IV. LISOGENIA
POCO tiempo después de que fueron descubiertos los fagos, se aislaron cepas bacterianas que parecían ser portadoras silenciosas de ciertos tipos de fagos. Los líquidos obtenidos a partir de cultivos de estas cepas portadoras mostraban la presencia de fagos; sin embargo, las cepas portadoras no eran sensibles a ser destruidas por el fago que portaban. Por otra parte, cepas de bacterias emparentadas con la cepa portadora resultaban ser sensibles al fago presente en las cepas portadoras. Las cepas de bacterias portadoras de fagos silenciosos o latentes fueron denominadas cepas lisogénicas. A principios de los años cincuenta se descubrió que los fagos son capaces de adsorberse a las bacterias lisogénicas, pero no se produce la subsecuente lisis de estas bacterias. Por otra parte, cuando el fago procedente de una cepa lisogénica es sembrado en una cepa de bacterias sensibles, se pueden aislar en estos cultivos colonias de bacterias que se comportan igual que las cepas lisogénicas. En 1950, André Lwoff cultivó una sola célula procedente de una cepa lisogénica de Bacillus megaterium y observó bajo el microscopio la división de esta bacteria. Posteriormente, Lwoff removió una de las células hijas junto con un poco del medio de cultivo. Este proceso fue repetido varias veces: cada vez que se dividía la célula remanente, era removida una de las células hijas y algo del medio de cultivo; las células hijas y el viejo medio de cultivo fueron sembrados en agar para determinar si daban origen a una población de bacterias lisogénicas y a la presencia de fago en el medio de cultivo. Estos experimentos mostraron que las bacterias lisogénicas pueden crecer y dividirse sin liberar fagos al medio de cultivo. Sin embargo, por alguna razón desconocida, algunos filtrados obtenidos a partir de extractos de bacterias lisogénicas mostraban la presencia de partículas virales, o sea, fagos. Lwoff razonó que en las cepas lisogénicas el fago se encuentra en forma de un precursor no infeccioso al que denominó profago. La lisis de algunas de estas bacterias lisogénicas ocurre solamente cuando estas células han sido estimuladas para producir fagos. Lwoff y colaboradores observaron que la irradiación con luz ultravioleta (U.V.) era capaz de inducir la producción de fagos en una población de bacterias lisogénicas, mismas que eran lisadas en la medida que se incrementaba la concentración de fagos liberados al medio de cultivo. Por lo tanto, una bacteria lisogénica posee la capacidad de heredar el fago a sus descendientes, muy pocos de los cuales se lisarán en forma espontánea. Sin embargo, la mayor parte de la progenie de una bacteria lisogénica puede ser inducida a producir el fago por medio de la irradiación con U.V. o tratamiento con otros factores inductores. Se denomina temperados a los bacteriófagos capaces de existir en forma de profago en el interior de una bacteria hospedera. Después de que el profago ha sido inducido por irradiación, ocurre un breve periodo de eclipse en el cual no se puede detectar la presencia del fago dentro de la bacteria. Sin embargo, es posible detectar la aparición de proteínas y ácido nucleico específicos del fago; estos elementos serán ensamblados para formar los nuevos fagos maduros poco antes de que ocurra la lisis de la bacteria hospedera.
En 1951, Esther Lederberg descubrió en forma accidental que la cepa de E. coli K12 era de tipo lisogénico. El fago latente en dicha cepa fue aislado al mezclar E. coli Kl2 con derivados no lisogénicos de esta cepa bacteriana. El fago resultante es ahora conocido como fago y representa el caso más estudiado del fenómeno de lisogenia.
Las bacterias infectadas por fagos temperados continúan dividiéndose por varias generaciones y son inmunes o resistentes a ser superinfectadas por el mismo tipo de fago que albergan o por otros fagos pertenecientes a clases emparentadas con el fago temperado original. Estas bacterias contienen cuando menos una copia íntegra del genoma del fago. Las bacterias infectadas por fagos temperados portan la información genética correspondiente al fago, a través de múltiples divisiones bacterianas, o sea, el genoma del fago es replicado al mismo tiempo que ocurre la replicación del genoma bacteriano; este hecho hace posible que la bacteria original pueda heredar el fago temperado a la subsecuente progenie bacteriana.
El genoma del fago l está constituido por una molécula
lineal de ADN de cadena doble; esta molécula posee extremos cohesivos
o "pegajosos" debidos a la secuencia de nucleótidos presentes en esa región
del genoma viral. Al penetrar en la bacteria, el ADN del fago l
se circulariza por la interacción entre los extremos cohesivos de la molécula.
Ocasionalmente, este anillo de ADN puede ser integrado en el cromosoma
de la bacteria. La integración ocurre en un sitio específico del genoma bacteriano;
en ese sitio existe una secuencia de nucleótidos que resulta homóloga a la de
una región del genoma del fago denominada att l
. Cuando en forma espontánea se alinean paralelamente estas secuencias homólogas,
el círculo del genoma del fago se rompe y la integración se lleva a cabo por
medio del entrecruzamiento entre el ADN del fago y el ADN
de la bacteria; este entrecruzamiento es mediado por un factor proteico codificado
por un gene del fago (int). El fago, una vez integrado, se encuentra
como profago y solamente unos cuantos de sus genes son expresados (figura IV.1).
Un gene en particular produce una proteína que se comporta como un represor
que inactiva la expresión de los otros genes virales. Este represor también
actúa como un factor de inmunidad que im pide la superinfección de la misma
bacteria por otro fago l .
Un factor crítico para el mantenimiento del estado de profago está representado
por la concentración de la proteína represora en el citoplasma bacteriano. Eventos
que disminuyen la concentración del represor inducen la expresión del genoma
viral completo. Un ejemplo de lo anterior ocurre cuando una bacteria lisogénica
masculina, o sea, poseedora del factor de conjugación F (bacteria F+), se conjuga
con una bacteria no lisogénica femenina (F-) introduciendo en el citoplasma
de la bacteria femenina un fragmento de ADN que incluye al profago.
Debido a que en el citoplasma de la célula receptora no existen moléculas del
represor para el fago l, se produce una rápida inducción
de la expresión del genoma viral y la subsecuente lisis de la bacteria receptora.
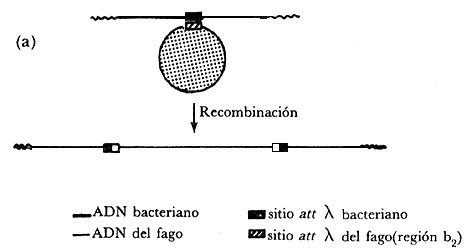
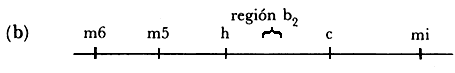
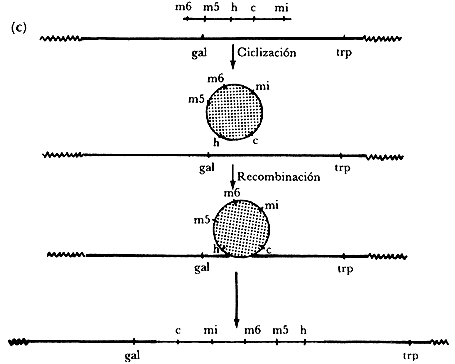
Figura IV. 1.(a) Modelo de Campbell para la inserción del ADN
del fago en el cromosoma bacteriano. (b) Mapa genético del fago. (c) Reordenamiento
de los genes del fago después de inserción del ADN del fago en
el cromosoma bacteriano. Los símbolos gal y trp representan los
operones de la galactosa y del triptófano, respectivamente.
La transición entre el estado de profago y la infección productiva de tipo
lítico inducen que el ADN viral se separe del cromosoma bacteriano
por medio de la formación de una asa de ADN viral, la cual es cortada
y liberada del cromosoma por mediación de proteínas codificadas por los genes
xis e int del fago. Una vez liberado el genoma viral, los extremos
cohesivos se reúnen formando una molécula circular de ADN viral.
Algunas veces ocurren errores en el proceso de corte o escisión, de manera
que el ADN liberado del cromosoma incluye pequeños segmentos correspondientes
al ADN bacteriano adyacente al profago. Por ejemplo, el ADN
bacteriano presente a la izquierda del profago se incluye secuencias correspondientes
de los genes del operón de la galactosa, los cuales codifican enzimas involucradas
en el metabolismo y procesamiento del carbohidrato galactosa. Generalmente,
el error en la escisión, mismo que resulta en la inclusión de una porción del
genoma bacteriano, también ocasiona una pérdida recíprocamente proporcional
del genoma viral. Por ejemplo, cuando por error se incluyen los genes del operón
de la galactosa en el ADN liberado, se pierden genes normalmente
presentes en el extremo derecho del genoma del fago. El genoma viral defectuoso
sirve tomo templado para la replicación del ADN, de manera que
los nuevos fagos tendrán genes para la utilización de la galactosa y a la vez
carecerán de algunos genes virales que salieron perdidos durante el proceso
de escisión. Cuando se infectan con este fago defectuoso bacterias mutantes
que tienen un defecto en el operón de la galactosa (bacterias Gal-), en condiciones
favorables al proceso de lisogenización, es posible que la inserción del genoma
viral en el cromosoma de la bacteria mutante resulte en la adquisición de las
funciones para la utilización de la galactosa, previamente ausentes en la bacteria
mutante. A este fenómeno se le conoce como transducción y los fagos capaces
de inducirlo son denominados fagos transductantes, los cuales tienen la capacidad
de introducir en las bacterias infectadas fragmentos de ADN que
codifican funciones previamente ausentes en esas bacterias. En general, los
fagos transductantes son defectuosos en su propia replicación debido a que carecen
de ciertos genes virales necesarios para este proceso. Por lo tanto, estos fagos
defectuosos sólo pueden propagarse en presencia de fagos normales (auxiliares)
capaces de proporcionar los factores codificados por la región del ADN
ausente en los fagos defectuosos.