorganismo, sin duda poseedor de antígenos diferentes a los del organismo humano, no despertara reacción inflamatoria alguna? ¿Cuáles son las supuestas enzimas, que lo mismo destruyen tejidos epiteliales que armazones conjuntivos? ¿Cómo vence la frágil amiba la formidable muralla de la mucosa intestinal y la compleja estrategia defensiva de células y moléculas encargadas de la inmunidad local? ¿Por qué es sólo el hombre y no otros mamíferos, víctima frecuente de la actividad agresora de la amiba?
Estas y muchas otras preguntas de interés para el conocimiento de la amibiasis invasora languidecieron durante décadas. Sin duda, el hecho más importante, responsable del resurgimiento de dicho interés, fue la obtención de un procedimiento para el cultivo de amibas libres de bacterias ideado por Diamond en 1961 (Figura 18) que permitió analizar el parásito sin ningún otro microorganismo asociado y emplearlo en intentos de reproducir la enfermedad humana en animales de experimentación. Hasta entonces, el experimentador se veía obligado a emplear muestras que, más que amibas, contenían, sobre todo, sinnúmero de bacterias indefinidas y gran cantidad de partículas, como arroz o glóbulos rojos, requeridas para satisfacer el voraz apetito de los minúsculos predadores.

Estas mezclas malolientes eran inyectadas en el intestino o en el hígado de sufridos roedores, al cabo de semanas o meses se extraían los órganos afectados para que, a través del examen histopatológico de esas lesiones, el experimentador diera rienda suelta a su imaginación y describiera, a partir de los restos de la batalla, los prolegómenos y el desarrollo de la confrontación entre las amibas y su caldo, por un lado, y las defensas del roedor en cuestión, por otro. Mal parados quedaban en esos análisis postreros los defensores, los leucocitos, a los que se les adjudicaba la culpabilidad de no haber siquiera advertido la llegada de los protozoarios intrusos.
El cultivo axénico cambió el panorama y dotó al experimentador de condiciones adecuadas para analizar el efecto devastador de las amibas patógenas, tanto en modelos de laboratorio, in vitro, como en modelos animales en los que ahora solamente se inoculan amibas.
Los sistemas in vitro han confrontado amibas patógenas con células humanas libres como glóbulos rojos, leucocitos polimorfonucleares y macrófagos, o bien con células fibroblásticas o epiteliales de mamífero, a los que se les añaden los parásitos. El tiempo de experimentación se reduce drásticamente en estos sistemas; la acción letal de las amibas se estudia, no en semanas o meses, como era tradicional en los modelos de animales de experimentación, sino en horas o aún en minutos.
A continuación se relata en forma resumida, la secuencia —no siempre uniforme, pues la amiba no sabe de formalismos— del efecto lítico de la amiba cuando se pone en contacto con células de mamífero.
El paso primero es la adhesión de los trofozoítos a las células blanco. La superficie de unos y otras entra en estrecho contacto sin llegar no obstante a la fusión de las membranas plasmáticas en interacción. A diferencia de lo que piensan muchos investigadores de mente molecular, pero de ignorancia microscópica —porque nunca observan al microscopio las células que estudian— el contacto no debe necesariamente ser prolongado. La microcinematografía revela cómo las amibas tocan a sus víctimas pero no se aferran a ellas; es el efecto que llaman los anglosajones de hit and run, "golpear y huir". Hay consenso general de que ese contacto estrecho, aunque fugaz, es necesario; de no ocurrir, no se inicia la fase siguiente, la del daño de la célula blanco. Esto significa que, o bien las amibas patógenas no liberan al medio los componentes que afectan las células, o estas toxinas son inactivadas rápidamente en el medio extracelular; se requerirá por ello contacto estrecho para crear un espacio cerrado en el que las toxinas amibianas se concentren y ejerzan su acción lítica, libres ya del efecto neutralizador o diluyente del líquido extracelular.
Movidos por la moda, sin duda plausible en su intento de interpretar toda relación entre parásito y huésped en términos moleculares, varios investigadores han buscado afanosamente compuestos amibianos que faciliten la adhesión. Se han descrito lectinas en la superficie de las amibas y se ha demostrado que al añadir azúcares específicos (si bien en concentraciones tales que más que endulzar el medio lo que se hace es convertirlo en verdadero jarabe) se impide la adhesión y se elimina parcialmente el efecto citopático. Seguramente existen ciertos mecanismos de reconocimiento molecular que facilitan la interacción entre parásitos y células víctimas, pero parece lógico pensar que la adhesión amibiana no es un fenómeno puramente químico que depende de la interacción de una especie molecular con otra.
Al cabo de pocos minutos, después del contacto con las amibas, las células blanco empiezan a dar señales de alteración; las delicadas microvellosidades que recubren la porción externa de las células epiteliales desaparecen o se engruesan grotescamente y las zonas de contacto entre células vecinas, o uniones celulares, pierden cohesión. Las capas celulares empiezan a fragmentarse; al retraerse las células individuales, se crean espacios cada vez mayores entre las células. Este daño incipiente sólo puede ser demostrado mediante microscopía electrónica o registros electrofisiológicos que analizan la estructura o la integridad funcional, respectivamente, de la superficie de las células empleadas como blanco de los parásitos. La microscopía electrónica de barrido muestra con claridad las deformaciones morfológicas de las microvellosidades de la superficie epitelial y la pérdida de continuidad de las monocapas celulares como resultado de la apertura de las uniones celulares (Figura 19). A su vez, la resistencia al paso de la corriente eléctrica de un lado a otro de la monocapa en cultivo, índice fiel de la integridad de la capa celular, se abate casi por completo, tan sólo cinco minutos después del inicio del enfrentamiento entre amibas patógenas y células epiteliales. Aun cuando no se han identificado con seguridad las moléculas responsables de las alteraciones descritas, existe, sin embargo, la posibilidad de que intervenga en el daño una proteína liberada por la amiba, llamada proteína formadora de poros, descubierta simultáneamente en la Universidad Rockefeller y en el Instituto Weizmann de Israel, en 1981. Dicha proteína tiene la particularidad de insertarse en las membranas de las células blanco y crear canales a través de los cuales entran y salen los iones, lo que rompe el gradiente iónico entre citoplasma y núcleo, requerido para funciones vitales de las células. No hay duda del interés del hallazgo de la proteína formadora de poros, o amiboporo, pero su papel en la génesis de las lesiones amibianas no ha sido demostrado; por ello, y por el hecho de que el efecto citopático es, como veremos, multifactorial, los intentos de reducir la amibiasis invasora a una enfermedad producida por la liberación de esa proteína resultan, en el mejor de los casos, ingenuos.
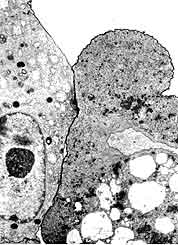
Figura 19. Región de contacto entre una amiba y una célula epitelial. En la porción superior ha penetrado parcialmente un colorante en la célula epitelial, a consecuencia de una lesión en la membrana plasmática.
El daño a las células blanco continúa progresivamente hasta llegar a producir degeneración del núcleo y del citoplasma y pérdida de la continuidad de la membrana plasmática; las células se redondean, se despegan del substrato y mueren (Figura 20). Además del efecto de la proteína formadora de poros, la lisis celular puede ser producida por la acción de proteasas, glucosidasas y toxinas amibianas descritas por numerosos investigadores en años recientes. Cada grupo proclama la preminencia de la toxina o de la enzima descrita por ellos; no es remoto que, a fin de cuentas, todos tengan razón, y la acción patógena de las amibas, después del contacto, se lleve a cabo por liberación de más de una toxina y más de una enzima.
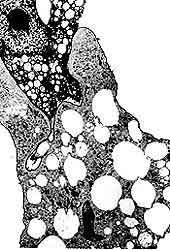
Figura 20. Fagocitosis de una célula epitelial muerta por un trofozoíto de Entamoeba histolytica.
Lo que hemos calificado de ignorancia microscópica ha hecho que los investigadores moleculares se olviden de la necesidad de tomar en cuenta la posible participación de fenómenos mecánicos en la realización del efecto citopático de las amibas patógenas. Las evidencias morfológicas muestran, sin embargo, que la movilidad de las amibas, al desplazar células alteradas e invadir activamente resquicios intercelulares, contribuye a la destrucción de las capas celulares. A ello se unen dos fenómenos, también dependientes de la motilidad del parásito: uno es el pinzamiento por parte de la amiba de pequeñas porciones de la célula blanco, seguido de la retracción del parásito; esto crea una solución de continuidad en la superficie y el inicio de la muerte de la célula. El otro proceso, la fagocitosis, ya ha sido mencionado anteriormente; las amibas incorporan al citoplasma, mediante succión, células generalmente dañadas previamente por contacto con el parásito.
La fase final del efecto citopático es la degradación intracelular de las células o del material extracelular ingerido. La fagocitosis juega un papel crucial en la realización de ese efecto citopático. Además de las pruebas citológicas, casi palpables, de la existencia de este fenómeno, Esther Orozco ha logrado demostrar cómo las variaciones en la virulencia de una cepa amibiana van acompañadas de modificaciones concomitantes en su fagocitosis; si se eliminan de una población heterogénea los elementos más fagocíticos, disminuye la virulencia; si, por el contrario, se recupera la virulencia de una cepa a través de pases sucesivos por el hígado de animales, el resultado será, junto con el incremento en la virulencia, el aumento en la capacidad fagocítica de esa cepa.
Así pues, las voraces amibas destruyen las células en sistemas in vitro por una combinación de factores que incluyen la lisis por contacto, la fagocitosis y la degradación intracelular; el resultado es la total destrucción del cultivo. A esto se aúna la capacidad de las amibas para liberar enzimas como la colagenasa, descrita por Lourdes Muñoz; al actuar esta enzima sobre la matriz conjuntiva de los tejidos, permite seguramente la invasión del parásito a través de los componentes extracelulares.
Recordemos que la acción patógena de un microorganismo no depende tan sólo de su capacidad para liberar moléculas que dañen las células del huésped o debiliten el tejido conjuntivo que mantiene la cohesión de los tejidos; debe, también, ser capaz de eliminar, o al menos atenuar, los efectos de las defensas del huésped. Este aspecto crucial de la interrelación huésped-parásito había sido muy poco estudiado. Se aceptaba, simplemente, que las amibas despertaban muy poca reacción inmunológica en el huésped; si acaso, se mencionaba la producción de anticuerpos ineficientes; prevalecía el dogma de la total ausencia de inflamación en la amibiasis invasora.
La experimentación in vitro dio información de gran interés para comprender los medios de los que se valen las amibas patógenas para reducir la eficacia de la reacción molecular y celular despertada por la presencia del invasor en los tejidos. del organismo humano. Se sabe ahora que las amibas son capaces de contender exitosamente con leucocitos polimorfonucleares y con macrófagos. En este asunto es particularmente difícil realizar en el laboratorio experimentos que tengan relevancia para la situación presente en la amibiasis invasora. ¿Cómo remedar la confrontación entre leucocitos y amibas?; sobre todo, ¿cuáles son las proporciones que se presentan durante el inicio de las lesiones? Sabemos ahora que una sola amiba patógena es capaz de eliminar varios cientos de polimorfonucleares; esa sola amiba puede producir la muerte de cerca de un centenar de macrófagos activados; es difícil decidir si la proporción es la correcta; en todo caso las observaciones atestiguan la formidable capacidad de las amibas para resistir y vencer las defensas celulares del organismo.
Estos parásitos han logrado también, a través de largo periodo de selección, adoptar mecanismos que les permiten evadir componentes moleculares de la reacción de defensa; las amibas patógenas resisten concentraciones elevadas de complemento o bien desarrollan gradualmente resistencia al mismo y, por otro lado, son capaces, como hemos visto anteriormente, de movilizar los complejos antígeno-anticuerpo localizados en la superficie del parásito, además de eliminar antígenos solubles que pueden realizar una labor de "distracción" al actuar sobre ellos los anticuerpos producidos contra las amibas.
Del conjunto de estudios realizados hasta la fecha sobre la acción patógena de la amiba, podemos concluir que se trata de un fenómeno complejo, multifactorial, no necesariamente ordenado en una secuencia definida. No existía duda alguna de que la amiba estuviese dotada de un armamento espeluznante capaz de desintegrar a la mayoría de los tejidos del cuerpo humano; pero ha sido sólo en los últimos años en los que se han empezado a conocer estas armas: moléculas agresoras y fenómenos dependientes de movilidad —adhesión, pinzamiento, fagocitosis— que, en conjunto, hacen que nuestro parásito tenga bien ganada la fama de su nombre.
Una cuestión relevante, aún no resuelta, es el aumento en la virulencia de las amibas producida por asociación con bacterias no patógenas. Durante muchos años después de la obtención de cultivos axénicos, se consideraban éstos como curiosidad de laboratorio, pues se pensaba que no tenían capacidad de producir lesiones. Fue gracias a la perseverancia de Miguel Tanimoto, en el hoy destruido Centro Médico Nacional, que se logró demostrar la patogenicidad de estas amibas; no se requiere en forma absoluta la asociación bacteriana para que las amibas ejerzan papel patógeno. La reiterada observación de la ausencia de bacterias en los abscesos hepáticos amibianos no había sido tomada en cuenta, a pesar de que indica en forma clara la virulencia de las amibas per se. Sin duda, es difícil ser profeta en su tierra; este hallazgo ha pasado casi desapercibido en el conocimiento de la amibiasis, pero ha sido mucho más importante que cualquiera de las complejas caracterizaciones de moléculas amibianas a las que nos hemos referido. Antes de que se demostrara la patogenicidad de las amibas axénicas, el problema de la virulencia amibiana se reducía a comprender cómo eran las bacterias capaces de impartir malévolas propiedades a las inocentes amibas; ahora sabemos que las amibas por sí mismas dañan animales de experimentación, la asociación con las bacterias se plantea en términos diferentes: entender por qué esta asociación exalta la virulencia del parásito. David Mirelman, en Israel, ha abordado el problema y encuentra que son sólo unos cuantos tipos de bacterias los que producen este aumento en la virulencia: pero nada sabemos aún sobre el mecanismo de la inducción.
Armados de un mejor conocimiento del efecto citopático producido por los trofozoítos y sabiendo que las amibas axénicas, por sí mismas, pueden provocar lesiones, iniciamos la indagación de la génesis de las lesiones en el absceso hepático y la ulceración intestinal amibiana.
Después de que Tanimoto demostró la formación de abscesos hepáticos en el hámster con inoculación directa de amibas axénicas, Víctor Tsutsumi perfeccionó el modelo al emplear la vía portal, más natural, como vía de inoculación. Logró la reproducción constante de lesiones que evolucionan hacia abscesos en todo semejantes a los encontrados en las fases finales de la amibiasis hepática en el humano. El modelo descrito ha permitido a Tsutsumi derribar, al menos en el caso de la amibiasis experimental, el dogma de la ausencia de reacción inflamatoria en la amibiasis invasora, y hacerlo, además, de forma contundente. Según sus observaciones, esta reacción no sólo está presente en los estadios iniciales del establecimiento de la lesión hepática, sino que es la misma lisis de las células inflamatorias la que provoca la extensa destrucción del hígado. Seis horas después de llegar las amibas al hígado, la reacción inflamatoria aguda alrededor de los trofozoítos es notable; a este tiempo empieza ya a observarse lisis de las células inflamatorias, ésta se acentúa rápidamente en el transcurso de las siguientes horas. Poco tiempo después, los leucocitos muertos son sustituidos por macrófagos, que experimentan la misma suerte que los primeros, a pesar de utilizar sus mejores tácticas de defensa, como es la formación de empalizadas, en un vano intento por contener la extensión de la infección. Esta inflamación granulomatosa evoluciona rápidamente hacia la necrosis; áreas cada vez más grandes del hígado se necrosan, las lesiones confluyen entre sí y producen al cabo de una semana uno o varios abscesos. Ignoramos si la secuencia de eventos aquí descrita ocurre en el humano durante el desarrollo del absceso hepático. Resulta evidente la dificultad de analizar en el humano, en el curso de los primeros días, lo que ocurre después de la llegada al hígado.
La amibiasis intestinal experimental era el último reducto de los que consideraban a las amibas axénicas como artificios de laboratorio, debido a que varios trabajos habían mostrado la imposibilidad de producir lesiones ulcerativas en el intestino de animales de laboratorio inoculados con tales parásitos. Fernando Anaya produjo esas lesiones en hámster o en cobayo, a condición de liberar, en la medida de lo posible, a las amibas de la influencia nociva del material intestinal normalmente presente en esos animales. Cuando esto se lleva a cabo, las amibas producen en menos de dos días úlceras visibles macroscópicamente en el ciego de los animales. El análisis aún inconcluso de este modelo sugiere también la participación de las células inflamatorias en la génesis de las lesiones intestinales.
A pesar de los grandes avances en cuanto al conocimiento de la patogenia de la amibiasis, aún queda mucho por saber; sobre todo, si recordamos que lo ya descrito es válido sólo para la amibiasis experimental en animales de laboratorio. Así, queda por demostrar cuáles de los fenómenos descritos tienen realmente un papel en la formación de las lesiones amibianas en el hombre. Creer que nuestros modelos son representativos de la enfermedad en el hombre tranquiliza nuestras conciencias, justifica nuestro trabajo y promueve la canalización de fondos para la investigación, pero al fin y al cabo debemos recordar que los resultados ofrecen tan sólo una posibilidad, pendiente de demostración final.