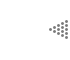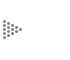III. LA ENERGÍA DEL MUNDO ANIMAL: EL APROVECHAMIENTO DE LOS ALIMENTOS
COMO ya se mencionó en los capítulos anteriores, las células están
compuestas de moléculas, a su vez constituidas en su mayor parte por seis elementos
principales, que son: carbono, hidrógeno, nitrógeno, oxígeno, fósforo y azufre;
estos elementos forman 99% de su peso. Por otra parte, el agua es la substancia
más abuntante en la célula y ocupa 70% de su peso. El átomo de carbono desempeña
un papel importantísimo en la biología, debido a que es capaz de formar moléculas
de gran tamaño y variedad, ya que puede formar cadenas o anillos (Figura III.1).
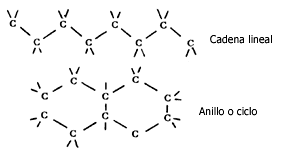
Figura III.1 Cadenas de carbonos.
Los átomos de carbono forman enlaces muy fuertes y resistentes ya sea entre ellos mismos o con otros átomos, los cuales se conocen como enlaces covalentes. Cada átomo de carbono se puede combinar con otros, y formar así un número muy grande y variado de compuestos.
Pero los enlaces, por su propia "fuerza" o energía, representan en realidad la forma en la que nuestras células reciben energía y la pueden utilizar, mediante complicados procesos, que trataremos de analizar en este capítulo.
Antes de empezar, señalaremos el significado de algunos términos que se utilizan con frecuencia al hablar de las transformaciones de las sustancias que se encuentran en los seres vivos. En primer lugar, las células cuentan con caminos para formar moléculas más pequeñas a partir de moléculas grandes, y a este proceso se le llama catabolismo. Hay un proceso inverso, que consiste en la formación de moléculas más grandes, a partir de otras más pequeñas, que recibe el nombre de anabolismo. De forma general, a todo el conjunto de transformaciones que sufren las sustancias en el organismo o en una célula se le llama metabolismo.
LA DEGRADACIÓN DE LAS MOLÉCULAS
Para que las células puedan aprovechar las sustancias en sus distintas funciones
deben primero degradarlas. Los procesos de degradación, o catabólicos, ocurren
en tres etapas; en la primera, se rompen las grandes moléculas en sus componentes
más sencillos, las proteínas en aminoácidos, los carbohidratos en azúcares sencillos
y las grasas en ácidos grasos (Figura III.2). Esta degradación de las moléculas
grandes libera energía que se disipa en parte en forma de calor. En una segunda
etapa, estas pequeñas moléculas son a su vez degradadas para formar moléculas
todavía más pequeñas, con la posibilidad de obtener energía útil para la célula.
Estas moléculas pequeñas son el piruvato y la acetil coenzima A; el piruvato
también a su vez se transforma en acetil coenzima A.
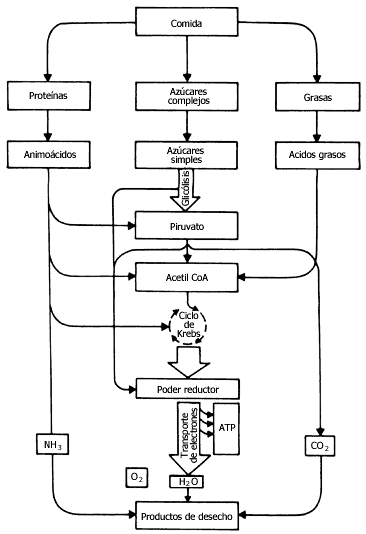
Figura III.2
Para el caso de los azúcares, por ejemplo, en la primera etapa se degradan los polímeros, como el glucógeno, para dar glucosa. En la segunda etapa, la glucosa se degrada para dar piruvato, y éste se convierte en acetil coenzima A. Finalmente, ésta se degrada para dar CO2 y H2O. Es necesario señalar que, de las tres etapas, sólo en las dos últimas se obtiene energía aprovechable por la célula, en forma de ATP. La degradación de la glucosa a piruvato u otros compuestos cercanos es probablemente el camino metabólico más antiguo que existe, y todavía algunos organismos lo utilizan para obtener ATP.
El esquema de la figura III.3 se presenta para dar sólo una idea de lo complicada
que puede ser una vía metabólica. Con objeto de obtener energía y otras sustancias,
tan sólo para partir a la molécula de glucosa en dos fragmentos iguales de piruvato
o lactato, se requiere de un gran número de pasos, catalizados cada uno por
una enzima diferente. La degradación de la glucosa, o glucólisis, se puede llevar
a cabo tanto en ausencia como en presencia de oxígeno. Sin embargo, lo más importante
del proceso es que parte de la energía contenida en los enlaces de la glucosa
puede transformarse, con bajo rendimiento, en la de los enlaces del ATP, directamente
aprovechable por la célula. Pero aunque una molécula de glucosa que se degrada
para dar ácido láctico sólo produce dos moléculas de ATP, esta vía puede funcionar
a gran velocidad en algunas células, las musculares, por ejemplo. Los atletas
que participan en las pruebas rápidas, como por ejemplo la carrera de los 100
metros, obtienen casi toda la energía para la competencia, de esta vía metabólica.
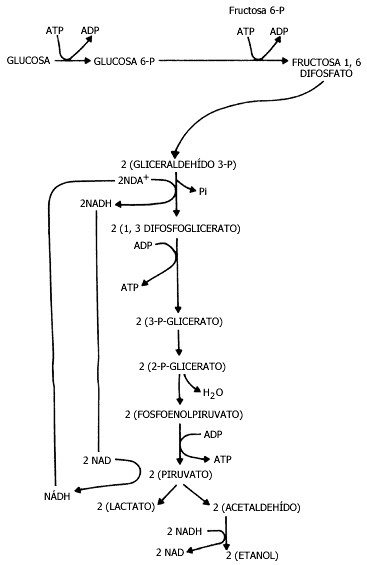
Figura III.3 Glucólisis.
La fermentación es una variante de este proceso de degradación de la glucosa hasta convertirla en CO2 y alcohol; la levadura, durante esta etapa, obtiene toda su energía a partir de la degradación de la glucosa. Es afortunado, en cierta forma, que la vía sea poco eficiente, pues para obtener la energía, estos hongos (las levaduras) deben transformar en alcohol y en CO2 grandes cantidades de glucosa. Por ello, la levadura puede utilizarse en la fabricación de pan, con el objeto de que produzca pequeñas burbujas internas de CO2, que al calentarse en el horno se dilatan y lo vuelven esponjoso. También la levadura puede producir grandes cantidades de alcohol, que pueden ser de gran utilidad en la industria y nos ofrecen, entre otras cosas, la cerveza y el vino.
Por cada molécula de glucosa se obtienen al final del proceso dos moléculas de lactato cuando se recorre el camino completo. y en el caso de la fermentación se producen dos moléculas de etanol (alcohol). Esta vía metabólica, la glucólisis, tiene una gran importancia pues además de proporcionar ATP a la célula, proporciona el piruvato que luego se ha de transformar en acetil coenzima A, que le permite continuar, utilizando otra vía metabólica, con la degradación hasta bióxido de carbono y agua, como veremos a continuación.
Las proteínas que se ingieren en la dieta no se aprovechan como tales, es decir, existen mecanismos de degradación que se llevan a cabo en el tubo digestivo. Mediante procesos más o menos complicados, se digieren para dar sus componentes, los aminoácidos, que se absorben por las paredes del intestino y son aprovechadas por nuestros tejidos. Las enzimas digestivas rompen las moléculas de proteína en fragmentos cada vez más pequeños, hasta degradarlas en sus constituyentes básicos, los aminoácidos, que de esta forma sí pueden ser absorbidos por la pared intestinal. Los aminoácidos se procesan dentro de la célula mediante distintas enzimas, que también pueden convertirlos en acetil coenzima A. No entraremos en los detalles de la transformación química de los aminoácidos; baste saber que sus esqueletos de carbono son utilizados como combustible para alimentar una vía metabólica de extraordinaria importancia, que es comparable con un molino, y cuya descripción completa se debe al trabajo de muchos científicos, pero fue integrada en 1935 por uno de ellos, Hans Krebs, en cuyo honor se le suele dar el nombre de ciclo de Krebs, o de los ácidos tricarboxílicos.
EL CICLO DE KREBS, PARA QUÉ SIRVE
Este ciclo de Krebs o ciclo de los ácidos tricarboxílicos es fundamental para
el metabolismo energético de la célula, ya que provee o alimenta de hidrógenos
a la cadena respiratoria, y sirve de base para la producción de la mayor parte
de la energía en los organismos aeróbicos. Se trata de un mecanismo complicado
que llevó varios decenios descubrir y entender. En forma muy resumida, puede
decirse que se alimenta de acetil coenzima A que proviene, como ya vimos, de
los carbohidratos, las grasas y las proteínas. Se dice que es un ciclo porque
termina en el mismo compuesto con que se inicia. La serie de transformaciones
que se muestran en la figura III.4, señala dos cosas principales, la primera
es que el ciclo puede alimentarse de moléculas de dos átomos de carbono (acetatos),
que le ofrecen la acetil coenzima A, y durante una vuelta, estos dos átomos
de carbono salen en forma de CO2. Pero la finalidad más importante
de este ciclo consiste en proporcionar un gran número de hidrógenos, que entran
en la cadena respiratoria mitocondrial para ser oxidados (combinarse con el
oxígeno), y así dar finalmente moléculas de agua y obtener ATP en el proceso.
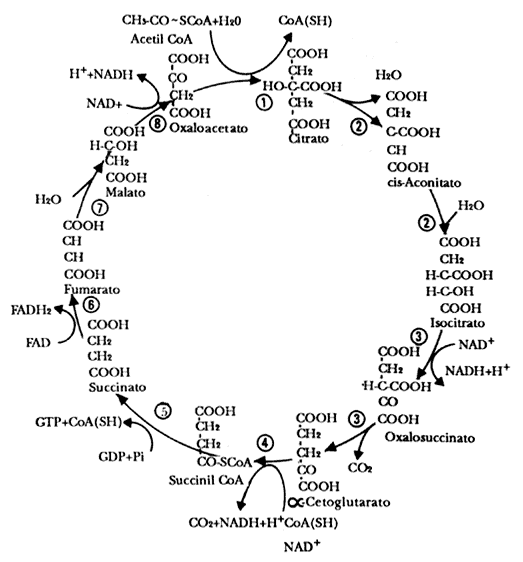
Figura III.4 Ciclo de Krebs. Las 8 enzimas que participan en el ciclo son:
1) citrato sintasa; 2) aconitasa; 3) isocitrato deshidrogenasa; 4) a
cetoglutarato deshidrogenasa; 5) succinato tio-cinasa 6) succinato-coenzima
Q reductasa; 7) fumarasa y 8 ) malato deshidrogenasa.
El fragmento de dos átomos de carbono, el acetato de la acetil CoA, entra al
ciclo de los ácidos tricarboxílicos o ciclo de Krebs, el cual produce los agentes
reductores que a su vez alimentan a la cadena respiratoria, la cual genera la
fuerza que se requiere para la síntesis de ATP (véase la figura III.5).
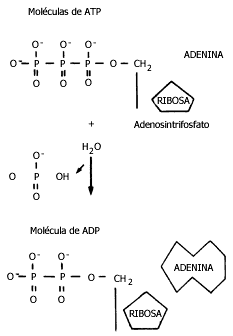
Figura III.5 Moléculas de adenosintrifosfato y adenosindifosfato.
Como ya se mencionó en el capítulo I, la molécula de ATP (Figura III.6) contiene
tres grupos fosfato y libera energía cuando se desprende el último de éstos
al ser "hidrolizado", al romperse con una molécula de agua. La cantidad de energía
que se libera puede en muchos casos servir para que otra reacción química ocurra.
Utilizando una analogía de la naturaleza, es como si el agua que corre por un
río, que siempre va cuesta abajo, corriera un día cuesta arriba; esto que parece
imposible, es lo que la célula tiene que hacer todo el tiempo para sobrevivir
y dividirse, ya que en un organismo vivo existe una constante tendencia al desorden
o al equilibrio con el medio que la rodea. Para evitar caer en este equilibrio
o desorden de manera total, todo organismo vivo debe gastar energía química
a partir de la cual se sintetizan componentes celulares o bien se llevan a cabo
procesos, como el transporte de nutrientes o el movimiento, que requieren de
ella.
Figura III.6 El flujo de energía en los seres vivos.
LA FOSFORILACIÓN OXIDATIVA: EL ATP Y EL PODER REDUCTOR
Como hemos visto, el metabolismo tiene dos componentes, uno de degradación y otro de síntesis; en pocas palabras, la fase degradativa produce ATP y la de síntesis lo utiliza. El ATP es probablemente la molécula más utilizada del organismo; esto ha hecho que un gran número de grupos de investigación en el mundo se hayan interesado en estudiar los mecanismos de síntesis de este compuesto. Veamos en qué consiste este mecanismo conocido como fosforilación oxidativa, cuyo nombre proviene del hecho de que una molécula de ADP adquiere un fosfato más (se fosforila), simultáneamente con una serie de transferencia de electrones u oxidaciones de distintas moléculas. Para entenderlo debemos primero revisar las reglas de este juego que diseñó la naturaleza.
Uno de los principios en los que se basa este fenómeno es que las células poseen
membranas que actúan como barreras impermeables que las aislan del medio que
las rodea; por otra parte, las células poseen en su interior organelos que a
su vez están contenidos por membranas que los aislan del medio que los rodea.
De esta forma los ambientes dentro de cada estructura están regulados y pueden
llegar a ser completamente diferentes. Esta es la clave del proceso de fosforilación
oxidativa que ya mencionamos, o sea la síntesis de ATP; el proceso ocurre dentro
de un organelo que se conoce como mitocondria en los seres superiores, y como
cloroplasto en las plantas superiores. Ambos tipos de organelos son altamente
especializados, y poseen dos clases de membranas, una externa y una interna;
la externa es altamente permeable y permite el paso de muchas moléculas que
se difunden libremente. La interna es impermeable y es en donde se encuentra
la maquinaria para sintetizar el ATP (Figura III.7).
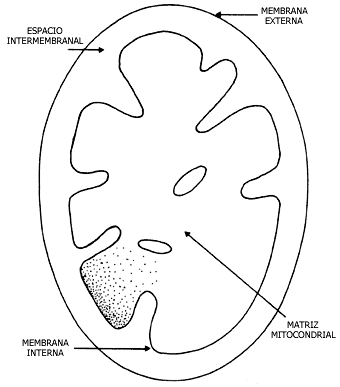
Figura III.7 Esquema de una mitocondria.
La maquinaria que se encarga de sintetizar la molécula de ATP está incluida o sumergida dentro de la membrana interna de la mitocondria y está constituida por proteínas especializadas en las funciones que a continuación describimos. Después de muchos años de investigación, se ha llegado a entender que existen proteínas que, a diferencia de la gran mayoría de las proteínas solubles, pueden llevar a cabo procesos de transporte de especies químicas que no pasarían a través de una membrana de no ser por ellas.
En la mitocondria estas proteínas aceptan y donan electrones, los cuales provienen originalmente de los hidrógenos que proporciona el ciclo de Krebs. Pero lo más importante es que, como ya mencionamos para el cloroplasto, tienen acoplados a su vez procesos de transporte. En otras palabras, cuando una molécula dona su electrón a una de las proteínas de la membrana mitocondrial, el electrón es transportado hacia el oxígeno, pero no en forma directa, sino a través de varios aceptores. En algunos de los pasos, de manera simultánea al transporte de los electrones hacia el oxígeno y aprovechando la energía con que esto sucede, se "bombean" protones, o hidrogeniones (H+) hacia el exterior de la mitocondria.
La esencia del proceso es que las proteínas de la membrana mitocondrial, que se llaman también transportadoras de electrones, se encuentran formando una cadena que termina en el oxígeno, y que al funcionar bombea protones al exterior. Estos protones tienen una gran tendencia a regresar al interior, y representan una forma de energía. Así se genera una fuerza capaz de proveer la energía que requiere el proceso de síntesis de ATP.
Como se mencionó en el capítulo anterior para el caso de la fotosíntesis y el cloroplasto, los protones tienden a regresar por la propia energía que su diferencia de concentración a ambos lados de la membrana les proporciona. En el caso de las bacterias, sucede lo mismo, pero es la membrana externa la que hace las veces de la membrana mitocondrial. La fosforilación oxidativa se lleva a cabo en la membrana interna mitocondrial o en la membrana plasmática de las bacterias; utiliza como sustrato para el proceso al adenosín difosfato, al cual se añade un grupo de fosfato en el extremo de la molécula, gracias a que existe una proteína membranal que se encarga de ello. La energía que proporciona la diferencia de concentración de protones se aprovecha gracias a una enzima que se llama ATP sintetasa o ATP sintasa, para unir al ADP con el fosfato y dar el ATP. Esta proteína está muy ampliamente distribuida en los seres vivos, desde los organismos más primitivos, como las arquebacterias, hasta las células de los organismos superiores, y en todas tiene esta función primordial de sintetizar el ATP.
EL CONTROL DE LA UTILIZACIÓN DE LOS ALIMENTOS
En el caso de la glucólisis es muy claro; si se revisa el esquema, en dos de las reacciones el ADP es un componente de ellas. Puede notarse que si no hay ADP, no es posible que la vía completa funcione. Aunque, desde luego, en condiciones naturales no existe el estado en el cual el ADP se agote, es un hecho que éste se produce con mayor o menor velocidad, dependiendo del trabajo que realicemos, pues durante el trabajo intenso se gasta mucho ATP, que se transforma en ADP y fosfato. Resulta así que la glucólisis en especial es indirectamente sensible al trabajo que hacemos, y si no trabajamos, no responde, o lo hace sólo para mantener nuestras funciones vitales, como el movimiento de los pulmones, nuestro corazón, etcétera.
En el caso de la fosforilación oxidativa, aunque de forma no tan clara, también
sucede que, incluso las mitocondrias aisladas, y aun presente cualquiera de
los intermediarios del ciclo de Krebs que les proporcione los hidrógenos para
alimentar la cadena respiratoria, no utilizan el oxígeno, a menos que tengan
ADP y fosfato. Así resulta también que la respiración y la fosforilación oxidativa
están controladas como se muestra en la figura III.8. Cuando realizamos trabajo
se gasta ATP, que se convierte en ADP y fosfato, y en especial el primero, o
sea el ADP, estimula la respiración. Aunque el proceso es más complicado y en
él intervienen muchos otros mecanismos, podemos fácilmente percibir que si corremos
un poco, o hacemos movimientos bruscos y constantes, o cualquier otro tipo de
ejercicio, se acelera nuestra respiración.
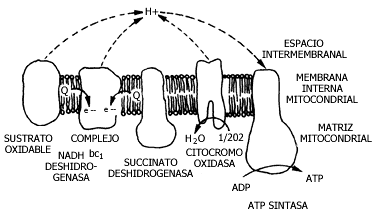
Figura III.8 Esquema de la cadena transportadora de electrones y de la fosforilación
oxidativa. En la membrana interna mitocondrial se encuentra dispuesto el sistema
que transforma la energía metabólica en química (ATP).
El otro elemento importante es que el trabajo celular no sólo acelera la respiración o la glucólisis; también debe aumentar el consumo de las sustancias o intermediarios metabólicos que provienen originalmente de nuestros alimentos. Por el mismo mecanismo, el consumo de alimentos está regulado por la cantidad de trabajo. En otro de los capítulos de este libro hablaremos de esta relación que existe entre el consumo de alimentos y el trabajo que realizamos.