VI. CON LAS CUENTAS CLARAS SE CONSERVAN LOS AMIGOS, O RECORDAR A MICHEL Y PERDERSE ENTRE NÚMEROS
El propósito de la taxonomía es agrupar en clases "naturales" los objetos a clasificar. Esta noción de "natural" ha sido definida de distintas formas, pero detrás de todas ellas está la idea común de que un miembro cualquiera de una de estas clases naturales está más próximo a los de su misma clase que a cualquier otro. Esto nos lleva a intentar definir lo que queremos indicar por "relación taxonómica". Los taxonomistas tradicionales pretenden identificar las relaciones taxonómicas con las relaciones evolutivas, pero, por otra parte, los taxonomistas numéricos han señalado que las relaciones taxonómicas son de tres clases. Reciben el nombre de "fenéticas" aquellas relaciones basadas en una semejanza general entre los objetos a clasificar, y "cladísticas" aquellas otras basadas en líneas comunes de descendencia ... La tercera clase de relación taxonómica es la "cronística" o temporal, relación de varias ramas de la evolución... Las clasificaciones "filogenéticas" de la taxonomía convencional se basan, en general, en una mezcla indefinida de relaciones fenéticas y cladísticas, y en muchos casos sólo representan una semejanza general entre los organismos clasificados, disfrazadas con una terminología evolucionista. Taxonomía numérica (1968) ROBERT SOKAL
LA AUSENCIA de un método generó numerosas disputas conceptuales, y los debates sobre los aspectos más controversiales de la taxonomía no hallaban una solución satisfactoria. Los procedimientos en taxonomía biológica eran severamente criticados por otros científicos, pues afirmaciones de "autoridad científica" como aquélla de que una especie es el conjunto de poblaciones que un taxónomo experimentado dice reconocer, sin señalar más requisito que la mera opinión del especialista, hacían que el conjunto de los científicos advirtiera que la clasificación biológica no distaba mucho de lo arbitrario y subjetivo.
Algunos de los representantes más destacados en el estudio de la evolución y de la taxonomía opinaron que ésta era una mezcla entre arte y ciencia; lo cual logró encender los ánimos aún más, y despertar en muchas mentes la inquietud de la búsqueda de un método natural que basara sus procedimientos clasificatorios en argumentaciones más rigurosas y formales.
Las polémicas son el común denominador en la ciencia; siempre existen diferencias en el estudio de los problemas. No hay ni se llega a menudo a opiniones monolíticas o exclusivas; durante el debate de las distintas opiniones se alcanzan algunos acuerdos, se aclaran los conceptos, se proponen nuevas teorías o explicaciones que permiten un mejor entendimiento de las cosas, de los fenómenos, de los eventos y de los procesos. Las ideas también evolucionan; nunca se quedan quietas.
Sokal, de quien hablaremos más adelante, señaló: "En algunos de los recientes congresos sobre biología, las sesiones correspondientes a la taxonomía numérica se han visto muy concurridas, en muchas ocasiones por personas remotamente interesadas en el asunto, que habían oído decir que 'iba a haber una buena pelea'. ¿A qué viene todo este ruido?"
Con frecuencia se tiene la idea de que la ciencia produce, verdades eternas e incuestionables; nada más lejos de la verdad. La ciencia se aprecia, desde un ángulo mítico, como infalible. Sin embargo, esto no significa que no tiene pretensiones acerca de encontrar un modo racional de explicar la naturaleza. Para ello parte de suposiciones tales como: el mundo es ordenado, hay patrones de orden y los patrones requieren explicaciones causales. En este sentido, la clasificación es una de las tareas fundamentales de la ciencia.
La ciencia es ambiciosa, y en el sendero de la historia que recorre, en agrias o amigables discusiones de sus practicantes, consigue paulatinamente una explicación de los hechos o, cuando menos, se alcanza algún progreso en el planteamiento de lo que se reconoce como problemas.
En aras de restar arbitrariedad o subjetividad a los procedimientos de la clasificación biológica, emergieron nuevas teorías taxonómicas. Fueron propuestas nociones inéditas por varias de las corrientes de pensamiento que se estaban gestando; una de las tendencias que apareció fue la llamada taxonomía numérica, fenética o neoadansoniana. Este último término no está bien aplicado, pues si bien los feneticistas tenían la idea de clasificar los organismos mediante el examen de todos los caracteres, sin darles valores distintos, ellos no pretendían encontrar las esencias de las especies, de los géneros y de las familias, como lo intentó Adanson. Había diferencias de intención, aunque la evaluación de la semejanza general constituyera la constante.
Hoy nadie rechaza el valor importante que los tratamientos numéricos poseen para hacer más rigurosos y formales los procedimientos clasificatorios. En lo que se difiere es en la aplicación de tales tratamientos según sean los procesos, objetos y fenómenos que se quieren clasificar. No es lo mismo clasificar a estrellas, átomos y moléculas que a seres vivos. Cada conjunto tiene propiedades distintas que deben tomarse en cuenta; no obstante, Roberto Sokal tiene razón cuando dice: "La naturaleza de la semejanza es, desde luego, uno de los problemas fundamentales de la taxonomía, cualquiera que sea el enfoque teórico de partida."
ENTRE ÁRBITROS Y CUENTACHILES:
EL ¿POR QUÉ? ETERNO DE LOS NIÑOS
La imaginación y la intuición no son ajenas a la ciencia. Aunque pretendidamente distanciadas del rigor y del sentido "estricto" del científico, son base importante en los descubrimientos y en las invenciones. Esto no quiere decir que sean el último "misterio" del intelecto humano, pero cuando menos les atribuimos el "genio" del científico. Se reconoce cada vez más que la capacidad de imaginar o de intuir descansa en el conocimiento de condiciones análogas sobre las que pensamos; inferimos comportamientos o relaciones equivalentes a partir de aspectos que conocemos. Señala un dicho que la imaginación es el sentido común glorificado.
Al tomar conciencia de esta situación, los taxónomos numéricos exigieron a la sistemática tradicional que hiciera explícitos sus razonamientos en las decisiones y procedimientos que se efectuaban. Era el destino inexorable de la ciencia según ellos: formalizar, cuantificar y medir: la periferia del sentido común.
Una de las dificultades por la que la taxonomía se privaba de su carácter científico y estaba imposibilitada de encontrar la tan cacareada clasificación natural estribaba en sus prácticas imprecisas que no permitían la cuantificación. No es que los feneticistas tuvieran la obsesión pitagórica de los números, el interés de encontrar números mágicos en la naturaleza, como Linneo con su sistema sexual que expresaba el orden en números. Se deseaba reducir las decisiones taxonómicas vagas, difusas e intuitivas que hacían de la taxonomía un "arte".
Por otra parte, la teoría de la evolución de Darwin no explicaba muchos aspectos, sino que se extrapolaban conclusiones que muchos no estaban dispuestos a aceptar. Tales extrapolaciones eran vistas como extraordinariamente subjetivas, sin ninguna base de observación, sin evidencia. Darwin había puesto la semilla en la búsqueda de explicaciones racionales en el estudio de la evolución, pero no todo era adaptación y selección natural.
Aquellos que creían que la clasificación no podía sustentarse en teorías, debido a que consideraban que era aceptar especulaciones que incluso rayaban en la fantasía, se opusieron en principio a usar numerosos conceptos, principalmente evolutivos. Pensaban que la clasificación de los seres vivos debía enfrentarse en una posición de franco empirismo.
Los antagonismos entre las dos corrientes de pensamiento más conspicuas en la clasificación incidían en concepciones muy distintas de lo que debía ser una clasificación natural y en las formas de encontrarla o descubrirla.
De lo "natural" en el descubrimiento de las esencias, a través del estudio de todos los caracteres para la construcción de las clasificaciones —como lo intentó Michel Adanson—, se había pasado al estudio de la semejanza total como una opción posible en la clasificación en tiempos darwinistas, pero que nunca se desarrolló debido a que muy pronto comenzó a erigirse la idea rectora de la genealogía como fundamento de la clasificación biológica. Dado que ésta no había desarrollado un método y conceptos teóricos que permitieran reconstrucciones filogenéticas rigurosas, con estricta lógica, alejadas de la "intuición" y de posiciones de seudoautoridad científica, la postura numericista planteaba una fórmula que equivalía al regreso del concepto de naturalidad por semejanza total, que hacía un siglo parecía haberse superado.
Ni Adanson ni los coetáneos de Darwin pudieron hacer prosperar la idea de manejar muchos caracteres, debido a un pobre desarrollo de las matemáticas de aquel tiempo y a la ausencia de computadoras, entre otros aspectos. Pero cuando ciencia y tecnología se encontraron en condiciones de ejecutarla, la idea de semejanza total se veía como una alternativa potencial para enfrentar el estudio de la diversidad organísmica, bajo una perspectiva empírica, "desprovista" de la teoría so pretexto de erradicar la especulación y las dificultades que no se habían podido superar en historia evolutiva, por ejemplo: un concepto de especie suficientemente general para todos los organismos y consecuente con el proceso evolutivo, así como un método y conceptos que admitieran la reconstrucción genealógica ausente de decisiones evidentemente subjetivas o arbitrarias.
Se pensaba que con el uso de la estadística y las probabilidades se podrían erradicar decisiones tendientes a la subjetividad; la estadística recién había mostrado su gran potencial e importancia en el estudio de la genética, de la selección natural y de la variación. Dicha posición tenía posibilidad de ganar adeptos.
La teoría sintética de la evolución y la nueva sistemática garantizaban muchos ingredientes y su calidad, pero en el momento de hacer el pastel no existía receta para explicar el orden y la relación de los ingredientes en los términos requeridos. No había un método con sustento teórico necesario y suficiente. Por tales motivos, la idealización estadística pareció atractiva a muchas mentes. Resultaba oportuna para sustituir temporalmente el principio de descendencia con modificación por el de semejanza total en la clasificación biológica.
Se había convertido en un mito la creencia de que la filogenia o la genealogía podían inferirse a partir de la clasificación y viceversa. Se alejaba la posibilidad de considerar que la proximidad de los nombres en una clasificación era la indicación de proximidad de descendencia, debido a que la proximidad de nombres en una clasificación significaría para los numericistas proximidad de semejanza total. Vistas así, eran dos posturas irreconciliables.
Para los feneticistas la clasificación era un asunto de ver y no de observar, la distinción aquí era para eliminar la "carga" teórica y conceptual que el resto de la ciencia incorporaba al quehacer clasificatorio tradicional. Ésta fue una posición que en sus orígenes sostuvieron los feneticistas o neoadansonianos, pero que tuvo que desecharse por ilusoria. Hablar de sexos, organismos, individuos, numerosas estructuras, formas, funciones, conductas y otros atributos de los seres vivos implícitamente incorporaba conceptos teóricos. Los numericistas no podían evadir las teorías científicas al considerar innumerables caracteres. No era posible reducir el conocimiento al mero ver y escudriñar, ya que las teorías no son reductibles a conjuntos de observaciones. La neutralidad teórica era una fantasía. La directriz teórica que sostenía las clasificaciones de los elementos, de los compuestos orgánicos e inorgánicos, de las estrellas y otras entidades, significaba una muestra de su desatinada concepción que pronto fue parcialmente abandonada.
Una segunda posición fue la de admitir sólo las teorías más aceptadas, es decir rechazar básicamente a la evolución como idea rectora de las decisiones clasificatorias y como fundamento de las relaciones taxonómicas. En esta segunda toma de posiciones se señaló que las interpretaciones evolutivas deberían basarse en las clasificaciones fenéticas. La sistemática fue concebida como una ciencia sin relación alguna con el tiempo y el espacio o con causa alguna, divorciada de los procesos y de las teorías que podían explicar a esos procesos.
Estas opiniones, que fundamentaban su forma de conceptuar a la sistemática, los obligaban a mantener que la valoración a priori de los caracteres era un error. La valoración de los caracteres a posteriori sería la forma adecuada de comparar la jerarquía de las propiedades en los seres vivos. No había razón, según ellos, para dar mayor valor o peso a los caracteres previamente al análisis de la semejanza total. El pesaje o la importancia de los caracteres debería demostrarse después d análisis de la semejanza, donde se consideraban con el mismo peso a todos; fuera un pelo, un estómago o una aleta, todos valían lo mismo a priori.
Para ellos la similitud era la homología; la comunidad de descendencia no tenía significado alguno para la homología, si no era evaluada a partir de un examen de caracteres. Los sokalianos confundían las similitudes análogas con las homólogas. Las primeras eran resultado de convergencias evolutivas y de homoplasias en general, las segundas resultaban de la proximidad de ancestría.
Uno de los taxónomos y evolucionistas más importantes de la corriente de la síntesis moderna de la evolución, Ernst Mayr, señaló el modo de razonamiento de los numericistas a través del ejemplo de los gemelos idénticos: "... dos hermanos no son gemelos idénticos debido a que ellos sean similares, sino que son similares debido a que se derivan de una sola célula, que es a lo que se debe que sean gemelos idénticos". Como puede advertirse, una falacia básica de los nominalistas es su interpretación desviada de la relación causal entre la similitud y la relación. En el caso de los gemelos, la relación de afinidad es lo que causa la similitud y no a la inversa.

Para los seguidores de Sneath y Sokal el grado de similitud hacia los grupos o taxa; ése era su fundamento más solicitado.
LAS ESPECIES MATEMÁTICAMENTE DEFINIDAS Y LOS BIÓLOGOS
RECOLECTORES DE VECTORES, MATRICES Y ECUACIONES
El reconocimiento de que en la especie existe variación, que no hay un solo organismo de la misma especie igual a otro y que si ocurriese podrían llegar a ser diferentes según sean las condiciones en donde se desarrollen, fueron conocimientos que se adquirieron con el abandono del concepto linneano de especie, un concepto esencialista. Darwin usó tal conocimiento para inventar su teoría de la selección natural. La genética de Mendel y los estudios poblacionales apoyaron su teoría con las consecuencias que se han señalado en el capítulo anterior, entre ellas, la de sustituir el concepto de especie esencialista con base en la idea de comunidad reproductiva y la separación de otras especies mediante mecanismos de aislamiento. Precisamente estas ideas demarcaron la noción de continuidad morfológica entre los distintos miembros de una especie, lo cual fomentó el uso de la estadística para reconocer en términos de números y medidas los caracteres de las especies. Se trataba de una respuesta de los feneticistas al criterio de autoridad con el que algunos taxónomos definían a la especie. La ausencia de un umbral de diferencia reconocible en la aplicación del concepto de especie biológica era lo que aparentemente permitía aceptar que había decisiones arbitrarias.
La opinión de los feneticistas de explicitar los procedimientos taxonómicos adquiría vigencia en la práctica del reconocimiento de especies, de su determinación o identificación y su clasificación. Las especies muy próximas en su parecido general y aquellas que llegaban a hibridizar planteaban serios problemas; los numericistas demandaron procedimientos explícitos que se traducían en echarle cuentas al asunto. Otros biólogos continuaron en dos senderos: 1) reconocer los atributos no estudiados que permitían diferenciar a las especies, esto es, ampliar el conocimiento biológico de las especies, "micromorfología, rasgos subcelulares y propiedades químicas" (este camino era el de Julian Huxley, quien opinaba que las especies podían ser de muy variados tipos y significancia en diferentes grupos; para él no había un criterio sencillo de especie), y 2) ya que el concepto de especie biológica tenía deficiencias en aspectos de estudios del proceso evolutivo, habría que perfeccionar un concepto de especie que sirviera para el análisis de la reconstrucción evolutiva, esto es, desarrollar un concepto filogenético de especie que tuviese aplicación en organismos que no presentaran sexualidad y, por ende, no tuvieran comunidad reproductiva.
Se requería inventar un nuevo concepto teórico de especie y un método para la reconstrucción genealógica. Como se señaló previamente, el filósofo de la biología David Hull expuso el problema en los términos siguientes: "Una cosa es el reconocimiento de la existencia de la variación entre formas contemporáneas como una consecuencia necesaria de la teoría sintética de la evolución, y otra es formular una metodología en taxonomía suficiente para manejar tal variación", e insistía:
La historia del concepto de especie
biológica es una historia de intentos sucesivos para definir la especie,
tal que los grupos resultantes sean unidades significativas en evolución.
|
Una primera definición de especie evolutiva fue propuesta por el paleontólogo George Gaylord Simpson. La carencia de la dimensión temporal en el concepto de especie biológica tenía limitaciones para el análisis del proceso evolutivo; por ello, Simpson incluyó en su definición el concepto de secuencia de poblaciones ancestro-descendientes, que sería desarrollado por otra escuela de pensamiento taxonómico, como se verá en el capítulo siguiente.
En la tendencia numericista se prosiguió con el desarrollo de importantes procedimientos estadísticos que aún hoy tienen gran aplicación en el estudio de la variación de las poblaciones y las especies. Desde luego, no fueron pocos los que usaron sus procedimientos para diferenciar a las especies en función de cantidad de parecido, una medida de distancia matemática. No había criterios biológicos en la demarcación de sus unidades.
Mientras que algunos, los feneticistas, continuaron con sus pretensiones de diferenciar a las especies con base en definiciones numéricas, los taxónomos evolutivos siguieron buscando razones biológicas para efectuarlas. Mientras que para los evolucionistas ciertos conceptos eran centrales a las teorías biológicas, para los feneticistas —que seguían una posición empirista—no tenían significado alguno, ya que éstos preferían una clasificación artificial basada en la mera aritmética de los caracteres que podía ser irrelevante biológicamente.
El proceso de especiación (generación de nuevas especies) era concebido como algo gradual de acuerdo con la teoría darwinista. En consecuencia, existía un continuo de especies incipientes que era más fácil diferenciar por métodos matemáticos, decían los neoadansonianos. Como la práctica era interpretar ciertas diferencias morfológicas en términos de discontinuidad reproductiva, entonces las medidas cuantitativas y métricas propuestas constituían una alternativa. En vez de especies hablaban de unidades taxonómicas operacionales, con su acrónimo: UTO's. Con las especies biológicas no podían operar, y consideraban que las prácticas subjetivas y arbitrarias en su reconocimiento prohibían su uso en la taxonomía numérica.
Los numericistas sólo reconocían individuos, puesto que los agrupamientos en términos de poblaciones, especies y grupos superiores, o todas las clases y conceptos de este tipo, sólo significaban para ellos artificios de la mente humana, meras convenciones o especulaciones teóricas que no guardaban un orden de operaciones para que tales conceptos pudieran ser reconocidos. Debido a ello, Sokal, Sneath, Colless, Camin y muchos otros feneticistas, señalaban que los nombres dados a los grupos de especies, por ejemplo: ratón, elefante, aves y muchos otros, únicamente eran nombres inventados por el hombre para grupos de individuos que él consideraba similares. Por tales puntos de vista también fueron llamados nominalistas.
LOS CAMINÁLCULOS Y LOS ARTIGÁLCULOS
A principios de la década de 1960 Joseph Camin, investigador de la Universidad de Kansas de los Estados Unidos, inventó un grupo de animales imaginarios, con el propósito de examinar las relaciones evolutivas entre ellos y obtener principios aplicables en la reconstrucción de la evolución de organismos reales, a través de las técnicas numéricas desarrolladas por los feneticistas. En honor e irreverencia simultánea, los 29 animales "actuales" y los 48 "fósiles" inventados fueron llamados caminálculos por los estudiantes de Camin. Tal modelo de animales había sido producido al seguir reglas que respetaran los principios evolutivos conocidos y que simularan el proceso de evolución real. Eran animales hipotéticos de los que su inventor sabía la "filogenia" bajo la cual se habían generado, pero los usuarios del modelo lo ignoraban. Animales menos complejos fueron inventados por Artigas en Argentina con fines equivalentes, a los que él mismo llamó artigálculos; una autoirreverencia inaudita en un argentino, aunque también puede interpretarse como una respuesta hacia la indiferencia dé los taxónomos latinos tradicionales, para no pasar inadvertido ante la moda.
De este modo los neoadansonianos deseaban examinar los diversos métodos de reconstrucción evolutiva que estaban en práctica, con el fin de contrastar la capacidad de alcanzar la "verdadera" filogenia por medio de distintos métodos. Se esperaba que los resultados de este modelo pudieran convencer a los taxónomos del poderío metodológico de la sistemática numérica; cuando menos eran de valor pedagógico y heurístico. Es importante mencionar que incluso feneticistas influyentes como Colless no admitieron el uso de los caminálculos para investigar la historia evolutiva, pues existía una gran aversión al estudio de la evolución. Por otra parte, esto muestra que no existe escuela de pensamiento en taxonomía que sea plenamente monolítica, ya que muchas ideas se imbrican; lo mismo ocurre con genealogistas y gradistas.
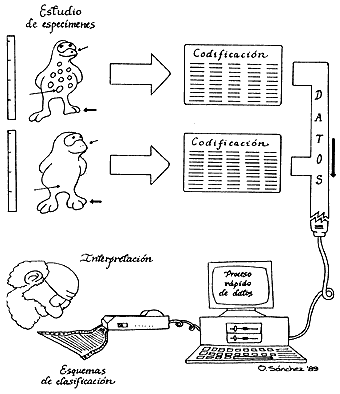
Los estudios efectuados condujeron a ramificaciones evolutivas diferentes, aunque los datos fueran consistentes —en cada método— con los resultados obtenidos. La elección de los distintos resultados no era fácil ni obvia. En la supuesta realidad subyacía una sola respuesta, una sola historia evolutiva; sin embargo, los esquemas que más se acercaban a la solución eran aquellos que presentaban el menor número de cambios evolutivos.
El procesamiento por medio de técnicas numéricas, ayudado por computadoras, se efectuaba con mayor rapidez, lo que les confería desde un principio ventajas adicionales.
El punto de partida de estos taxónomos, al no reconocer las especies y no asignar una valoración ponderada de los caracteres, hace la diferencia con investigadores posteriores que también han usado las matemáticas para hacer formales y rigurosos los análisis de la filogenia de organismos. La diferencia no se hace por el uso de las matemáticas, sino por los supuestos que conlleva en su uso.
Colless decía —como resultado de esta situación— que la taxonomía numérica a menudo erraba por la rigidez de confinar su atención hacia la taxonomía como mera tecnología, sin relacionarla a la ciencia de la sistemática a la cual debe servir.
Hay poca duda ahora de la enorme ayuda que las técnicas de la taxonomía numérica pueden ofrecer en este campo, sobre todo en la clasificación de entidades que no provienen de un proceso de ancestría-descendencia. En el estudio de la reconstrucción genealógica de las especies, hoy existen métodos más coherentes con el proceso evolutivo, tan robustos como las técnicas meramente numéricas y más consecuentes con las bases teóricas de la clasificación biológica.
La polémica aún no concluye, aunque ha menguado su ánimo original; recientemente, las proposiciones de los feneticistas son más moderadas y ya no significan la panacea como se propuso en un principio. No obstante, el impulso que generó la taxonomía numérica cumplió con el propósito de que los taxónomos fueran más rigurosos y formales en sus juicios; además, se logró avanzar en conceptos de gran valor para el estudio de las relaciones filogenéticas entre los seres vivos.
Con la introducción de la taxonomía numérica se insistió más en otros aspectos de las clasificaciones biológicas a los que se había atendido poco, por ejemplo: compatibilidad, congruencia y consenso de los caracteres, los diagramas de ramificación o las clasificaciones, predictibilidad de las clasificaciones, valor explicatorio de las clasificaciones y provisionalidad de tales explicaciones; pero, sobre todo, se discutió mucho si una clasificación de organismos debería reflejar la filogenia de éstos.
Es obvio que con las tecnologías disponibles actualmente, el incremento y la recuperación de información en las clasificaciones biológicas ha avanzado casi en términos fantásticos. La ayuda de medios electrónicos hace reales ciertas visiones futuristas de un cuarto de siglo atrás. No está lejano el día en que efectuemos "identificaciones" de organismos de forma automatizada, de modo expedito. Parece obvio que nuestra destreza en gran cantidad de tareas curatoriales aumentará con la disponibilidad de nuevos elementos tecnológicos; esto es, las tareas de obtención de muestras de seres vivos, su estudio y análisis en museos y laboratorios de investigación, progresarán conforme la ciencia y las tecnologías mejoren.
Los ideales feneticistas aún no desaparecen. Algunos podrían hacerse realidad con el enorme progreso de la informática y la robótica, al aplicarse a las diversas tareas taxonómicas que aún nos esperan.