II. LA ESTRUCTURA DE LOS VIRUS
EL ÁCIDO nucleico de un virus contiene la información específica
y el potencial operacional para modificar la maquinaria de la célula infectada
y para dirigirla hacia la producción específica de los componentes de las nuevas
partículas virales.
Los ácidos nucleicos son macromoléculas constituidas por cadenas de nucleótidos, los cuales a su vez están constituidos por una base nitrogenada asociada a un azúcar del grupo de las pentosas y a uno o más grupos de fosfatos. La base nitrogenada puede derivarse de la purina o de la pirimidina. Las dos bases púricas más importantes son la adenina y la guanina. Las tres bases pirimídicas más importantes son la citosina, el uracilo y la timina (figura 1.6). En un ribonucleótido el azúcar presente es la ribosa, mientras que en un desoxirribonucleótido el azúcar presente es la desoxirribosa. Los nucleósidos son análogos a los nucleótidos, pero carecen de grupos fosfato (figura I.7)
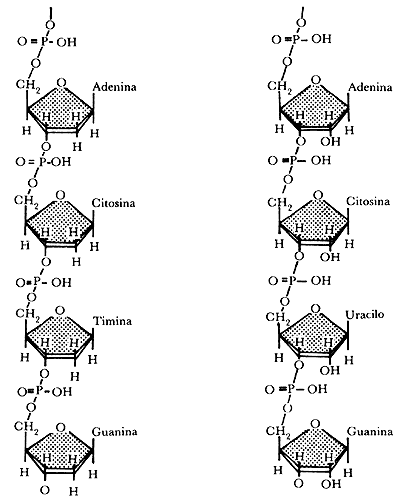
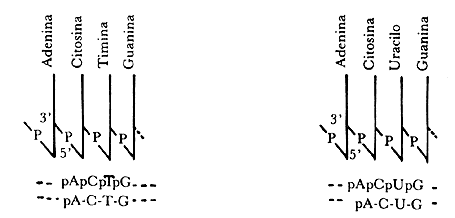
Figura II.1a. Segmentos de una cadena de desoxirribonucleótidos o ADN
(a la izquierda) y de una cadena de ribonucleótidos o ARN (a la
derecha). Las notaciones condensadas son ilustradas junto a cada polinucleótido.
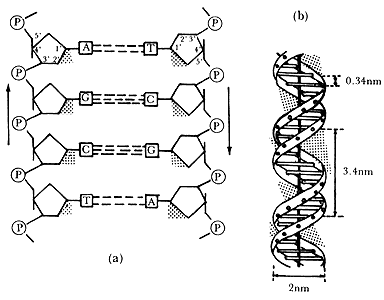
Figura II.1b. Formas de representación esquemática del ADN.
(a) Esquema que muestra la polaridad de las cadenas de desoxirribonucleótidos
y el apareamiento de bases. (b) Esquema que muestra la estructura en doble hélice
y los parámetros helicoidales de la molécula.
Los ácidos nucleicos pueden existir en forma de cadena sencilla o de cadena
doble. Las bases nitrogenadas presentes en una cadena pueden aparearse con las
bases de la cadena opuesta por medio de un tipo de enlace químico conocido como
puente de hidrógeno. Las características químicas y estructurales de
las bases nitrogenadas hacen que el apareamiento ocurra entre la guanina y la
citosina, y entre la adenina y la timina o el uracilo. Generalmente, el ácido
ribonucleico (ARN) es de cadena sencilla, aunque también puede
existir en forma de cadena doble. Las bases normalmente presentes en el ARN
son la adenina, la citosina, la guanina y el uracilo. El ácido desoxirribonucleico
por lo general existe en forma de cadena doble formando la famosa estructura
en doble hélice. Normalmente el ADN contiene las bases adenina,
guanina, citosina y timina (figuras II. (a) y (b)).
Los ácidos nucleicos de cadena doble, como el ADN, pueden ser
desnaturalizados, o sea, sus cadenas pueden ser separadas por medio de un tratamiento
con álcali o por medio de elevar la temperatura, pues los puentes de hidrógeno
son rotos por calor y el pH elevado (alcalino). Si una molécula de ADN
es sometida a desnaturalización térmica, las cadenas complementarias tienden
a separarse conforme se eleva la temperatura. Sin embargo, estas cadenas tenderán
a aparearse de nuevo tan pronto la temperatura desciende a 25°C o menos.
Cuando se añade a esta mezcla de reacción una molécula de ARN,
cuya secuencia es complementaria a cualquiera de las cadenas del ADN
desnaturalizado, es posible obtener híbridos ADN-ARN.
Cuando las macromoléculas del tipo del ARN y el ADN
son sometidas a centrifugación en presencia de una solución concentrada de sales
pesadas como el cloruro de cesio (CsCl), las fuerzas opuestas de sedimentación
y difusión producen un gradiente de concentración de la sal, generando un aumento
continuo de la densidad en dirección de la fuerza centrífuga. Las macromoléculas
presentes en semejante gradiente son impulsadas por la fuerza centrífuga hasta
la región donde la densidad de la solución salina es igual a la densidad de
flotación característica de cada tipo de macromolécula.Cuando existen varias
especies de macromoléculas dentro del gradiente, cada especie formará una estrecha
banda en la posición donde la densidad del CsCl es igual a la densidad de flotación
de la especie molecular en cuestión. Las cinco posibles clases de ácido nucleico:
ADN de cadena doble, ADN de cadena sencilla, ARN
de cadena sencilla, ARN de cadena doble, híbridos ADN-ARN,
pueden ser separadas por medio de centrifugación en gradientes de CsCI. Por
otra parte, es posible introducir marcadores de densidad en los ácidos nucleicos,
haciendo crecer la fuente del ácido nucleico (virus, bacteria, célula, etc.)
en un medio que contenga isótopos pesados de algún elemento que puede ser incorporado
en la estructura de los ácidos nucleicos (15N,
18O, 2H),
o análogos de las bases nitrogenadas como el 5.bromouracilo, cuyo peso molecular
es mayor que el de la timina normalmente presente en el ADN. De
esta manera, el nuevo ácido nucleico tiene una densidad de flotación mayor que
la del ácido nucleico normal equivalente, y esta característica puede ser de
gran utilidad cuando se desea establecer el destino final de una macromécula
en particular.
Existen cuatro posibles tipos de ácido nucleico viral: ADN de
cadena sencilla, ADN de cadena doble, ARN de cadena
sencilla y ARN de cadena doble. Virus que contienen cualquiera
de estos tipos de ácido nucleico pueden ser encontrados tanto entre los fagos
como entre los virus que infectan a plantas o animales. El ADN
de algunos bacteriófagos se caracteriza por contener bases raras que substituyen
alguna o algunas de las bases normalmente presentes en el ADN.
Por ejemplo, el fago PBSl tiene uracilo (normalmente presente en el ARN)
en lugar de timina mientras que los fagos T2, T4 y T6 tienen en su ADN
hidroximetilcitosina en lugar de citosina. La presencia de estas bases raras
permite distinguir con relativa facilidad entre el ácido nucleico viral y aquel
correspondiente a la célula hospedera.
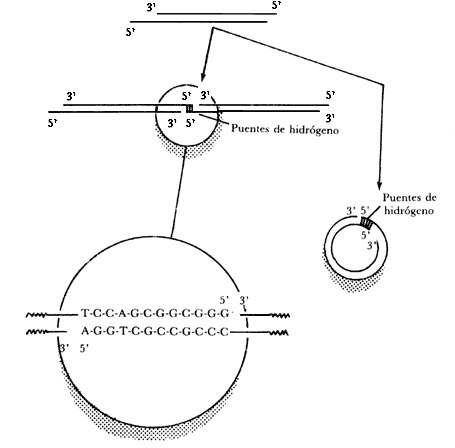
Figura II.2. Formación de dímeros y círculos de ADN del fago
después de una incubación bajo condiciones que favorecen la circulación del
ADN. En la figura se muestran las secuencias de bases que constituyen
los extremos o términos pegajosos.
El ADN de cadena doble presente en algunos virus (como el fago
l), se caractenza por tener segmentos de cadena sencilla en ambos
extremos de la molécula. Debido a que son complementarias las secuencias de
nucleótidos presentes en ambos extremos, resulta posible que entren en contacto
para formar puentes de hidrógeno dando origen a moléculas circulares de ADN
o a dímeros formados por dos moléculas longitudinales de ADN unidas
por sus extremos (figura II.2.). Estos extremos de cadena sencilla presentes
en una molécula de ácido nucleico que por lo demás es de cadena doble, son denominados
como extremos pegajosos o cohesivos.
Cuando es desnaturalizado el ADN de algunos virus, como los adenovirus,
se observa que cada una de las cadenas de ADN es capaz de formar
un círculo por separado. Esto implica que son complementarias las secuencias
de nucleótidos presentes en ambos extremos de cada cadena y por lo tanto deben
tener la misma secuencia, pero repetida en sentido inverso (figura 11.3.). A
este tipo de secuencias se les conoce como repeticiones invertidas. Algunos
virus contienen ácido nucleico que está circularizado en forma natural. Este
tipo de ácido nucleico es insensible a la acción degradante de enzimas como
la exonucleasa III, que tienen la capacidad de digerir moléculas de ácido
nucleico que poseen un extremo libre, lo cual no ocurre en las moléculas circulares.
El ADN naturalmente circular puede ser de cadena sencilla como
en el fago ØXI74, o de cadena doble, como en el virus SV4O. Existe
evidencia de que algunos virus ARN que producen infecciones en
vegetales como el limonero y la papa contienen moléculas circulares de ARN.
La secuencia de los nucleótidos presentes en las cadenas o bandas de los ácidos nucleicos constituye la base del código genético. Cada codón (o letra del código) es definido por una tripleta de nucleótidos que, a través del proceso conocido como traducción es interpretada como una letra que corresponde a uno de los veinte aminoácidos diferentes que constituyen las proteínas. En términos generales, la secuencia de codones que constituyen un gene codifican la secuencia de aminoácidos que constituyen una proteína en particular.
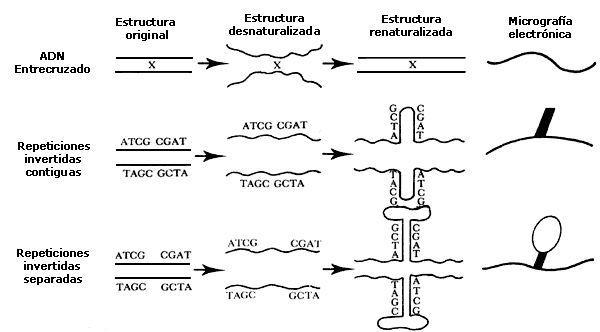
FIGURA II.3. Las secuencias de nucleótidos que corresponden
a repeticiones invertidas permiten que se formen regiones de cadena doble debidas
a reasociación intramolecular. Cuando las repeticiones se encuentran muy próximas
entre sí, se forma una horquilla doble con extremo de cadena sencilla. La última
columna en el diagrama muestra la apariencia de estas estructuras producidas
por la reasocación de las repeticiones invertidas, cuando son observadas al
microscopio electrónico: las regiones de cadena doble se distinguen por su mayor
grosor
En los últimos diez años se han desarrollado una variedad de técnicas y métodos
que permiten determinar la secuencia de nucleótidos en cualquier tipo de ácido
nucleico. La primera secuencia completa de un ARN viral fue determinada
en el fago MS2 por el grupo de Walter Fiers en 1976. En 1977, Fred Sanger y
colaboradores publicaron la secuencia completa del genoma del fagoØXl74,
constituido por ADN de cadena sencilla. Posteriormente, muchos
otros genomas virales de mayor tamaño y complejidad han sido secuenciados en
parte o en su totalidad. Una vez que se conoce la secuencia del genoma viral
es posible establecer la forma como están organizados los genes presentes en
el ácido nucleico. Los avances de la biología molecular han permitido determinar
la naturaleza de las secuencias de nucleótidos que actúan como signos de puntuación
en la lectura de la información genética. Normalmente en los genomas de bacterias,
plantas y animales cada gene abarca uno o varios segmentos del ácido nucleico,
estos segmentos están separados de los segmentos correspondientes a los genes
adyacentes. En el caso de los virus, los genomas virales resultan ser muy pequeños
cuando se les compara con los genomas de las bacterias más simples; esto implica
que los virus tienen una capacidad muy limitada para contener información genética.
Sin embargo, algunos virus han desarrollado estrategias para obtener una máxima
capacidad de almacenamiento de la información genética. Una de estas estrategias
consiste en la traslapación de genes, de manera que un segmento del ácido nucleico
puede contener la secuencia de nucleótidos correspondiente a la totalidad del
gene A y a la vez contener la secuencia inicial correspondiente al gene B, mismo
que se continúa en otra región del ácido nucleico posterior al término del gene
A. La otra estrategia consiste en la superposición de genes, de manera que el
segmento del ácido nucleico que corresponde al gene C incluye también al gene
D que codifica una proteína más pequeña que la codificada por el gene C. Esta
multiplicidad de marcos de lectura característica de algunos genomas virales
implica la necesidad de una compleja regulación y coordinación de la expresión
genética en esos virus (figura II.4.).
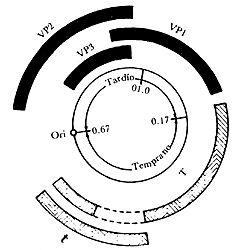
Figura II.4. Mapa genético del ADN del virus SV40 que contiene
5 243 pares de bases. La región temprana (transcrita en el sentido opuesto de
las manecillas del reloj) es ilustrada en gris, y la región tardía (transcrita
en el sentido de las manecillas del reloj) es ilustrada en negro. El origen
de replicación es marcado Ori.
b) LA FUNCIÓN PROTECTORA Y MORFOLÓGICA DE LAS PROTEÍNAS VIRALES
El análisis de partículas virales purificadas muestra que con tienen entre 50 y 90% de proteína. Si consideramos que los ácidos nucleicos en solución son susceptibles de ser fácilmente fragmentados o degradados, podemos asumir que el componente proteico de los virus tiene fundamentalmente un papel protector.
Una tripleta de nucleótidos (codón) tiene un peso molecular promedio de alrededor de 1000 daltones y sólo codifica un aminoácido cuyo peso molecular promedio es de aproximadamente 100 daltones. Por lo tanto, un ácido nucleico puede especificar cuando mucho una décima parte de su peso molecular en proteína. Con mucha frecuencia los virus contienen más de un 50% de su peso en forma de proteína; esto sugiere que deben estar presentes varias copias de una misma proteína de bajo peso molecular, ya que se requiere de menos material genético para especificar un solo tipo de molécula de proteína la cual puede ser usada como subunidad para construir la cubierta del virus. Sin embargo, no es esencial que la cubierta del virus esté formada por subunidades idénticas, siempre y cuando los pesos moleculares combinados de los diferentes tipos de subunidades sean suficientemente pequeños en relación con el peso molecular de la molécula del ácido nucleico viral. La construcción de las partículas virales a partir de subunidades estructurales incrementa la estabilidad genética del propio virus, ya que al reducirse el tamaño de las subunidades estructurales se reduce la posibilidad de que ocurran mutaciones nocivas en el gene que codifica dicha subunidad.
El fenómeno de autoensamble es particularmente relevante para los virus y otros sistemas biológicos debido a sus atributos de economía y eficiencia. Por ejemplo, en la industria de la construcción ha sido posible abatir los costos e incrementar la eficiencia utilizando unidades prefabricadas que a su vez son ensambladas en forma rápida y barata en el sitio de construcción. Esto involucra dos procesos: la fabricación de unidades básicas y el ensamble de las mismas para formar edificaciones complejas. En el caso de los sistemas biológicos, la fabricación de unidades es análoga a la síntesis de moléculas de proteína a partir de aminoácidos. Esta síntesis depende de las instrucciones contenidas en la secuencia de nucleótidos presentes en los ácidos nucleicos. El proceso de ensamble es independiente de instrucciones externas ya que la información necesaria para construir los complejos moleculares está incluida dentro de los propios componentes individuales. El mecanismo de autoensamble tiene la ventaja de que puede ser controlado en cada nivel de organización. Por ejemplo, las subunidades defectuosas son eliminadas automáticamente durante el proceso de ensamble, de manera que estructuras complejas son construidas con exactitud y eficiencia. En el caso particular de los virus, las moléculas de proteína que constituyen las subunidades de la cápside interaccionan con la cadena o cadenas del ácido nucleico viral para formar una partícula viral. El proceso depende de la formación de enlaces entre las subunidades y al igual que en el proceso de cristalización, la regularidad de la estructura final es consecuencia de factores termodinámicos que obligan a formar un máximo número de uniones no covalentes entre las subunidades. Sin embargo, en un cristal todas las moléculas se encuentran en ambientes similares, mientras que tal situación rara vez ocurre en el caso de estructuras hechas a partir de subunidades de proteína. Una proteína es una macromolécula compuesta por una variedad de moléculas individuales denominadas aminoácidos. Los aminoácidos se encuentran unidos por enlaces químicos conocidos como uniones o enlaces peptídicos; por lo tanto, toda proteína puede ser considerada como una cadena polipeptídica, la cual tendrá una conformación espacial o estructura terciaria que depende directamente de la composición de los aminoácidos presentes en dicha proteína. Debido a su composición química; los aminoácidos se dividen en neutros, hidrofóbicos e hidrofílicos. Así, cuando una proteína se encuentra rodeada completamente por un medio acuoso, los grupos hidrofóbicos tienden a juntarse en el interior de la molécula de proteína, mientras que los grupos hidrofílicos tienden a permanecer en la superficie de la molécula, haciendo contacto con el medio. Estas interacciones determinan la conformación espacial de la proteína.
En el caso del virus del mosaico del tabaco (VMT), una proteína compuesta por 158 aminoácidos constituye la subunidad básica a partir de la cual se construye la cápside del virus. En dicha proteína cuando menos la mitad de los aminoácidos presentes en el interior de la macromolécula son de tipo hidrofóbico, mientras que en la superficie de la misma hay tan sólo cuatro grupos hidrofóbicos en un segmento constituido por 24 residuos de aminoácidos. Por lo tanto, se puede decir que los mismos principios fisicoquímicos que rigen el desarrollo de la estructura terciaria y cuaternaria de las proteínas son causa de la organización de las cápsides virales. Las múltiples uniones formadas durante la agregación de las subunidades de proteína contribuyen al enmascaramiento de sitios potencialmente susceptibles a la acción de enzimas capaces de degradar proteínas; de esta manera las proteínas de la cápside viral adquieren también mayor resistencia al calor y otros agentes físicos.
Es bien sabido que las suspensiones de partículas virales pueden ser mantenidas por largos periodos en el laboratorio esto implica que dichas partículas constituyen estructuras estables. Una condición necesaria para lograr la estabilidad de cualquier estructura consiste en que la estructura se encuentre en su estado de mínima energía libre; esto se logra estableciendo un máximo número de uniones entre las subunidades que constituyen dicha estructura. Debido a que las subunidades de la cubierta viral son relativamente asimétricas, se requiere que estén dispuestas o ensambladas en forma simétrica para que pueda formarse el mayor número de uniones entre dichas subunidades. Existe un número limitado de formas que permiten el ensamble simétrico de subunidades asimétricas.
Una posible forma de generar una estructura simétrica tridimensional a partir
de componentes asimétricos como las proteínas consiste en distribuir las proteínas
alrededor de una circunferencia de manera que formen un disco. Apilando un gran
número de discos se obtiene una estructura tridimensional con una cavidad interna
apropiada para albergar la molécula del ácido nucleico (figura II.5.). Un ejemplo
de virus filamentosos está dado por el virus del mosaico del tabaco. El examen
detallado del VMT muestra que las subunidades de proteína no están dispuestas
en forma de anillos sino en forma helicoidal. Esto resulta lógico considerando
que el ARN del VMT tiene una forma helicoidal y, por lo tanto,
al disponer las subunidades de proteína en forma helicoidal es posible formar
el mayor número de uniones entre las subunidades de proteína y el ácido nucleico.
Todos los virus filamentosos estudiados hasta la fecha muestran una estructura
helicoidal indicando que el ácido nucleico es el factor esencial que rige este
tipo de arreglo estructural.
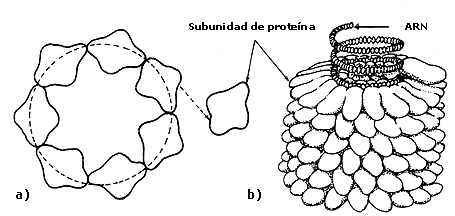
Figura II.5.(a) Disposición de componentes asimétricos idénticos alrededor
de una circunferencia para lograr una estructura simétrica. (b) Segmentos de
una partícula de virus de mosaico del tabaco que muestra las subunidades de
proteína formando una estructura helicoidal. El ARN se localiza
en un surco helicoidal formado por las subunidades de proteína.
También se puede obtener una partícula simétrica disponiendo las subunidades de proteína alrededor de los vértices o caras de un cuerpo con simetría cúbica como el icosaedro, constituido a partir de 20 triángulos equiláteros. Multiplicando el número de subunidades presentes en cada cara por el número de caras, obtenemos el número mínimo de subunidades que pueden ser acomodadas alrededor de tal cuerpo geométrico. En el caso del icosaedro, el número es de 60 subunidades. Este tipo de arreglo representa una de las pocas formas en que objetos asimétricos pueden ser acomodados en forma simétrica sobre la superficie de una esfera. Las micrografías electrónicas de un gran número de virus diferentes muestran que éstos tienen una silueta esférica que al ser examinada más detalladamente corresponde a una estructura con simetría icosaédrica. Varios de los virus esféricos con simetría icosaédrica contienen más de 60 subunidades de proteína. Por ejemplo los adenovirus contienen aproximadamente 1 500 subunidades en su cubierta (figura II.6.). Sin embargo, Donald Caspar y Aaron Klug han descrito las reglas que gobiernan la estructura de los virus esféricos. Tales reglas fueron inspiradas por el estudio de las estructuras geodésicas diseñadas por el arquitecto norteamericano Buckminster Fuller. En dichas estructuras la superficie de una esfera es subdividida en facetas triangulares, las cuales son arregladas con una simetría icosaédrica. Este método de triangulación de la esfera representa el diseño óptimo para una cubierta cerrada construida a partir de subunidades idénticas unidas en forma regular. Ninguna otra forma de subdividir una superficie cerrada puede proporcionar un grado similar de equivalencia. Este tipo de estructuras tienen un mínimo de energía libre, lo cual explica en parte la abundancia de los virus con simetría icosaédrica.
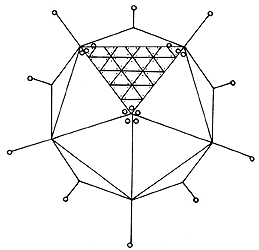
Figura II.6. Organización de las subunidades de proteína en la cápside del
adenovirus.
e) VIRUS CON MÁS DE UN TIPO DE SUBUNIDAD
Las micrografías electrónicas de los adenovirus muestran que las 1 500 subunidades de proteína están arregladas en 240 hexámeros y 12 pentámeros, y en cada vértice del virus se proyecta una fibra de proteína. Está bien establecido que las fibras, los pentámeros y hexámeros están constituidos por diferentes tipos de proteínas. Los adenovirus representan el ensamble regular de tres diferentes tipos de proteína. Esto puede lograrse arreglando los pentámeros y las fibras en los vértices del icosaedro, y los hexámeros en las caras del icosaedro. Una forma diferente de arreglo estructural ocurre en los reovirus en los cuales la cápside está constituida a partir de 8 diferentes proteínas dispuestas en dos capas o cubiertas que poseen simetría icosaédrica. Tres de las proteínas están dispuestas en la cubierta externa, y las cinco proteínas restantes en la cubierta interna.
Muchos de los virus animales más grandes y unos cuantos virus de plantas y bacterias están envueltos por una cubierta membranosa de proximadamente 75Å de grosor. Esta envoltura se deriva en gran parte de las membranas de la célula hospedera y puede ser degradada por medio del tratamiento con detergentes o solventes orgánicos como el éter, ocasionando así la pérdida de la inefectividad del virus. A este tipo de virus también se les conoce como virus sensibles al éter. Los virus de la influenza son representativos de los virus animales con envoltura. Estos virus han sido descritos como pleomórficos (multiformes), aunque las partículas virales en células de animales vivos tienen una forma filamentosa. La apariencia pleomórfica se hace evidente cuando estos virus son cultivados en embriones de pollo. Estos virus poseen debajo de la envoltura una capa o cubierta de proteína llamada proteína M (proteína de la membrana o proteína de la matriz). Dentro de la cubierta formada por la proteína M, pero separado de la misma, se encuentra el componente nucleoproteico constituido por un cilindro flexible de ARN y proteína dispuesta en forma similar a un pasador para el pelo que ha sido doblado. La estructura de la nucleocápside del virus de la influenza es de tipo helicoidal. Cada uno de los diferentes virus de la influenza codifica cuando menos 4 diferentes proteínas que son ensambladas en el virión, dando origen a diferentes estructuras virales.
Algunos virus con envoltura tienen nucleocápsides icosaédricas. Particularmente
los virus ARN productores de tumores también conocidos como retrovirus,
tienen una nucleoproteína que está superenrollada en forma de una esfera hueca
rodeada por la envoltura, la cual se deriva de la membrana celular y es modificada
por la inserción de glicoproteínas específicas del virus.
g) VIRUS CON MORFOLOGÍA DE CABEZA Y COLA
Este tipo de morfología sólo ha sido encontrada en cierto tipo de bacteriófagos y está directamente relacionada con la forma en que tales virus infectan a sus bacterias hospederas. El número de fagos con arquitectura de tipo cabeza-cola es considerable; estos fagos pueden ser subdivididos en fagos de cola corta, fagos de cola larga no contráctil, y fagos con colas contráctiles y complejas que también pueden poseer otro tipo de estructuras como collares, placas basales y fibras (figura II.7.). A pesar de su complejidad estructural, los principios que gobiernan el ensamble de estos fagos son similares a los descritos en relación con otros virus cuya arquitectura es más sencilla. Las cabezas de los fagos tienen usualmente una simetría osaédrica mientras que las colas tienen una simetría helicoidal.
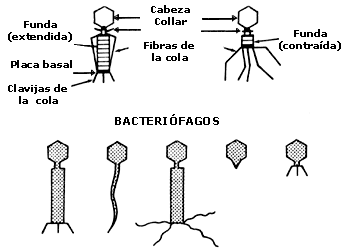
Figura II.7. Representación esquemática de las estructuras de algunos bacteriófagos
con cola.
Todas las estructuras presentes, como las placas basales poseen alguna forma de simetría.
h) EL PRINCIPIO DE AUTOENSAMBLE
Los virus son construidos de acuerdo con principios de diseño eficientes y,
por lo tanto, son capaces de autoensamblarse sin la participación de ningún
factor organizador externo. Esto es posible debido a la formación de un gran
número de uniones débiles cuando los componentes del virus son colocados en
la configuración adecuada por medio de movimientos al azar propiciados por factores
termodinámicos. Por ejemplo, si se trata al VMT con una solución concentrada
de urea, ocurre una descomposición del virus en subunidades de proteína y ARN.
Si se elimina la urea y son incubadas de nuevo las subunidades de proteína en
presencia del ARN viral, las macromoléculas se agregan en forma
espontánea dando origen a partículas virales infecciosas. Sin embargo, cuando
se omite el ARN viral durante el proceso de repolimerización, se
obtienen cilindros de proteína, pero en este caso las subunidades están apiladas
en forma de discos en lugar de tener una disposición helicoidal. Además, estos
cilindros son menos estables que la particula viral completa, lo que señala
la importancia del ácido nucleico en la estabilización de la estructura viral.
En el caso de algunos virus icosaédricos, ha sido posible realizar también experimentos de reconstrucción o reconstitución in vitro (o sea, en el tubo de ensayo). Sin embargo, modificando las condiciones de estos experimentos, ha sido posible obtener estructuras tubulares y otras variables morfológicas. Esto indica que las propiedades de empacamiento de las proteínas son las que en última instancia determinan la estructura de la partícula viral.
El autoensamble resulta económico para los virus, pues no requieren de información
genética específica para lograrlo, y además proporciona un método sencillo y
eficiente para eliminar subunidades defectuosas producidas durante la replicación
de los componentes virales. Sin embargo, algunos bacteriófagos complejos del
tipo cabeza-cola, como el fago T4, muestran cierto grado de control genético
en el proceso de ensamblaje; este fenómeno será discutido más adelante.