VI. INTERFERÓN E INMUNIDAD A LAS INFECCIONES VIRALES
EL SISTEMA inmune constituye la principal barrera que poseen los
animales superiores para protegerse de las infecciones en general. La inmunidad
inespecífica es aquélla que actúa contra cualquier material extraño que invade
al organismo; este tipo de inmunidad es innata y en ella participan barreras
físicas como la piel y las mebranas mucosas; barreras químicas como el ácido
clorhídrico del jugo gástrico o el ácido láctico presente en el sudor; proteínas
que inactivan a los agentes infecciosos o destruyen a las células infectadas,
como la lisozima presente en las lágrimas y secreciones mucosas, las enzimas
del sistema de complemento y los interferones. Existe también una importante
variedad de células que participan en los mecanismos de inmunidad inespecífica:
los macrófagos o fagocitos que ingieren y destruyen células y partículas extrañas,
al igual que los leucocitos neutrofilos que además son importantes mediadores
del proceso de inflamación. Los macrófagos también actúan como células presentadoras
de antígenos que estimulan la respuesta inmune específica; los leucocitos basófilos
que secretan mediadores químicos que promueven la inflamación con el fin de
facilitar el flujo sanguíneo y la accesibilidad de las células inmunitarias
en las regiones donde se localiza una infección; los leucocitos eosinófilos
que participan en la destrucción de parásitos y de células infectadas por parásitos
intracelulares; las células asesinas naturales (NK por "natural killers") que
son estudiadas más adelante.
Existen dos tipos de inmunidad específica: humoral y celular. La inmunidad humoral es mediada por los anticuerpos o inmunoglobulinas, que son proteínas específicas sintetizadas por las células plasmáticas. Los progenitores inmediatos de estas células son los linfocitos B que son capaces de reaccionar con moléculas extrañas al organismo, llamadas antígenos. Los linfocitos B se originan en la médula ósea a partir de células precursoras. La unión del antígeno con el linfocito B estimula a este último para que se divida y se diferencie dando origen a una gran cantidad de células plasmáticas y células poseedoras de la llamada memoria antigénica. Las células plasmáticas sintetizan y secretan moléculas de anticuerpos, las cuales reacionan en forma específica con el antígeno que originalmente estimuló la producción de dichos anticuerpos. Las células con memoria antigénica no sintetizan anticuerpos pero conservan la capacidad potencial de sintetizar dichos anticuerpos contra un antígeno específico, pues las células con memoria antigénica pueden transformarse, en condiciones adecuadas, en células plasmáticas productoras de anticuerpos específicos. Por esta razón, el sistema inmune de un organismo responde en forma más potente y rápida cuando se encuentra por segunda vez ante la presencia del mismo antígeno.
Las inmunoglobulinas se dividen en cinco grupos: IgA, IgG, IgD, IgE e IgM. Estas proteínas actúan directamente uniéndose a componentes del virión y produciendo la neutralización del virus. Sin embargo, las IgG pueden unirse por medio de sus fragmentos Fc a la superficie de otras células del sistema inmune conocidas como macrófagos, los cuales adquieren la capacidad para reconocer los antígenos virales presentes en la superficie de las células infectadas.
Las IgM e IgG pueden activar el sistema del complemento que consiste en una cascada de enzimas que liberan componentes capaces de atraer diversos tipos de células del sistema inmune al sitio donde se localiza el anúgeno; estas enzimas también expresan una actividad de fosfolipasa capaz de lisar las células infectadas por el virus.
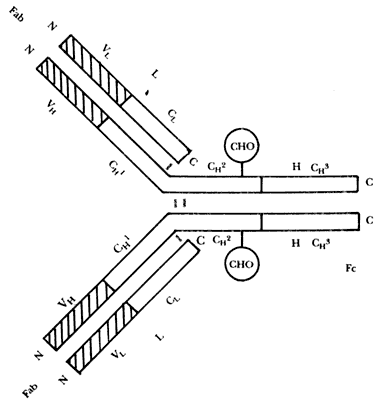
Esta representación esquemática de una molécula de IgG muestra las cadenas pesadas (H) y las cadenas ligeras(L). Los extremos N de los fragmentos Fab corresponden a las regiones de unión del anticuerpo con su antígeno específico. El fragmento Fc es común a todas las moléculas de IgG independientemente de su especificidad antigénica. Se indican las regiones con secuencias variables (V) y con secuencias constantes de aminoácidos (C). La sigla CHO indica la posición de moléculas de carbohidratos asociados con la proteína IgG; estos puentes disulfuro estabilizan la estructura de la IgG.
La inmunidad celular es mediada en primer lugar por los linfocitos T, los cuales se originan en el timo. La especificidad de los linfocitos B yT está determinada por receptores específicos que están presentes en las membranas de estas células. Estos receptores reconocen motivos estructurales específicos que están presentes en los antígenos; dichos motivos estructurales son conocidos como epitopes y son muy diferentes entre sí. El reconocimiento y ligado de un epitope por parte del receptor celular específico es el evento que determina que sean activados sólo los linfocitos que poseen dichos receptores. Los recepctores específicos presentes en un linfocito B son en realidad inmunoglobulinas que están fijas a la membrana celular y que tienen la misma especificidad que las inmunoglobulinas secretadas por dicho linfocito.
En el caso de los linfocitos T, el receptor para epitopes antigénos específicos se denomina receptor de célula T o receptor T y consiste en una proteína que a su vez resulta de la conjunción de dos proteínas diferentes denominadas alfa y beta, respectivamente. Dicho receptor T presenta un extremo variable que determina la especificidad del receptor por un epitope en particular. Cuando el epitope es ligado por el receptor T se produce una señal que estimula al linfocito T para que prolifere y produzca una "clona" o familia de células idénticas que manifiestan la misma especificidad por el mismo epitope antigénico. Algunas de las células T resultantes serán responsables de contribuir a mantener la memoria inmunológica que permite una respuesta más eficaz y rápida cuando el organismo encuentra al mismo antígeno por segunda vez.
Los principales tipos de linfocitos T son:
1) linfocito T auxiliar: este tipo de linfocito prolifera después de haber reconocido y ligado un epitope específico, dando origen a clona de linfocitos auxiliares que producen una variedad de moléculas solubles conocidas como linfocinas cuyo papel consiste en estimular a otras células inmunocompetentes (linfocitos B y T) para que se activen y proliferen. Los linfocitos T axuliares son orquestadores de la respuesta inmune específica; sin la participación de estos linfocitos no es posible la producción de otras células inmunocompetentes que participan en la respuesta inmune específica.
2) linfocito T citotóxico: este tipo de linfocito responde a la activación mediada por el reconocimiento de un epitope específico y la presencia de linfocinas en el medio, dando origen a una clona de células T citotóxicas con capacidad para destruir células que portan antígenos foráneos específicos como son las células infectadas por virus. Los linfocitos T citotóxicos producen también ciertas linfocinas como el interferón gamma, el cual estimula la actividad de los macrófagos.
3) linfocito T supresor: este tipo de linfocito se encarga de disminuir o amortiguar la maginitud de la respuesta inmune y de esta manera controla la respuesta de otras células inmunocompetentes para evitar que sea exagerada.
Los linfocitos denominados células asesinas naturales (células NK) son linfocitos grandes con citoplasma granular y son diferentes de los linfocitos T y B. Los linfocitos NK destruyen a células infectadas por cualquier tipo de virus, por lo cual su actividad es inespecífica. Las células NK se adhieren a las células infectadas y liberan gránulos tóxicos que lisan (destruyen) a las células blanco.
En 1957, Isaacs incubó la membrana corioalantoica de un embrión de pollo en presencia de una suspensión de virus de la influenza que había sido inactivado por medio de tratamiento térmico. Esta membrana fue transferida a una solución amortiguadora y se almacenó por 24 horas. Posteriormente, la membrana fue descartada y la misma solución amortiguadora fue utilizada para incubar una nueva membrana de pollo en presencia de virus infeccioso de la influenza. Isaacs observó que la nueva membrana no permitía el crecimiento del virus y por lo tanto concluyó que algún producto soluble con actividad antiviral había sido liberado en la solución amortiguadora como respuesta a la infección viral de la membrana original. Esta sustancia antiviral fue denominada interferón.
Los interferones son proteínas que tienen asociados carbohidratos senciales
para su actividad. Por convención, la actividad antiviral del interferón es
estimada midiendo la inhibición que éste produce en la incorporación de uridina
radiactiva en el ARN viral en células infectadas por un togavirus. La actividad
es expresada como la cantidad de interferón necesaria para reducir en 50% el
nivel normal de síntesis de ARN viral; esta cantidad es arbitrariamente definida
como una unidad de interferón. Por ejemplo, 1 mg de proteína de interferón purificado
tiene actividad antiviral en el orden de 109 unidades.
Los interferones fueron identificados como proteínas secretadas por células infectadas por virus que son capaces de proteger de la infección viral a otras células, debido a que los interferones estimulan en las células no infectadas la producción de proteínas que inhiben la replicación de diferentes tipos de virus. Sin embargo, hoy en día el término interferón se refiere a varias proteínas que manifiestan esta actividad antiviral aunque no todas estas proteínas son producidas por células infectas por virus. Existen dos tipos principales de interferón. los tipo 1 están representados por el interferón alfa (IFN alfa) y el interferón beta (IFN beta); ambos tienen una actividad biológica muy similar siendo ejemplos de la llamada respuesta inmune inespecífica y sus estructuras moleculares son muy parecidas. El IFN alfa es producido principalmente por los leucocitos infectados por virus, mientras que el IFN beta es producido por fibroblastos infectados por virus. El IFN alfa también estimula la sintésis de proteínas clase 1 del complejo mayor de histocompatibilidad (MHC- clase 1), estas moléculas están presentes en las membranas de todas las células con núcleo y participan en la presentación de antígenos (en particular de antígenos virales) para que sean reconocidos por el sistema inmune.
La figura VI.1 describe el mecanismo de acción antiviral del interferón. Una gran variedad de virus es capaz de inducir la producción de interferón tipo 1; el cual es activo contra un amplio espectro de virus diferentes y no sólo contra el virus inductor. Sin embargo, el interferón parece ser más específico en relación con la especie a partir de la cual se obtienen las células productoras del mismo. Por ejemplo, interferón obtenido a partir de células de ratón es poco eficiente para proteger células de rata, pollo o simio contra la infección viral.
La inducción del interferón provoca la liberación de esta molécula y la producción
de un estado antiviral en las células que entran en contacto con el interferón.
Los genes que codifican a los interferones humanos tipo 1 están ubicados en
el cromosoma 9, mientras que el gene del interferón tipo 2 está en el cromosoma
12. Todos los virus que se multiplican activamente son capaces de inducir la
producción de interferón y se cree que moléculas de ARN de cadena doble actúan
como inductores específicos. El interferón liberado produce.el estado antiviral
en otras células por medio de la unión con un receptor presente en la superficie
celular. El gene que codifica al receptor para IFN alfa y beta está en el cromosoma
21, mientras que el gene que codifica el receptor para IFN tipo 2 o gamma, está
en el cromosoma 6. Se sabe que la presencia de ARN de cadena doble estimula
la fosforilación (adición de grupos fosfato) de ciertas proteínas reguladoras;
esto impide que estas proteínas participen en el proceso de iniciación de la
síntesis de proteínas. Este ARN de cadena doble también activa una ribonucleasa
que degrada al ARN mensajero y, por lo tanto, detiene la síntesis de proteínas.
Sin embargo, todavía no se conoce con claridad el mecanismo por el cual las
actividades inducidas por el interferón son capaces de distinguir ente la síntesis
de proteínas celulares y la síntesis de proteínas virales De hecho, la actividad
inhibitoria de la síntesis de proteínas parece ser la responsable del efecto
antitumoral del interferón, efecto que ha sido claramente demostrado in vitro
y en animales experimentales. Sin embargo, el uso de los diferentes interferones
en el tratamiento del cáncer no ha dado los resultados esperados, ya que por
lo general los interferones producen efectos secundarios negativos en los pacientes
tratados con dosis altas de estos agentes. Hasta la fecha, sólo algunos tipos
de cáncer muy raros, como por ejemplo, la leucemia de células pilosas, han sido
tratados con éxito mediante el uso de interferón alfa.
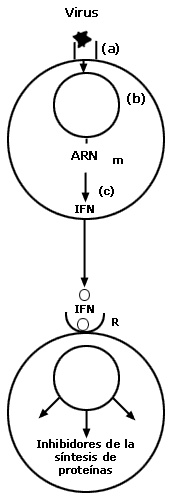
Figura VI.1. Mecanismo de acción del interferón: el virus infecta a la célula
1 después de unirse con el receptor (a). La infección viral enciende la maquinaria
celular para permitir la replicación del genoma viral (b). La presencia de ácido
nucleico viral induce la expresión de genes de interferón (c). El interferón
es secretado por la célula infectada y se pega a su receptor específico presente
en la membrana de una célula no infectada (d). La unión del interferón con su
receptor induce la producción de enzimas que interfieren con la síntesis de
proteínas. Una de estas enzimas inhibe la traducción de ARN mensajero viral,
mientras que otra enzima estimula la acción de enzimas endonucleasas que degradan
el ARN mensajero viral. De esta manera, la célula receptora del interferón queda
protegida de la infección viral. Al inducir estas dos acciones enzimáticas que
interfieren con la síntesis de proteínas, el interferón inhibe también el crecimiento
de las propias células, por ello ha sido utilizado experimentalmente como inhibidor
de la proliferación de células cancerosas.
El interferón tipo 2 está representado por el IFN gamma, también denominado
inmunointerferón, debido a que es producido por linfocitos activados al entrar
en contacto con antígenos durante una respuesta inmune. La principal acción
de este tipo de interferón consiste en actuar como una linfocina, es decir,
como una molécula capaz de activar otras células del sistema inmune, como son
las células asesinas naturales (NK) los macrófagos, y los linfocitos B. De hecho,
el IFN gamma puede influir sobre la clase de anticuerpo que es producido por
los linfocitos B en el marco de una respuesta inmune. La inducción de este tipo
de interferón no depende de la presencia de ARN de cadena doble y al parecer
tiene un papel suplementario en la respuesta inducida por el interferón tipo
1. El interferón tipo 2 es particularmente eficaz en el control de infecciones
producidas por virus capaces de inhibir la síntesis de ARN y proteínas celulares
antes de que haya sido posible la síntesis de interferón tipo 1.