I. INTRODUCCIÓN HISTÓRICA AL ESTUDIO DE LOS VIRUS
a) UN ANTIGUO PROBLEMA EN LA PATOLOGÍA DE LOS SERES VIVOS
El términó virología ha sido incorporado al vocabulario durante las
últimas décadas. La primera revista científica dedicada exclusivamente al campo
de la virología: Archiv für die Gesamte Virusforschung, hoy conocida
como Archives of Virology, empezó su publicación en 1939. El primer texto
dedicado sólo a la virología: General Virology, de Salvatore Luria, fue
publicado en 1953. Sin embargo, los virus han estado acompañando al hombre durante
toda su historia y el término virus tiene muchos siglos de existencia, aunque
su uso y connotaciones han variado notablemente a lo largo del tiempo. Se puede
decir, en forma un tanto arbitraria, que los orígenes de la disciplina científica
hoy día conocida como virología apenas se remontan a las décadas finales del
siglo XIX. Pero considerando aspectos epidemiológicos*  y
semiológicos**
y
semiológicos** ![]() dentro
del registro histórico, encontramos que enfermedades como la rabia han sido
descritas y registradas meticulosamente por más de dos mil años. En el caso
particular de la rabia, su misterioso origen, su capacidad para transformar
a un perro domesticado en bestia feroz, su largo e impreciso periodo de incubación
y los dramáticos síntomas que preceden al final fatal de esta enfermedad en
los humanos, constituyen un cuadro pavoroso y a la vez irresistible, que ha
merecido la atención de los escritores y pensadores de la Antigüedad.
dentro
del registro histórico, encontramos que enfermedades como la rabia han sido
descritas y registradas meticulosamente por más de dos mil años. En el caso
particular de la rabia, su misterioso origen, su capacidad para transformar
a un perro domesticado en bestia feroz, su largo e impreciso periodo de incubación
y los dramáticos síntomas que preceden al final fatal de esta enfermedad en
los humanos, constituyen un cuadro pavoroso y a la vez irresistible, que ha
merecido la atención de los escritores y pensadores de la Antigüedad.
Si se comparan las antiguas descripciones hechas por cronistas griegos y romanos con los casos más recientes de rabia tanto en animales como en humanos, encontramos que desde el punto de vista clínico el virus de la rabia no ha cambiado a lo largo de los siglos. Esta característica lo distingue de otros virus patógenos (capaces de producir enfermedad), confiriéndole una posición única en la historia de la medicina. Diferentes ideas acerca de la causa y el origen de la rabia han sido concebidas en diferentes épocas. Una de las primeras descripciones de la rabia que han sobrevivido hasta nuestros días es la de Aristóteles. "la rabia vuelve loco al animal, y cualquier especie de animal, con excepción del hombre, será contaminado con esta enfermedad si es mordido por un perro rabioso. La enfermedad es fatal para el propio perro y cualquier otro animal mordido por éste, con excepción del hombre". Autores posteriores muchos de los cuales dudaron en cuestionar la credibilidad del gran filósofo griego, se mostraron sorprendidos por la referencia de Aristóteles a la supuesta insusceptibilidad del ser humano para ser contaminado por la rabia. Así, algunos autores propusieron que originalmente el agente causal de la rabia no podía contagiar al hombre y solamente al cabo de los siglos el misterioso agente causal de esta enfermedad había adquirido la capacidad de afectar al ser humano. Sin embargo, el médico renacentista Girolamo Fracastoro hizo notar que Aristóteles solamente quería recalcar el hecho de que no todos aquellos humanos mordidos por un perro rabioso desarrollarían la enfermedad en forma obligatoria.
Una descripción de la rabia más precisa y detallada fue proporcionada en el
libro De medicina, escrito por Celso, un médico que vivió en el siglo
I, en el apogeo del Imperio romano. En particular, hay una frase en el
texto de Celso que ha llamado poderosamente la atención de los historiadores
de la medicina: "...especialmente en los casos en que el perro es rabioso, el
virus debe ser drenado con una ventosa de vidrio..." Por supuesto nadie
se atrevería a pensar que Celso identificó al agente de la rabia como un virus
en el sentido moderno del término. Sin embargo, es importante hacer notar que
Celso utilizó el término virus para denotar al agente causal de la rabia, mientras
que en el mismo texto utilizó la palabra venenum para describir la ponzoña
de las serpientes. Es probable que tal distinción no haya sido accidental, sobre
todo si consideramos que el término latino virus puede significar veneno
o también líquido viscoso. Por lo tanto, quizá Celso estaba advertido de que
el agente de la rabia era transmitido por medio de la saliva viscosa del perro
rabioso.
Después de Celso, el término virus se utilizó durante siglos en forma casual
como sinónimo de ponzoña o veneno, hasta que a finales del siglo XVIII
adquirió claramente el significado de un agente infeccioso, debido a la creciente
advertencia general de que existen muchas enfermedades contagiosas y transmisibles.
La gradual aceptación del término virus en la literatura médica corrió paralela
con el desarrollo de los conceptos de infección y contagio, los cuales deben
su origen al estudio de una enfermedad también de naturaleza viral, pero que,
a diferencia de la rabia, ha tenido enorme influencia sobre el curso de la historia
social y política de la humanidad: la viruela.
En términos de la devastación causada en las sociedades medievales, la viruela es solamente comparable con la peste bubónica. Sin embargo, la historia temprana de la viruela presenta grandes dificultades para el historiador de la medicina, ya que, a diferencia de la rabia, es muy difícil establecer un diagnóstico diferencial entre la viruela y otras enfermedades eruptivas de tipo febril, como el sarampión, la varicela o la escarlatina, a partir de las descripciones proporcionadas por los cronistas de la Antigüedad. Pero no cabe duda de que los conquistadores españoles contaron con un inesperado, silencioso y mortal aliado que contribuyó notablemente al éxito de Cortés y a la pronta caída de Tenochtitlán. Un soldado de la expedición de Pánfilo de Narváez arribó a México enfermo de viruela, enfermedad hasta entonces desconocida en Mesoamérica. La falta de inmunidad natural a la viruela permitió que ésta se extendiera rápidamente entre la población indígena con desastrosas consecuencias para la misma. En pocas semanas miles de indígenas sucumbieron a la viruela; recordemos que el propio Cuitláhuac, penúltimo emperador azteca, falleció por causa de esta enfermedad. Recientes estimaciones epidemiológicas han llevado a postular que durante los primeros veinticinco años posteriores a la Conquista más de un tercio de la población indígena sucumbió a la viruela. Es probable que tal devastación natural haya contribuido en forma radical al establecimiento del régimen colonial, explicando también en parte por qué imperios tan poderosos y organizados como el azteca y el inca fueron borrados del mapa, sin mayor oposición, en unos cuantos años.
Durante los siglos XVII y XVIII ocurrieron en Europa
severas epidemias de viruela, posiblemente debidas a la aparición de nuevas
cepas del virus. Médicos y sabios que testimoniaron estas epidemias pudieron
darse cuenta de que algún factor esencial era transmitido de persona a persona
y de casa en casa. En la antigua China, mucho antes de nuestra era, ocurrieron
los primeros intentos para proteger a los individuos contra la viruela. Así,
los chinos desarrollaron la práctica conocida como variolación, la cual
consiste en la inoculación de individuos sanos con fluidos obtenidos de las
vesículas eruptivas de las personas afectadas por casos leves de viruela. En
1721, Mary Worfley Montagu, esposa del embajador británico en Turquía, introdujo
la variolación en Gran Bretaña. Dicho procedimiento resultaba muy peligroso,
ya que con frecuencia los individuos inoculados desarrollaban la enfermedad
y morían a consecuencia de ella, en lugar de ser protegidos contra la viruela.
Los problemas causados por las severas epidemias de viruela y la controversia
en relación con la variolación fueron la base de mucha bibliografía médica producida
en el siglo XVIII. Algunos autores se permitieron especular sobre
la naturaleza del agente transmisible causa del contagio, al cual se denominó
virus cada vez con mayor frecuencia.
En 1730, Thomas Fuller publicó un pequeño libro sobre las fiebres eruptivas (que incluyen la viruela). Ahí escribió: "La forma principal y más común de contraer las fiebres contagiosas, como la viruela y el sarampión, es por medio de infección, o sea, recibiendo a través del aliento o de los poros de la piel los corpúsculos virosos peculiares a la crianza de dichas enfermedades."
Por su parte, Angelo Gatti, el gran precursor de Jenner, publicó sus reflexiones sobre la variolación en 1764. Sus observaciones en relación con la naturaleza de la infección y con la necesidad de encontrar un método para atenuar "el virus varioloso", fueron de carácter profético. Sin embargo, Gatti murió en enero de 1798, antes de haber podido conocer el modesto panfleto publicado en el mismo año por el médico británico Edward Jenner, en el cual comunicaba su descubrimiento de que la inoculación con el virus de la vacuna (o sea, con los fluidos obtenidos de las lesiones de la viruela bovina), era capaz de prevenir la infección de la viruela en seres humanos. El descubrimiento de la vacuna es uno de los sucesos más importantes no sólo en la historia de la medicina, sino también en la historia de las sociedades humanas. A pesar de este descubrimiento, se requirieron casi doscientos años para lograr la erradicación de la viruela.
A principios del siglo XIX, el término virus se encontraba
bien establecido en la bibliografía médica, aunque era utilizado para describir
una gran variedad de agentes infecciosos. Este uso persistió durante toda la
época del surgimiento de la bacteriología médica, o sea, los primeros dos tercios
del siglo XIX. Claude Bernard erigió una infraestructura para la
medicina experimental, apoyada en la tradición filosófica de Descartes y Pascal.
Por ejemplo, Bernard citó los resultados obtenidos a través de la simple observación
en el caso de la garrapata y su relación con enfermedades de la piel. Según
Bernard, la misma metodología aplicada en la observación directa de las garrapatas
podía ser aplicada para explorar la validez de las teorías que presumían la
existencia de ...el virus, una criatura de la razón. Así, Claude Davaine
empezó en 1860 a utilizar una combinación de experimentación y observación para
rastrear aquello a lo cual se refirió sucesivamente como el virus, el agente
tóxico y finalmente, el bacteridium del ántrax. A falta de filtros artificiales
adecuados, Davaine demostró la diferencia entre sangre infectada y no infectada
por el ántrax, utilizando como filtro la placenta del cerdo. Sin embargo, en
algunos experimentos de Davaine el agente del ántrax era capaz de atravesar
el filtro placentario. Robert Koch descubrió en 1876 que el bacilo del ántrax
a veces forma pequeñas esporas capaces de atravesar la membrana placentaria.
Mientras tanto, Chauveau empezó a trabajar en la identificación del agente de la viruela, utilizando métodos de filtración. Pero no pudo obtener resultados definitivos, por lo cual fue el primero en hacer una distinción entre enfermedades virulentas, causadas por virus, y enfermedades contagiosas, en las cuales era transmitido un microbio o parásito bien conocido. Por lo tanto, el término virus fue restringido a denotar una misteriosa entidad infecciosa capaz de ejercer su efecto patogénico solamente por medio de la asociación en solución de ciertos elementos o partículas de naturaleza desconocida. Chauveau identificó estos cuerpos elementales (granulations élémentaires) como el origen de la actividad patogénica, introduciendo así un término que sobreviviría durante décadas.
En los años 1865-1866, ocurrió en la Gran Bretaña un desastroso brote de una enfermedad llamada ictericia hemática bovina o peste del ganado, la cual exterminó una gran cantidad de animales. El médico John Gamgee había urgido, sin éxito, a las autoridades para que impusieran una cuarentena y programas de matanza controlada para evitar la diseminación de la enfermedad. A raíz de su fracaso, Gamgee abandonó el país no sin antes publicar un libro con sus observaciones sobre esta enfermedad; ahí escríbió:
Al igual que la mayoría de las ponzoñas animales, el virus de la peste del ganado se reproduce con maravillosa rapidez en los cuerpos de los animales enfermos, de los cuales es también liberado. Tanto el aliento de la res enferma aspirado por un animal sano, como lo productos sólidos de la enfermedad, parecen tener la capacidad de inducir el padecimiento y los antídotos son aplicados demasiado tarde cuando se intenta alcanzar la ponzoña presente en el sistema animal. No conozco ningún antídoto capaz de ser usado internamente en los animales enfermos... Me temo que nunca lograremos alcanzar el virus una vez que éste se encuentra en el animal vivo.
Tales palabras desalentadas merecen ser recordadas considerando los escasos avances logrados hasta la fecha en el tratamiento de las enfermedades virales.
La etiología u origen del ántrax fue cabalmente entendida gracias al advenimiento
de una herramienta que se volvió invaluable en los estudios bacteriológicos,
misma que dio lugar al surgimiento a fines del siglo XIX del concepto
de virus como entidad filtrable. Al parecer, placas de cerámica blanca
no esmaltada y conectadas a una bomba de vacío capaz de ejercer succión fueron
utilizadas en trabajos bacteriológicos por primera vez en 1870, por Edwin Klebs.
Seis años después, Louis Pasteur utilizó para el mismo propósito filtros hechos
con la llamada goma de París. Así, Pasteur, en colaboración con Joubet, aisló
el bacilo del ántrax y adoptó la palabra microbio inventada por Séedillot
en 1878, de manera que Pasteur declaró: todo virus es un microbio, haciendo
caso omiso de la distinción hecha por Chauveau entre enfermedades virulentas
y enfermedades contagiosas. A partir de entonces y a través del trabajo pionero
sobre las vacunas contra el ántrax, el cólera aviario y la rabia, Pasteur y
todos aquellos involucrados en estudios patológicos utilizaron el termino virus
para denotar cualquier agente infeccioso (ya fuera o no fuera éste de naturaleza
bacteriana) capaz de producir inmunidad después de la convalecencia del organismo
afectado por el propio agente infeccioso.
Pasteur buscó afanosamente y en vano el agente causal de la rabia. Sin embargo, no existe evidencia de que haya sospechado que dicho agente era esencialmente diferente de los otros microbios patógenos que había logrado caracterizar a lo largo de su vida. En 1897, dos discípulos de Robert Koch encabezaron un estudio sobre el origen de la fiebre aftosa del ganado vacuno. Dichos investigadores demostraron que el agente causal de esta enfermedad era de naturaleza filtrable, o sea, capaz de atravesar los filtros bacteriológicos más finos disponibles en aquel entonces. Loeffler y Frosch observaron que este agente filtrable podía ser pasado de un animal a otro a pesar de que cada pasaje implicaba una gran dilución en la concentración del propio agente filtrable. Por lo tanto, Loeffler y Frosch llegaron a la conclusión de que el agente infeccioso podía reproducirse en los animales infectados, descartándose de esta manera la posibilidad de que se tratara de una toxina, y concluyeron que se trataba de un microbio muy pequeño. No es de sorprender que los bacteriólogos médicos se negaran a buscar una explicación que estaba por fuera del concepto de microbio patógeno. Por otra parte, es notable que un microbiólogo botánico fuera capaz de sugerir, unos cuantos meses después de haberse publicado los resultados de Loeffler y Frosch, una explicación que finalmente resultó correcta en relación con estudios y observaciones similares realizados respecto al agente causal de una enfermedad vegetal conocida como mosaico del tabaco.
b) EL VIRUS DEL MOSAICO DEL TABACO
A finales del siglo XIX, los éxitos de la microbiología médica
habían establecido sólidamente la teoría de que las enfermedades infecciosas
son causadas por gérmenes microscópicos que podían ser cultivados en medios
nutritivos especiales y aislados por medio de filtros muy finos capaces de retener
dichos microorganismos. Sin embargo, en 1892 el ruso Dimitri Ivanovski demostró
que el agente causal de la enfermedad vegetal conocida como mosaico del tabaco
era capaz de pasar a través de los filtros a prueba de bacterias y no podía
ser observado bajo el microscopio ni cultivado en medios artificiales. Estos
resultados hicieron a Ivanovsky especular que el mosaico del tabaco era causado
por un germen productor de toxinas las cuales podían atravesar el filtro bacteriológico.
En 1898, el holandés Martinus Beijerinck realizó experimentos más rigurosos,
los cuales demostraron que el agente del mosaico del tabaco pasaba a través
de filtros de porcelana y era capaz de multiplicarse en los tejidos de las plantas
infectadas, hecho que descartaba la posibilidad de que se tratara de una toxina.
Beijerinck denominó Contagium vivum fluidum al agente del mosaico del
tabaco. En aquel entonces, el concepto de macromolécula no existía todavía,
y las substancias eran divididas básicamente en dos grupos; las substancias
corpusculares o particuladas, suspendidas en los fluidos circundantes, como
las bacterias y los glóbulos rojos, y las substancias disueltas o solubles,
moléculas de bajo peso molecular.
Para Beijerinck era de la mayor importancia establecer si el virus era disuelto
o corpuscular. Apoyándose en la observación de que el virus era capaz de atravesar
filtros a prueba de bacterias, Beijerinck concluyó que debía tratarse de una
substancia disuelta o soluble y, por lo tanto, de naturaleza molecular, quizá
soluble en agua y capaz de replicarse, pero solamente cuando se encontraba incorporada
al protoplasma de la célula viva infectada. Las conclusiones de Beijerinck resultaron
sorprendentemente cercanas al moderno concepto de virus; sin embargo, también
resultaron ser demasiado avanzadas para su tiempo y encontraron poca aceptación
entre los patólogos de principios del siglo. A pesar de esto, en los primeros
años del siglo XX diversos investigadores descubrieron otros agentes
infecciosos filtrables, capaces de producir enfermedades en animales, y gradualmente
el término virus, en principio usado por los médicos de la antigua Roma
para designar cualquier veneno de origen animal, pasó a denominar estos recién
descubiertos agentes infecciosos filtrables. En 1908, dos patólogos daneses
informaron haber transmitido la leucemia (un tumor caracterizado por una desmedida
proliferación de los glóbulos blancos de la sangre) a pollos, por medio de un
filtrado libre de células. Peyton Rous descubrió en 1911 que algunos tumores
sólidos, conocidos como sarcomas de los pollos, podían ser transmitidos a pollos
sanos por medio de un filtrado obtenido a partir de células tumorales. Sin embargo,
estos primeros indicios de la asociación entre los virus y el cáncer permanecieron
ignorados por muchos años.
En 1915, el inglés F. Twort publicó un artículo en la revista médica The Lancet, en el cual describió un curioso fenómeno observado en ciertos cultivos de micrococos bacterianos que después de incubaciones prolongadas desarrollaban regiones transparentes, las cuales examinadas al microscopio mostraban la ausencia de células bacterianas y la presencia de minúsculos gránulos cristalinos. El material cristalino podía ser pasado por filtros de porcelana y una gota de ese filtrado era suficiente para destruir (lisar) un nuevo cultivo de micrococos. Twort sugirió que el fenómeno podía ser explicado por la existencia de un virus bacteriano o por la producción de una enzima bacteriana capaz de degradar a las propias bacterias.
En 1917, Félix d'Herelle observó un fenómeno similar en cultivos del bacilo de la disentería y demostró que era posible usar cultivos de este bacilo utilizando filtrados obtenidos a partir de heces fecales. Félix d'Herelle optó inmediatamente por la explicación de que era un virus la causa de este fenómeno y, debido a que el supuesto virus era incapaz de multiplicarse, excepto a expensas de bacterias vivas, D'Herelle decidió llamarlo bacteriófago (devorador de bacterias).
Durante los años veinte la definición del virus se basaba en conceptos puramente negativos: no podían ser vistos al microscopio, no podían ser cultivados en medios artificiales, y además no eran retenidos por los filtros a prueba de bacterias. Sin embargo, las observaciones de D'Herelle permitieron la preparación de grandes cantidades de bacteriófagos (fagos será el término utilizado en adelante), a partir de la lisis de bacterias cultivadas en medios líquidos. Estos lisados podían ser utilizados para infectar otros cultivos. El propio D'Herelle observó que cultivos de bacterias en medio sólido infectados con lisados que contenían fagos, mostraban "claros" o placas en la, por otra parte, uniforme capa de bacterias; el número de estas placas era inversamente proporcional a la dilución del lisado con fagos añadido a dicho cultivo. Por lo tanto, el título de una suspensión de fagos podía ser estimado en términos de unidades formadoras de placas (u.f.p.). De acuerdo con esto, si cada partícula viral presente en la preparación infectante da origen a una placa, entonces la eficiencia de plaqueo (e.p) es igual a la unidad. Muchos años después, este método sería modificado y aplicado al ensayo de los virus de plantas y animales. De hecho, uno de los avances más importantes en el estudio de los virus animales consistió en el ensayo en placa diseñado por Renato Dulbecco en 1952. En este caso, una suspensión de células animales es colocada en una caja de Petri; las células se adhieren y crecen a lo largo y ancho de la superficie de vidrio u otro material sólido, hasta que forman una monocapa de células Entonces se elimina el medio de cultivo y se añade una suspensión viral diluida. Después de una breve incubación que permite que las partículas virales se adhieran (adsorban) a las células, se elimina el exceso de inóculo viral y se añade una capa de agar nutritivo que cubre las células. Después de una incubación que puede durar varios días, las células son teñidas por medio de un colorante vital que solamente penetra en las células vivas, de manera que las muertas por causa de la infección viral permanecen incoloras y se manifiestan como placas desteñidas en la monocapa de células teñidas.
Retomando a D'Herelle y los fagos, este investigador creía que la partícula infectante (fago) se multiplicaba dentro de la bacteria y que su progenie era liberada después de la lisis de la bacteria hospedera. En 1939, Ellis y Delbrück realizaron experimento conocido como "curva de crecimiento en un solo paso", el cual constituyó una sólida prueba en apoyo de la hipótesis de D'Herelle (figura I.1.). Sin embargo, persistía una controversia en cuanto a si el crecimiento intracelular del fago introducía la lisis de la bacteria o en realidad dicha lisis bacteriana era un fenómeno secundario sin relación directa con el crecimiento del fago. Experimentos realizados por Delbrück resolvieron esta controversia y demostraron que en la lisis de las bacterias infectadas participan factores tanto dependientes como independientes de la infección viral. Cuando la infección ocurre a baja multiplicidad, o sea, cuando la relación fago/bacteria no es mayor de 2, el fago penetra la bacteria, se multiplica e induce la lisis de la célula bacteriana. Sin embargo, cuando la infección ocurre en altas multiplicidades, o sea, cuando hay un exceso de partículas infectantes, la lisis bacteriana se debe principalmente a un debilitamiento de la pared celular causado por el exceso de fagos adsorbidos a dicha pared.
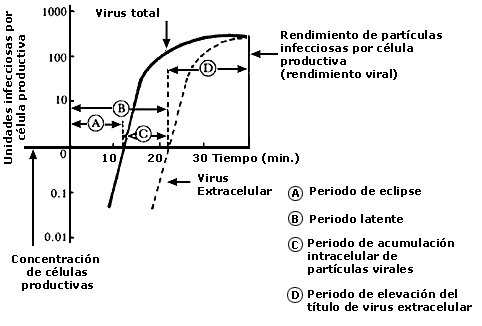
FIGURA I.1. Diagrama de la curva de multiplicación en un solo paso (o escalón)
del bacteriófago T2.
La primera purificación de un virus fue lograda por Max Schlesinger en 1933 utilizando una técnica conocida como centrifugación diferencial, la cual se basa en el hecho de que en un tubo de ensayo sometido a la acción de una fuerza centrífuga las partículas más pesadas sedimentan a menor velocidad que las partículas ligeras. De esta manera es posible separar los fagos de los restos de las células bacterianas. El análisis químico de los fagos purificados demostró que están compuestos por proteínas y ácidos nucleicos en proporciones casi iguales.
d) LA NATURALEZA QUÍMICA Y MOLECULAR DE LOS VIRUS
En 1935 el químico Wendell Stanley pudo aislar el virus causante del mosaico
del tabaco y purificarlo en forma cristalina. Este hecho representó la cristalización
de un material biológico supuestamente vivo y, por lo tanto, tuvo un enorme
impacto desde el punto de vista filosófico, pues llevó al cuestionamiento de
los conceptos establecidos respecto a la naturaleza y propiedades de los seres
vivos. En 1937 Bawden y Pirie lograron la total purificación del virus del mosaico
del tabaco y demostraron que estaba compuesto por proteína y ácido ribonucleico
(ARN). Esto ocurrió en una época en que todavía no se conocía la
naturaleza del material genético. Ya en 1928 Fred Griffith había descubierto
que cepas no patogénicas de neumococos bacterianos, conocidas como cepas R,
podían ser transformadas a la variedad patogénica S cuando eran incubadas en
presencia de un extracto libre de células obtenido a partir de neumococos S
que habían sido "muertos" (o sea, inactivados) por tratamiento térmico. En 1944,
Avery, Mac Leod y Mc Carty informaron que el ácido desoxirribonucleico (ADN)
obtenido a partir de neumococos S, era el único factor capaz de transformar
a los neumococos R y volverlos patogénicos.
Sin embargo, la ortodoxia científica señalaba a las proteínas como posibles
constituyentes del material genético. En aquel entonces era una opinión generalizada
que los ácidos nucleicos carecían de la versatilidad estructural necesaria para
satisfacer el complicado papel atribuido a los hipotéticos genes. Fue en 1952
cuando Hershey y Chase demostraron sin lugar a dudas que la información genética
reside en los ácidos nucleicos. Para esto utilizaron al bacteriófago T2, que
usualmente infecta a la bacteria Escherichia coli. En primer lugar, Hershey
y Chase crecieron el fago en cultivos de E. coli, en los cuales el medio
de cultivo había sido suplementado con azufre o fósforo radiactivos (35S
y 32P). El fósforo radiactivo sirvió para
marcar exclusivamente el ADN presente en los fagos recién sintetizados
por las bacterias infectadas, mientras que el azufre permitió marcar la proteína
asociada con las nuevas partículas virales. Posteriormente, estos fagos radiactivos
fueron utilizados para infectar otros cultivos de E. coli, permitiendo
que los fagos se adsorbieran a las bacterias. Después de una corta incubación,
estas suspensiones de fagos y bacterias fueron sometidas a intensa agitación
para interrumpir la asociación entre fagos y bacterias. La suspensión resultante
fue centrifugada de manera que las bacterias sedimentaron en el fondo de los
tubos de ensayo y las partículas virales permanecieron en el líquido sobrenadante.
Ambas fracciones (sedimento y sobrenadante) fueron analizadas en su contenido
de radiactividad. El resultado indicó que la mayor parte del 32P
había penetrado en las bacterias, mientras que casi todo el 35S
había permanecido en el sobrenadante. Por otra parte, a pesar de este tratamiento,
las bacterias fueron capaces de producir nuevas partículas virales, las cuales
contenían un alto porcentaje de 32P
y casi nada de 35S. Estos resultados
indicaron que solamente el ADN viral penetra en las bacterias y
que este ácido nucleico es suficiente para causar una infección productiva en
dichas bacterias.
La proteína de la cubierta viral no es necesaria para producir la infección debido a que no contiene la información genética. Sin embargo, esta proteína es necesaria para proteger el ácido nucleico y para permitir la adsorción del virus a las células. El experimento mencionado sirvió para demostrar que el ácido nucleido constituye el material genético. (figura I.2.)
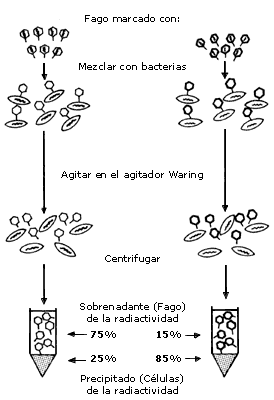
Figura I.2. El experimento de Hershey y Chase.
En 1956, Gierer y Schramm purificaron el ácido nucleico del virus del mosaico
del tabaco (VMT), y demostraron que ARN viral purificado podía
ser infeccioso si se tomaban las precauciones necesarias para protegerlo de
ser inactivado por la acción de enzimas capaces de degradar ácido ribonucleico.
Experimentos previos habían demostrado que las partículas de VMT podían ser
disociadas en proteína y ARN, ambos componentes podían ser reasociados
en el tubo de ensayo para formar partículas virales infectivas por completo
y morfológícamente maduras. Basándose en esta observación, Frankel Conrat y
Singer procedieron a disociar dos cepas diferentes de VMT. El ARN
obtenido de una cepa fue reasociado con la proteína obtenida de la otra cepa
y viceversa, originando partículas híbridas, las cuales fueron usadas para infectar
plantas de tabaco. En todos los casos, las plantas infectadas dieron origen
a progenies virales que correspondían al tipo del ARN presente
en el híbrido infectante (figura 1.3).
Al igual que todos los organismos, los virus pueden generar mutantes en el transcurso de su ciclo de crecimiento; estas mutaciones pueden afectar el tipo de placa formada por la infección viral en los cultivos infectados, el rango de hospederos susceptibles de ser infectados por el virus e incluso las propiedades fisicoquímicas del virus. Un problema asociado con las mutaciones consiste en que varias de éstas pueden ser de tipo letal, o sea, bloquean totalmente la capacidad de replicación del virus y por lo tanto no pueden ser detectadas. Sin embargo, en 1963 Epstein y Edgar descubrieron un tipo muy particular de mutantes virales que ahora son conocidos como "mutantes letales condicionales". Una clase de estos mutantes está constituida por los mutantes sensibles a la temperatura. Estos virus mutantes son capaces de crecer a una cierta temperatura, la temperatura permisiva, pero no pueden hacerlo a una más alta, la temperatura restrictiva misma temperatura que no afecta el crecimiento de virus normales. Otra clase de mutantes condicionales está constituida por los mutantes ámbar, los cuales no pueden crecer en ciertas cepas celulares llamadas no permisivas, mientras que sí pueden hacerlo en cepas celulares permisivas. También se han descrito otros tipos de mutantes condicionales sensibles al frío, los cuales se comportan a la inversa de los mutantes termosensibles anteriormente descritos.
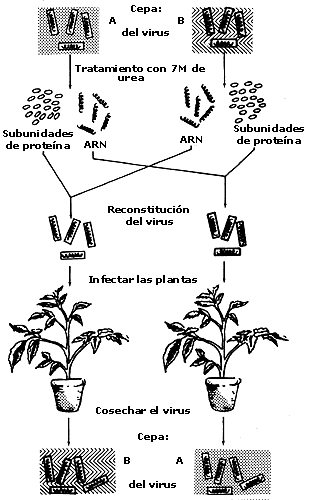
Figura I.3. El experimento de Frankel-Conrat y Singer que demostró que
el ARN es el material genético del virus del mosaico del tabaco
(VMT).
Los virus constituyen partículas extremadamente pequeñas cuyo tamaño varía
más o menos entre 20 y 300 nanómetros (nm).* Por
lo tanto, los virus sólo pueden ser observados bajo el microscopio electrónico.
En muchos casos los virus infectan a la célula hospedera por medio de interacción
directa entre la célula y la partícula viral, pero en otros casos los virus
son transmitidos por medio de un agente animal o vegetal que actúa como vector
intermediario entre el virus y su hospedero final. Las células que hospedan
al virus contienen ambos tipos de ácido nucleico (ADN y ARN),
mientras que los virus contienen solamente un tipo de ácido nucleico, el cual
será ADN o ARN según el tipo de virus en particular.
El virus se reproduce totalmente a partir de su material genético constituido
por el ácido nucleico, mientras que la célula hospedera se reproduce a partir
de la suma integral de sus componentes. El virus nunca se origina directamente
a partir de un virus preexistente, mientras que toda nueva célula se origina
de manera directa de una célula madre. Los componentes de un virus son sintetizados
en forma independiente y son ensamblados posteriormente para formar una partícula
viral madura. En cambio, el crecimiento de las células consiste en un aumento
de todos sus componentes sin que la célula pierda en ningún momento su integridad.
Los virus dependen de la maquinaria metabólica y sintética presente en la célula
hospedera para poder sintetizar el ácido nucleico y las proteínas virales.
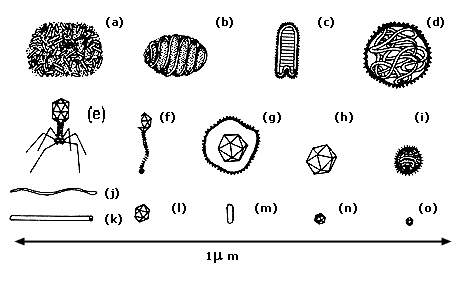
Figura I.4. Diagrama que muestra el tamaño relativo y las formas de diferentes tipos de virus. (a) Poxvirus (vacuna). (b) Poxvirus (dermatitis pustular). (c) Rabdovirus. (d) Virus de la parainfluenza (parotiditis). (e) Bacteriófago. (g) Herpesvirus. (h) Adenovirus. (i) Virus de la influenza. (j) Virus de la papa. (k) Virus del mosaico del tabaco. (l) Polioma/papiloma virus. (m) Virus del mosaico de la alfalfa. (n) Virus de la polio. (o) Fago ØX174.
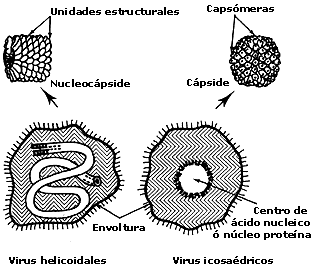
Figura I.5. Diagrama que ilustra la terminología utilizada para describir
los virus con simetría helicoidal (izquierda) y con simetría icosaédrica (derecha).
Existe una terminología bien establecida para describir los diferentes componentes
y estructuras de los virus. Una partícula viral completa e infectiva es denominada
virión. En el caso de los virus con morfología icosaédrica, la cubierta
de proteína es conocida como la cápside que a su vez está compuesta de
unidades morfológicas o capsómeras. La cápside rodea un centro
compuesto de ácido nucleico y proteínas. La cápside y el centro forman en conjunto
la nucleocápside. Ejemplos de virus icosaédricos son el virus de la poliomielitis
y el bacteriófago ØXI74. Otros virus tienen aspecto de bastones o
cilindros. En tales viriones el ácido nucleico está rodeado por una cápside
cilíndrica cuya estructura helicoidal puede ser resuelta bajo el microscopio
electrónico. Ejemplos de viriones helicoidales son el virus del mosaico del
tabaco y el bacteriófago M13. En viriones de morfología más compleja, la nucleocápside
está rodeada por una laxa envoltura membranosa. Dichos viriones tienen
forma relativamente esférica, pero son muy pleomórficos (multiformes) debido
a que la envoltura no es rígida. Ejemplo de virus icosaédricos con envoltura
son los herpesvirus. En el caso de los virus helicoidales con envoltura como
el virus de la influenza, la nucleocápside está enrollada dentro de la envoltura.
En viriones cuya estructura es todavía mas compleja, como en el caso del virus
de la viruela, no es posible identificar una sola cápside, ya que este tipo
de virus tiene varias cubiertas alrededor del ácido nucleico. Las envolturas
virales tienen características similares a las de las membranas celulares debido
a que tales envolturas se derivan de la membrana de la célula hospedera durante
los estadios finales de la infección viral. Por lo tanto, las envolturas virales
son ricas en lípidos (grasas) y proteínas, algunas de las cuales forman complejos
con diferentes tipos de carbohidratos (azúcares), constituyendo las llamadas
glicoproteínas. Los carbohidratos asociados a las proteínas tienden a protuir
de la envoltura viral y a veces es posible reconocerlos bajo el microscopio
electrónico en forma de espigas presentes en la superficie externa del virión.
Las figuras 1.4 y 1.5 dan una idea general de las diversas morfologías y componentes
estructurales de los virus.
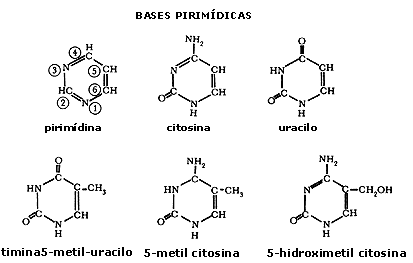
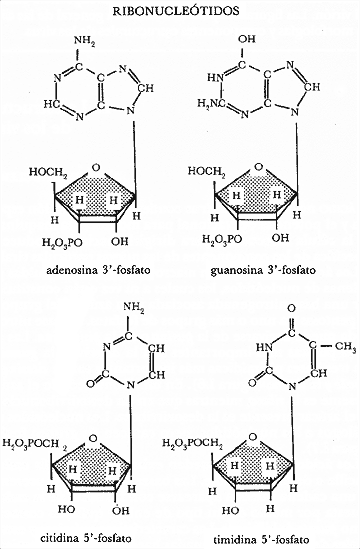
Figura I.7. Estructura de algunos ribonucleótidos.