IV. EN QUÉ SE GASTA LA ENERGÍA
TENEMOS ya claros los mecanismos de las transformaciones de la
energía, de lo que podríamos llamar sus formas generales, como la energía luminosa
o la de los enlaces químicos de las moléculas, en formas propias de los seres
vivos: la del poder reductor o la de los enlaces del ATP entre sus dos fosfatos
últimos, o las diferencias de concentración de sustancias neutras o cargas a
ambos lados de algunas membranas. Otro de los grandes capítulos de la bioenergética
consiste en el análisis de una gran variedad de cambios en los que estas formas
de energía "participan" en ciertos procesos, para permitir su curso y constituir
así la vida misma.
Ha quedado también ya claro que las formas de la energía de los seres vivos no son diferentes de otras, y que se les puede analizar y cuantificar mediante los conceptos más generales de la física o la fisicoquímica. Aunque en esta época es del dominio común que no hay una "fuerza vital" especial presente en los fenómenos relacionados o que constituyen la base de la vida, es interesante conocer con cierto detalle los componentes que los seres vivos tienen en particular para realizar las transformaciones de la energía.
En los capítulos anteriores revisamos los mecanismos mediante los cuales los seres vivos transforman la energía del exterior, la luminosa o la de los alimentos, en otras formas directamente aprovechables o utilizables por nuestros propios sistemas biológicos. Resta, sin embargo, describir ahora los mecanismos o lo que podríamos llamar el "destino", la finalidad de esas formas de energía, o más correctamente, el cambio de la energía en otras formas que, en conjunto, representan la vida misma de los organismos.
LA SÍNTESIS Y LA RENOVACIÓN DE LAS MOLÉCULAS
Aunque es real, no todos tenemos claro que la totalidad de nuestros componentes está en continua renovación. Éste es un concepto relativamente nuevo, que si bien era claro en el caso de algunos de los tejidos que nos componen, era difícil de imaginar para otros.
Hacia finales del siglo pasado, el científico francés Claude Bernard estableció que en los organismos vivos había sustancias que se podían, por así decirlo, "gastar" o desaparecer. Con base en experimentos realizados en animales a los que se les privaba del alimento, encontró que algunos de los componentes desaparecían antes de que se produjera la muerte; otros, por el contrario, parecían persistir, y si éstos desaparecían, se producía la muerte. Ante estos datos experimentales se propuso, y se aceptó, que había en los organismos vivos un "elemento variable" y uno "constante". Por lo demás, parecía lógico pensar que algunos componentes eran perennes; esto era más fácil en el caso del sistema nervioso, por ejemplo.
Sin embargo, con el descubrimiento de los isótopos de los átomos, que son elementos con un núcleo más pesado, y algunos de ellos radiactivos, el primero de los cuales fue el deuterio, se encontró, por principio de cuentas, que era posible introducirlo prácticamente en cualquier molécula del organismo que contuviera hidrógeno en forma natural. Pero, además, se descubrió que una vez marcadas las moléculas, esta marca desaparecía con una velocidad que no era la misma para todos los componentes, como grasas, azúcares o proteínas, y que también era variable para componentes iguales o semejantes. Si el estudio se hacía en diferentes órganos marcando una proteína, la albúmina por ejemplo, ésta aparecía y desaparecía con mayor rapidez en el plasma sanguíneo que en el hígado de los animales.
Lo más interesante de los descubrimientos realizados era tal vez que ninguna sustancia química estudiada escapaba a esta renovación o recambio constante; parecía (y puede parecer aún) que esto es un gran desperdicio de energía diseñado por la naturaleza. De cualquier manera, no hubo más remedio que aceptar la realidad: los seres vivos invierten gran cantidad de materiales obtenidos del exterior y de la energía derivada de su degradación en mantener este continuo recambio de sus componentes.
No hay estudios detallados, y serían difíciles de realizar, sobre la velocidad con que se renuevan todos los distintos componentes de cada órgano o tejido de los organismos que existen en el planeta. Esto es explicable; todavía no describimos siquiera la totalidad de los componentes de uno solo de los organismos vivos, menos aún podríamos saber la velocidad con que individualmente se renueva cada uno de ellos. No obstante sí es posible saber en términos generales en qué se gasta la energía al renovarlos.
Cuando se sintetizan las moléculas grandes, es decir, cuando se forman a partir
de otras más pequeñas, requieren de la formación de enlaces nuevos entre estas
últimas. Una proteína se forma de aminoácidos que se unen unos con otros; una
grasa implica la unión de ácidos grasos con glicerol y otros componentes; un
polisacárido como la celulosa, el almidón o el glucógeno se forma por la unión
de muchas, muchísimas moléculas individuales de glucosa (Figura IV.I). Cada
uno de los nuevos enlaces que se forma implica, por así decirlo, la inyección
o la administración de una cierta cantidad de energía, que resulta o proviene
de la de otros enlaces. Si ya mencionamos que los enlaces químicos del ATP y
el poder reductor son los modos principales en que se transforma la energía
que existe en otros compuestos químicos, es fácil ver que para todos estos procesos
de síntesis hay presentes complicados mecanismos que implican, a final de cuentas,
la transformación de la energía química de los enlaces del ATP en la propia
de los enlaces de esas grandes moléculas de las cuales hay miles diferentes
en nuestro organismo o en el de cualquier otro animal.
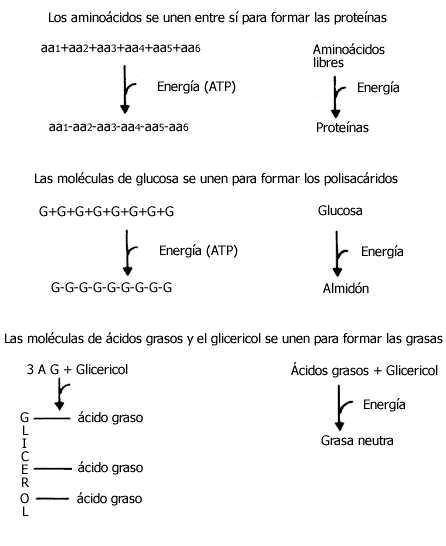
Figura IV.1 Las moléculas más simples se unen unas a otras para formar las
más grandes, en un proceso que requiere energía que, directa o indirectamente
proviene del ATP. Cuando las moléculas grandes se rompen (hiabolizan) para dar
sus componentes, la energía de sus enlaces se transforma en calor.
La figura IV.2 muestra el proceso que tiene lugar ahora para la degradación de estas mismas moléculas. La ruptura en sus componentes ocurre por la simple introducción de una molécula de agua en los enlaces intermedios. Pero la energía que había en ellos no es utilizable; al romperse las uniones, esa energía química se transforma en calor.

Figura IV.2 Cuando se rompen las moléculas más grandes para dar las unidades
que las componen, la energía utilizada en sintetizarlas se transforma en calor.
Es así que para sintetizar o formar las moléculas grandes, sus enlaces se forman a expensas de la energía contenida en los del ATP. Las células deben entonces "gastar" energía, que a su vez proviene de otras formas, la luz en un principio, o los alimentos y sus componentes. Al degradarse esas moléculas, la energía se transforma o se "disipa" en forma de calor. El balance es a final de cuentas que una gran parte de la energía que se requiere para llevar a cabo esta constante renovación de sus componentes está contenida en los alimentos de los seres vivos o de otras formas externas aprovechables por ellos.
Los organismos que estamos acostumbrados a ver en la vida diaria, incluso nosotros mismos, tienen la necesidad de tomar del exterior sustancias para vivir. Los animales necesitan alimentos muy diversos que contienen a su vez una gran variedad de sustancias, y agua. Las plantas necesitan agua y algunas sustancias que, o bien pueden generarse en el mismo suelo, o debemos proporcionarles, como los fertilizantes, que no son otra cosa que diferentes tipos de sales de amonio y otros minerales. Los organismos unicelulares, como las bacterias o los hongos unicelulares, también deben tomar del medio en que viven materiales que les son indispensables para subsistir. Pero todos los seres vivos están rodeados de cubiertas protectoras que los aislan del medio e impiden la pérdida de los materiales de que están compuestos. Estas cubiertas protectoras van desde la piel de los animales y la corteza de los árboles, hasta la membrana de las células individuales, y los materiales que requerimos del exterior deben cruzar esa membrana o capa protectora para llegar al interior. Igualmente, los animales, los vegetales y las células deben eliminar a través de esas cubiertas aquellos materiales que les son innecesarios o hasta dañinos. (Para una exposición más detallada sobre el tema de las membranas, puede consultarse Las membranas de las células, número 18 de esta misma colección.)
Pero hay otra circunstancia; muchas de las sustancias que existen dentro de
los seres vivos se encuentran en concentraciones mayores en su interior que
en el medio en que viven. Por ejemplo, el potasio (K+) está casi
siempre en todas las células a una concentración mucho mayor que las del medio
que las rodea (Figura IV.3), y no es éste el único caso. Es un hecho conocido
de todos que cualquier sustancia colocada en un líquido, por ejemplo, tiende
a distribuirse en éste por igual. Una gota de tinta colocada en un vaso con
agua termina con el paso del tiempo por distribuirse uniformemente en todo el
líquido. Esto se debe a que las moléculas del colorante tienen un movimiento
constante que se debe a una forma de energía, la energía cinética, que resulta
del simple hecho de encontrarse a una temperatura superior al cero absoluto;
ese constante movimiento es el responsable de la distribución uniforme, y su
origen es una forma de energía que las moléculas contienen. Como resultado de
ello, cualquier sustancia tiende a desplazarse de los sitios de mayor a los
de menor concentración.

Figura IV.3 Para que los iones de potasio (K+) entren a una célula
en la que se encuentran a mayor concentración que en el exterior, se necesita
energía que suele provenir, directa o indirectamente, del ATP. Si el potasio
sale a un medio de menor concentración, se disipa energía en forma de calor.
Es así que cada molécula o partícula de una solución tiene una cantidad de energía; la cantidad total de ésta depende de la cantidad de esa sustancia en un espacio dado. El caso es el mismo que para el aire comprimido: la energía contenida en un tanque depende de la cantidad que se haya logrado introducir, y la energía se manifiesta como la presión. Para comprimir el aire se requiere energía; la salida del aire, a su vez, se puede utilizar para realizar trabajo moviendo una compresora, una turbina, etcétera.
Esta comparación es válida para el movimiento de las sustancias a través de las membranas. Para introducir una sustancia a una célula o a un organismo, en contra de su tendencia natural de movimiento, se necesita energía. En ocasiones inclusive hay sustancias que se mueven a favor de esa tendencia natural, pero aun así utilizan energía, simplemente para asegurar que, una vez en un lado de la membrana, ya no van a regresar al otro.
Aunque todos los días comemos materiales sólidos e ingerimos líquidos (agua), pocos estamos conscientes de las complicaciones que supone este proceso. Dado que no es el tema que estamos tratando, sólo señalaremos aquí que en el aparato digestivo, estómago e intestinos, tiene lugar primero un proceso de digestión, en el cual las moléculas grandes deben ser separadas en sus componentes más pequeños (digeridas). Así, el almidón del pan, pastas, papas, etc., se transforma en glucosa; las proteínas de la carne, huevos, leche y otros alimentos se convierten en los aminoácidos que las componen; las grasas, a su vez, se rompen en su mayoría para dar también ciertos componentes, entre los cuales destacan los ácidos grasos. Hay además sales minerales cuyos componentes deben también cruzar la pared intestinal para ser aprovechadas por el organismo.
Las sustancias que resultan de la digestión no pueden cruzar libremente la pared intestinal; sólo una parte de los ácidos grasos y otras sustancias puede ser absorbida en forma directa. Ni siquiera puede pensarse en que todas son absorbidas por el intestino con la misma velocidad para pasar al interior del organismo. Tiene lugar un complicado mecanismo que requiere de un gran número de moléculas (proteínas) que están en las membranas de las células intestinales y son las que primero reconocen a las sustancias que deben ser absorbidas; es decir, no cualquier sustancia entra a las células por cada uno de estos sistemas, ni tampoco lo hace con la misma velocidad. Sólo una vez reconocidas y aceptadas se les permite el paso a través de la pared intestinal.
Volviendo al tema de la energía, es bueno preguntarse cuál es el componente
energético de este proceso. Hay que tomar en cuenta que muchas de esas sustancias
se absorben, y al absorberse deben transformarse en otras que no puedan regresar
por el mismo sistema que permitió su entrada. Éste es, por ejemplo, el caso
de la glucosa y otros azúcares: al entrar se les transforma, uniéndoles un fosfato
(Figura IV.4) que garantiza que el mismo sistema que los trajo al interior de
la célula ya no los reconozca, impidiendo así su salida. El fosfato que se les
une, por otra parte, proviene del ATP, y por tanto su unión representa el gasto
de energía. Pero el proceso no termina ahí, la célula de la pared del intestino
debe, luego, en el lado que ve hacia el interior del organismo, quitar de nuevo
el fosfato al azúcar y pasar éste hacia el interior propiamente dicho. De esta
forma, para tomar una molécula de glucosa del intestino (absorberla), se gasta
al menos un fosfato de alta energía del ATP.

Figura IV.4 Algunos aspectos sobre la energía y el transporte de los azúcares
y los aminoácidos en el intestino.
Hay otras sustancias que deben capturarse mediante procedimientos diferentes. Los aminoácidos, por ejemplo, no siempre están en el intestino en concentraciones altas; por tanto debe haber un mecanismo que asegure que se puedan utilizar, aun si sus cantidades son pequeñas. Para ello las células del intestino invierten energía, no directamente en la tarea de meterlos a la célula, sino en expulsar iones de sodio (Na+) al exterior. Dado que entonces habrá mayor concentración de éstos en el exterior, su tendencia natural es regresar al interior de la célula, y hay un sistema capaz de aprovecharla para introducir, y basta concentrar dentro de las células, grandes cantidades de aminoácidos (Figura IV.4). Veamos pues que, para este caso, si bien el gasto de energía es indirecto, en cierto modo tiene lugar en dos pasos.
También las plantas deben tomar, aunque en menor diversidad, sustancias que hay en el suelo para nutrirse. Para ellas el proceso no es mucho más simple; los vegetales requieren en cantidades grandes algunos minerales más o menos abundantes y amonio, pero también una gran cantidad de otros minerales que se encuentran en muy pequeña cantidad en el suelo, y que por tanto deben ser absorbidos mediante sistemas de altísima eficiencia y gran selectividad. Así como para las células intestinales hemos dicho que hay un sistema que se encarga de bombear el sodio al exterior, en los vegetales hay sistemas semejantes, aún no conocidos todos ellos, que también son capaces de tomar directamente del exterior algunas sustancias y concentrarlas en el interior, utilizando directamente la energía de los enlaces del ATP.
El transporte a través de las membranas celulares
Una vez que las sustancias del exterior son capturadas por los órganos especializados:
las raíces de las plantas o el intestino de los animales, y también por las
numerosas transformaciones internas de los organismos, resulta una enorme variedad
de sustancias, tanto dentro como fuera de las células. Todas están sujetas a
un tráfico de gran intensidad; deben ir y venir sin cesar de un lado a otro,
y muchas son producidas por unas células y utilizadas por otras, para lo cual
deben cruzar, de salida, la membrana de aquella que las produce, y de entrada,
la membrana de la que las recibe. Así pues, sucede que no sólo en las membranas
de las células intestinales o las de las raíces tiene que haber numerosos sistemas
de transporte; esto se requiere por fuerza también en todas las células del
interior. Nuevamente, en todos esos casos se debe invertir primero una gran
cantidad de energía en producir los sistemas de transporte, que generalmente
son proteínas membranales, y además, las células tienen sistemas que en su mismo
proceso de funcionamiento necesitan energía. Uno de los sistemas más universales
en los animales es la llamada bomba o ATPasa de sodio y potasio, que se encarga
de una función importantísima: mantener alta la concentración celular interna
de potasio y baja la de sodio, en contra de lo que sucede en el exterior, donde
la concentración de potasio es baja y alta la de sodio. El funcionamiento de
la ATPasa se ilustra en la figura IV.5. Para cada dos iones de potasio que entran
y tres de sodio que salen, se necesita gastar una molécula de ATP. Este sistema
funciona en todas las células animales y su funcionamiento implica un gasto
constante de energía. Las células vegetales también tienen sistemas para mantener
un contenido elevado de potasio en su interior, y sistemas semejantes para mantener
la composición ideal de iones del medio interno y del que las rodea.

Figura IV.5 La llamada "bomba" de sodio, que utliza la energía del ATP para
mantener una elevada concentración interna de potasio (K+) y baja concentración
de sodio (Na+) en las células de los animales.
Aunque hemos descrito la existencia de sistemas para mantener las concentraciones de potasio y sodio dentro y fuera de las células, hay muchos otros que conservan las concentraciones de diferentes iones, como el fosfato, el cloruro, el magnesio, el calcio, e inclusive algunos que deben existir en concentraciones extremadamente bajas pero que son necesarios para el funcionamiento celular.
Además del enorme tráfico de sustancias que hay en todas las células de un organismo, es importante considerar la cantidad de sales que ingerimos diariamente, por ejemplo, de cloruro de sodio que es la sal común, y que son absorbidas por el intestino y deben ser eliminadas por el riñón. Éste es otro ejemplo de una complicadísima combinación de estructuras que están diseñadas no sólo para filtrar constantemente nuestra sangre, sino además para mantener una composición invariable de nuestro líquido interno y finalmente de todo nuestro organismo.
Pero la complicación no termina ahí; así como los organismos están constituidos por órganos, tejidos y células que deben mantener un constante intercambio de sustancias, dentro de las células también hay una estructura y compartimentos separados por membranas. También aquí se deben establecer intercambios constantes, y las membranas deben contar con sistemas de transporte con características semejantes a las que hemos mencionado para otros sistemas. Las mitocondrias, el núcleo, los lisosomas, etc., deben mantener un activo intercambio de materiales, y ello implica que deben pasar por sistemas especiales a través de las membranas de los diferentes compartimentos.
También en el enorme mundo de los microorganismos, las bacterias, los protozoarios, las levaduras, etc., existen los mismos problemas. Una célula cerebral vive en un ambiente más o menos constante que le permite realizar una serie de actividades de gran especialización, pero que no requiere de una adaptación a condiciones diferentes ni variables de la solución que la baña. Sin embargo, una bacteria en el curso de su vida puede enfrentar condiciones de gran variabilidad. Lo mismo puede encontrarse en un medio nutritivo rico, que en otro en el cual apenas si habrá huellas de los materiales nutritivos y sales que necesita para vivir. Es por esta razón que los microbios necesitan de una gran variedad de sistemas de transporte para vivir. Por ejemplo, no es raro encontrar que una bacteria posea un sistema capaz de tomar sustancias del medio con una gran velocidad, pero con una afinidad relativamente baja, y que al encontrarse en un medio en el que ciertos materiales nutritivos disminuyen, desarrolle la capacidad de capturarlos con una gran eficiencia, aun si están en concentraciones sumamente bajas. Es claro, nuevamente, que para ello se requiere de energía.
Asimismo existen células en las cuales el transporte es tal vez el elemento esencial de su gasto de energía, pues su funconamiento está directamente relacionado con él. Tal es el caso de las células nerviosas, que transmiten los impulsos nerviosos mediante movimientos de iones de sodio y de potasio a través de sus membranas.
Esta forma de gasto de energía es tal vez la más simple de comprender; desde la escuela elemental se nos explica el papel de los alimentos como elementos necesarios para la realización del ejercicio físico. También es bien sabido que al hacer ejercicio físico se acelera la respiración; la ecuación que señala los materiales que intervienen en la degradación de los azúcares, como un ejemplo, puede explicar la relación entre el ejercicio físico, la respiración y el consumo o gasto de los alimentos:
Por otra parte, en el capítulo III se señalaron los mecanismos de control de la respiración y del metabolismo que permiten obtener energía en forma de ATP, y cómo éste se utiliza para realizar diferentes tipos de trabajo. Ahí se mostró de nuevo el mecanismo de control del metabolismo por el ADP, y la manera en que se relaciona el trabajo y el gasto propiamente dicho de energía.
El mecanismo de enlace entre el ATP y el trabajo mecánico es, por otra parte, sumamente simple. El ATP es utilizado directamente por las fibras musculares para contraerse; la energía del útimo enlace de fosfato es la que sirve en forma directa para este fin. Visto así, éste es tal vez el mecanismo más simple de conversión de una forma de energía, la química del ATP, en otra, la mecánica de la contracción muscular.
Ya hemos visto que hay un gasto muy grande de energía cuando se lleva a cabo una serie de funciones que pueden ser más o menos obvias dentro de los organismos. Sin embargo, es necesario considerar que para que éstas se lleven a cabo, se requieren muchísimas moléculas que las ejecuten. Así, hay enzimas para mover todo el metabolismo, hay proteínas que funcionan en las diferentes membranas celulares y que se encargan de mover a las diferentes sustancias de un lado al otro; obviamente hay moléculas que son las responsables del movimiento de las células o de los organismos; aun las células vegetales, que pertenecen a organismos que no se mueven, tienen una gran movilidad que se manifiesta durante muchas de sus funciones. El elemento común de estas moléculas ejecutoras de las diferentes funciones celulares es que son proteínas, es decir, moléculas muy complicadas formadas por la unión de 20 aminoácidos diferentes en número y orden, que son perfectamente constantes y definidos, pero característicos para cada una de ellas y diferentes de las otras proteínas, que así dan la posibilidad de realizar tantas y tan variadas funciones con precisión verdaderamente impresionante en todos los casos.
Todos los organismos deben tener, de algún modo, no sólo la información necesaria para sintetizar todas estas moléculas, sino también la capacidad de transmitirla prácticamente sin equivocación de unas células a otras; y lo que más llama la atención es que esta transmisión permite conservar las características de todas y cada una de las células y da lugar a la generación ininterrumpida de individuos esencialmente iguales a través del tiempo.
Esta información, por otra parte, y especialmente desde el punto de vista de nuestro tema de análisis de los seres vivos, requiere de energía para mantenerse y para interpretarse. La base de esto es la existencia de moléculas que la contienen, y que deben poderse duplicar durante la duplicación de una célula; pero este complicado sistema de síntesis implica un gasto de energía. Luego, para que esa información se pueda convertir en realidad, hay también un complejo sistema de ejecución o interpretación, que requiere igualmente de un gasto grande de energía.
Todos sabemos que la molécula en la que se mantiene la información para la síntesis de todos los componentes de una célula es el DNA (abreviatura del nombre en inglés del ácido desoxirribonucleico), al cual algunos llaman también ADN. De manera fundamental, podemos considerar el gasto de energía que se requiere para duplicar y mantener en cada célula en forma constante, la información genética y transmitirla; luego, hay otro gasto necesario para que esta molécula exprese esa información de manera útil para las células, es decir, mediante la síntesis de moléculas de proteína.
La síntesis del DNA implica, primero, la síntesis de las moléculas llamadas
nucleótidos, a partir de las cuales se va a formar. La figura IV.6 muestra esquemáticamente
los nucleótidos (que en este caso se llaman desoxinucleótidos, porque el azúcar
que en ellos interviene es la desoxirribosa y no la ribosa); éstos se parecen
al ATP, pero pueden tener, en lugar de la adenina, otras bases orgánicas: la
guanina, la citosina y la timina. Sin embargo, todos se llegan a sintetizar
como los trifosfatos, semejantes al ATP, porque para sintetizar al DNA la enzima
encargada de ello, utilizando como molde a otra molécula ya existente de DNA,
va formando la nueva a partir de los trifosfatos, pero eliminando al formar
cada enlace a dos de los fosfatos, los cuales son de "alta energía", igual que
en el caso del ATP. Esto da idea de la cantidad de energía que se requiere para
duplicar la molécula de DNA, que en primer lugar tiene dos hilos, y en segundo,
es muy larga, ya que cada aminoácido se codifica con tres nucleótidos.
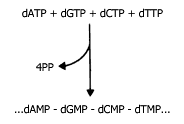
Figura IV.6 Síntesis de una cadena de nucleótidos en el DNA: Cada nucleótido
que se une implica la ruptura de dos fosfatos.
La historia, además, no termina ahí; el DNA no existe en las células en forma libre; se debe asociar a otros componentes, de los cuales los más abundantes son proteínas llamadas histonas, que puede decirse que lo "visten" y le permiten convertirse en una estructura que también nos es conocida: los cromosomas de las células (las bacterias tienen su DNA desnudo de estas proteínas). Así pues, el solo hecho de mantener la información en una célula implica un gasto muy alto de energía. Es necesario mencionar inclusive que el DNA está sujeto a numerosos accidentes en su vida; la naturaleza no se permite el lujo de que su información se deteriore, y cuenta con mecanismos para reparar sus moléculas cuando llegan a sufrir daños.
La información contenida en el DNA debe luego contar con un sistema de utilización,
para que a partir de ella se sinteticen las proteínas, que serán las verdaderas
ejecutoras de toda la información que hemos mencionado. Para ello, a partir
del DNA de cada célula, pero sólo de una parte de él, se producen "mensajes"
mediante un proceso que por analogía con un mecanismo de información se llama
"transcripción", y en el cual se produce el ácido ribonucleico (RNA), que algunos
abrevian como ARN. En especial, el RNA que lleva estos mensajes, leídos o transcritos
del DNA, recibe el nombre de RNA mensajero, y se le abrevia RNAm (Figura
IV.7).

Figura IV.7 Esquema de la transcripción de un mensaje del DNA al RNA. Cada
nucleótido que se agrega implica el "gasto" de dos fosfatos de alta energía.
Este mismo mecanismo opera en la síntesis del RNA mensajero (RNAm),
el ribosomal (RNAr) y el de transferencia (RNAt).
La síntesis de las proteínas no se realiza simplemente por la formación del RNAm; en el proceso intervienen también los ribosomas, que a su vez están formados en gran parte por otro tipo de RNA, el llamado ribosomal (RNAr). También interviene otro tipo de RNA, el llamado de transferencia (RNAt). Lo interesante en el proceso es que todos los tipos de RNA se sintetizan "leyendo" del DNA la información que se encuentra en él contenida, en un proceso en el cual los nucleótidos trifosforilados ATP, GTP, CTP y UTP se unen unos con otros mediante reglas sencillas, utilizando como "molde" al DNA, y eliminando o "gastando" en la unión de cada uno, dos de los fosfatos, como se hizo en el caso del DNA. Así, la lectura de esa información para obtener las moléculas de RNA necesarias para la síntesis de las proteínas cuesta a las células una elevada cantidad de energía.
Pero además de todo esto, hay que considerar que los ribosomas, organelos en los cuales se realiza la síntesis misma, están compuestos no sólo por el RNAr, sino por toda una serie de enzimas que catalizan las distintas reacciones, y también hay enzimas adicionales aparte de los ribosomas, que intervienen en el proceso. Visto así el problema, se puede tener una idea vaga pero más o menos aproximada de la magnitud de gasto de energía que la conservación y el procesamiento de la información contenida en las células puede costarles.
Ya se mencionó cómo se puede medir la cantidad de energía que se "libera" en forma de calor al quemar una cierta cantidad de glucosa; la verdad es que lo que se mide es la energía que se transforma en calor durante el proceso de oxidación del azúcar.
Cualquier organismo vivo, por el solo hecho de existir, tiene que realizar numerosas transformaciones de energía para poder mantener todas las funciones que de alguna forma se han mencionado aunque sea parcialmente en este capítulo. Todos los días ingerimos alimentos que se degradan, y la energía de sus enlaces se transforma en la de los del ATP, y ésta a su vez es la que, en lo fundamental, es también aprovechada por muy diferentes sistemas para la realización de las funciones vitales de los organismos. De suerte que la energía del ATP puede ser aprovechada para el movimiento, la síntesis de ciertas moléculas, el movimiento de otras a través de las membranas, etcétera.
Pero tal vez lo más importante de todo este proceso es que se trata de un ciclo, es decir, desde el punto de vista mecánico iniciamos nuestras actividades diarias en la mañana, y las terminamos en la noche, o a la mañana siguiente esencialmente en el mismo sitio que el día anterior. Esto quiere decir que no llegamos a acumular ninguna forma de energía mecánica en nuestra vida diaria, y cada día, la que desarrollamos termina por convertirse también en calor. De la misma forma, cada día que transcurre, salvo en el caso de los niños, sobre todo pequeños, el número y cantidad de moléculas de nuestro organismo se mantiene constante. Se "gasta" energía para sintetizar proteínas, moléculas grandes de azúcares y grasas, pero cuando éstas se degradan la misma cantidad de energía se transforma en calor.
También cada día, no obstante que todas nuestras células realizan una gran actividad de transporte de moléculas de diferentes tipos, el balance diario es cero. Otra vez, la energía que se gasta o utiliza para mover las moléculas, cuando éstas regresan a su estado anterior es liberada o transformada en calor.
En conjunto, pues, cada día la energía de las moléculas con que nos alimentamos se utiliza para la realización de todas nuestras funciones, pero al final, es transformada en calor. Así, el resultado diario es que, en términos de energía, toda aquélla contenida en los alimentos es transformada en calor. Por otra parte, la cantidad de éste es un excelente indicador de la energía que hemos utilizado para realizar nuestras funciones. A fin de cuentas, tal como lo predijo Lavoisier en el siglo XVIII, la energía calorífica que produce un ser vivo alimentado con glucosa, por ejemplo, es la misma que la que se libera por la combustión de esa sustancia en un calorímetro. De aquí resulta que es también posible introducir a un animal o a un ser humano en un espacio cerrado, semejante en todo a un calorímetro, y medir el calor que desprende como resultado de las transformaciones de energía que ocurren en su interior.
Si además es posible saber la cantidad de calor que se desprende de los azúcares, las grasas y las proteínas, es también posible, conociendo el desprendimiento de calor de una persona, saber la cantidad de alimentos que necesita para satisfacer los requerimientos de energía. Se sabe que un individuo adulto que realiza una actividad promedio diaria necesita alrededor de 30 kilocalorías por cada kilogramo de peso corporal. Un individuo de 70 kilogramos, por ejemplo, requiere alrededor de 2 100 kilocalorías diarias distribuidas entre sus diferentes alimentos.