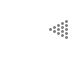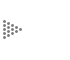VII. MADRE... SÓLO HAY UNA. HENNING, NELSON Y WILEY: PROPUESTA Y EVOLUCIÓN DEL MÉTODO PARA ELABORAR CLASIFICACIONES NATURALES
Los taxónomos son más parecidos a los artistas que a los críticos de arte, ellos practican su oficio y no lo discuten. He aquí un ejemplo de la actitud que sustentan ciertas corrientes frente a un examen profundo de las bases teóricas y los métodos de su ciencia. Pero, a pesar de ello, es evidente que la sistemática es una ciencia, y no un arte; para toda ciencia llega alguna vez el momento a partir del cual no hay progreso sin exámenes de tal naturaleza. Elementos de una sistemática filogenética (1968) WILLIE HENNIG El lenguaje de una ciencia debe ajustarse al contenido de la misma... Una de las tareas del biólogo es encontrar criterios racionales para agrupar juntos ciertos individuos o procesos y distinguirlos de otros... La sede del alma (1968) LONGUET-HIGGINS La objetividad debiera analizarse en función de criterios como la precisión y la coherencia. La tensión esencial (1982) THOMAS KHUN
A PESAR de la importancia práctica y teórica de la sistemática biológica, el estudio de otras ciencias experimentales en la biología había atraído la atención de los científicos y de quienes comenzaban a interesarse en la ciencia. Taxónomos de gran importancia como Willie Hennig opinaban de la sistemática que: "...no ha logrado una apropiada arquitectura doctrinal de sus problemas, tareas y métodos, acorde con la importancia real que tiene en el contexto general de las disciplinas biológicas".
Hacia finales de la década de 1960, Ernst Mayr sostenía que las tendencias en la sistemática se caracterizaban por tres líneas de desarrollo: 1) un gran interés en el examen de la teoría taxonómica, 2) el uso de las computadoras electrónicas y, 3) la introducción vigorosa de técnicas bioquímicas, con un gran interés en la comprensión de la filogenia de los organismos como punto de partida para entender la evolución de las macromoléculas.
Sin duda alguna es el primer aspecto el más trascendente en la sistemática, ya que las opiniones de los feneticistas, así como la proposición de un nuevo método y terminología para el análisis filogenético de los organismos cautivaban la atención de la mayoría de los taxónomos.
Desde luego no sólo se trataba de un método desprovisto de argumentos teóricos pues incluía conceptualizaciones diferentes respecto a la historia evolutiva de los organismos.
Si prácticamente no se había dicho nada nuevo y significativo sobre métodos, conceptos y teoría de la reconstrucción de la evolución de los organismos desde tiempos darwinianos, era obvio que lo que se propusiera causaría enorme prurito. El largo periodo sin examen de la teoría evolutiva causaría muchos desacuerdos; no iba a ser fácil convencer a los viejos sistematas ni a los nuevos e inexpertos deslumbrados por la impresionante tecnología que se ponía en boga. El lema evolutivo de corte darwiniano aún esta muy arraigado en los medios científicos.
No obstante, las debilidades metodológicas eran de tal magnitud que demandaban un serio debate sobre los fundamentos de la sistemática biológica; así, resultaba factible recibir proposiciones que desempantanaran el estilo arbitrario de las decisiones taxonómicas por un lado, como también sería posible examinar la relación entre historia evolutiva y clasificación. Se necesitaba dar una respuesta a la crítica de los feneticistas y superar las subjetividades del juicio taxonómico prevaleciente.
ESCINDIRSE O CAMBIARSE DE ROPAJES:
¿CUÁNDO CONSIGO SER DIFERENTE?
La disputa entre dos grupos de taxónomos que consideraban a la historia evolutiva de los organismos como elemento fundamental de las clasificaciones biológicas tuvo lugar a partir de la proposición que hiciera Hennig en su libro de 1950. Originalmente, la discusión no tuvo el ímpetu que debiera esperarse de acuerdo con el panorama que se ha planteado a grandes rasgos hasta este momento, pero tenía una explicación. El libro de Hennig originalmente se publicó en idioma alemán, lo cual dificultó mucho la lectura y el análisis para la mayoría de los taxónomos. Es por ello que hasta finales de la década de 1960 —cuando existía traducción al inglés y al castellano— se inició una de las polémicas más excitantes en el mundo de la biología y que aún no termina.
La rica discusión que se generó a partir de dicha polémica permitió la aclaración de numerosos conceptos y, como es frecuente en estas controversias, se llegaron a construir más conceptos con sus respectivos términos que los denotan. El calor de la discusión obligaba a eliminar la ambigüedad de las palabras que se usaban, pues sólo así sería posible avanzar; sin líos de significado que encubrieran las diferencias de concepción entre los contendientes intelectuales.
Por ejemplo, la palabra clasificación era usada con muy diversas connotaciones, y en algunos casos implicaba meramente un sistema de palabras, pero podía ser la monografía biológica entera. Lo anterior ocurría porque no se diferenciaba entre proceso y resultados del quehacer clasificatorio. Entonces, cuando se hablaba del aspecto de predictibilidad y recuperación de información en las clasificaciones, cuya discusión dependía de lo que se entendiera por clasificación, se podían prestar oídos sordos a los planteamientos de los taxónomos en pugna, sin acordar nada ante la carencia de términos claros.
La pregunta central en torno a la discusión, la que más discrepancia generaba entre los taxónomos de formación darwiniana, consistía en la precedencia de la genealogía de otro componente de la filogenia, como alternativas para la clasificación de referencia universal en la biología. Había aún numerosas confusiones conceptuales respecto a la relación de la historia evolutiva con la clasificación, lo que podría considerarse conveniente y lo que se aceptaba como natural.
Si una clasificación de organismos producía grandes asimetrías, si hacía inestable la nomenclatura en mayor o menor grado u otro problema, eran asuntos secundarios, posibles consecuencias del principal dilema planteado.
Resultaba claro que no había un método apropiado para la reconstrucción genealógica, y tampoco existía acuerdo en que la genealogía fuera el aspecto más fundamental de la filogenia. Habría de transcurrir más de una década para aclarar esta controversia. Entre las objeciones más importantes de los taxónomos tradicionales estaba precisamente el hecho de que sus principales representantes defendían que la genealogía sólo representaba uno de los procesos de la evolución. Argumentaban que la ramificación de los linajes o cladogénesis sólo es un proceso en filogénesis.
El mismo Hennig había desarrollado su teoría de reconstrucción filogenética con base en un concepto teórico que limitaba el análisis genealógico: el concepto de especie biológica. Era necesario inventar un concepto más general en términos de proceso filogenético que fuera utilizable —sin contradicciones— para el estudio de la cladogénesis o especialización, es decir, el origen de las distintas ramas evolutivas independientes que se habían generado durante la evolución.
Los puntos de vista de Hennig fueron fuertemente criticados, sobre todo en relación con la clasificación, y a que en un principio planteó un mecanismo muy simple de generación de nuevas especies, donde la especie generadora se extinguía después del evento generador de especies. Pero él arguía que sólo se trataba de una regla metodológica y no de un juicio sobre los hechos.
Las diferencias en concepción respecto a la filogenia y sus componentes estaban íntimamente relacionadas con la apreciación diferencial del valor de la cantidad de similitud-diferencia para reconocer grupos con una única historia evolutiva.
La historia evolutiva —filogenia— de un conjunto de organismos se caracteriza por poseer varios componentes: la dirección o polaridad evolutiva, la cantidad de cambio genético (cantidad de divergencia evolutiva o relaciones patrísticas), el tiempo de evolución (relaciones temporales o cronísticas), la diversificación o radiación evolutiva, la especiación o cladogénesis y las relaciones espaciales o biogeográficas. Los taxónomos tradicionales daban gran valor a la cantidad de divergencia debido a que decían que la magnitud de las diferencias (grado de divergencia) podía ser más importante que la proximidad de descendencia evolutiva. De aquí se deriva el nombre de gradistas. De hecho, con esto se generaban concepciones distintas de lo que significaba un grupo con una historia evolutiva única y su importancia para la reconstrucción filogenética y su aplicación o relación con la clasificación.
El ejemplo de los cocodrilos y las aves puede permitir un mejor entendimiento de las diferencias entre genealogistas vs. gradistas: aves y cocodrilos comparten un ancestro común inmediato, los seudosuquios (una especie de ellos, desde luego). Sin embargo, aves y cocodrilos han divergido tanto que a pesar de tener una historia común de descendencia, los gradistas los ubican en grupos separados mientras que los cladistas son consecuentes en este sentido y los colocan en un mismo grupo.
La razón es que la cantidad de divergencia es una cuestión de variación en
la velocidad de evolución, que es distinta para cada rama evolutiva en cada
especie o grupo de especies. Por lo tanto, una decisión sobre la cantidad de
divergencia es arbitraria y subjetiva, y no hay un umbral que nos permita ser
homogéneos en nuestras decisiones o resoluciones. ¿Qué tantas diferencias y
similitudes podemos reconocer y qué grado o porcentaje deberíamos admitir, para
respetar o no la descendencia común en el reconocimiento de grupos con una misma
historia? Los gradistas no tendrían una respuesta satisfactoria a esta pregunta
para sostener su punto de vista.

Estudios actuales muestran que, a pesar de sus diferencias aparentes, las
aves son parientes cercanas de los extintos dinosaurios, y hermanas de los cocodrilos.
De lo que se trata es de ubicar la diferencia entre dejar que la similitud por sí sola —como cantidad de cambio de un grupo a otro— tome precedencia sore el aspecto cualitativo de comunidad de descendencia, definida ésta sobre la base de una interpretación evolutiva de la similitud. Cantidad de diferencia vs. calidad de diferencia.
Ambas escuelas de pensamiento en sistemática biológica mantenían concepciones distintas respecto a la apreciación de lo que significaba una misma historia evolutiva.
Para los feneticistas la similitud total resolvía la naturalidad de los grupos a clasificar; para los gradistas era la comunidad de descendencia la que lo resolvía, siempre y cuando la cantidad de similitud se conservara dentro de ciertos márgenes, sin decir qué tanto, y para los genealogistas o cladistas la comunidad de descendencia —en donde todos los descendientes de un ancestro se integraban al grupo— resolvía la naturalidad, sin hacer excepciones. Debido a estas consideraciones, los feneticistas afirmaban que las clasificaciones de los gradistas mostraban una mezcla indefinida de grupos con relaciones fenéticas y cladistas.
En aspectos terminológicos los cladistas decían a los gradistas que su concepto de grupo con una misma historia de descendencia estaba mezclado, pues no era una definición estricta y consecuente. Cuando sus grupos incluían al ancestro y todos sus descendientes, eran realmente aceptables o monoféticos y servían para reconstruir los aspectos cualitativos más fundamentales de la evolución. Pero cuando sustraían de un grupo de descendientes, a una o más especies —por grado de divergencia—, estaban cercenando la unidad de historia, es decir, estaban formando grupos parafiléticos que no son aceptables para reconstruir los aspectos más estructurales de la evolución. Una clasificación que contuviera grupos parafiléticos reflejaría decisiones arbitrarias y subjetivas, por tanto, no podía funcionar como el sistema de referencia general para la biología, cuando justo eso se deseaba descubrir.
El quid del asunto estribaba en que decisiones arbitrarias sobre la cantidad de divergencia generarían más de una clasificación con supuestas bases evolutivas, aunque fueran pretendidamente naturales; mientras que las clasificaciones cladistas —con base en grupos estrictamente monofiléticos— garantizaban una sola clasificación, pues sólo hay una historia de descendencia de los organismos por descubrir.
La idea rectora de la teoría taxonómica de Hennig y algunos de sus descendientes intelectuales consistía en que las relaciones que permiten la cohesión de organismos y especies, vivientes y extintas, son relaciones genealógicas. Otros tipos de relación —fenotípicas y genéticas—, abarcan fenómenos correlacionados con la descendencia genealógica con modificación, y por ende, son mejor entendidos dentro de ese contexto. La similitud por sí sola, cualquiera que sea, no nos permite el establecimiento de relaciones genealógicas. Los caracteres deben ser interpretados a la luz de la evolución y sólo algunos de ellos pueden utilizarse para el descubrimiento de la ancestría común inmediata.
Hacia mediados de la década de 1970 la idea principal de Hennig había sido aceptada por la sistemática, y varios taxónomos evolutivos reconocían la importancia sin precedente que adquirían los conceptos de caracteres derivados compartidos para el análisis de las filogenias de organismos. Desde luego, no había acuerdo en una relación directa entre filogenia y clasificación; no obstante, sus implicaciones como fundamento de reconstrucción de historia no se ponían en duda. Pronto sus implicaciones en biogeografía y cronística —otros componentes de la filogenia— causaron grandes controversias que aún continúan en las revistas más prestigiadas acerca de la biología comparada.
En palabras del ya célebre divulgador de la ciencia, Stephen Jay Gould, el planteamiento del debate estaba en los términos siguientes:
Por desgracia, estos dos tipos de
información —el orden de ramificación y la similitud global— no
siempre dan resultados congruentes. Los cladistas rechazan la similitud
global como una trampa y una ilusión, y trabajan exclusivamente con el
orden de ramificación; los feneticistas intentan trabajar exclusivamente
con la similitud global e intentan medirla en una vana búsqueda de objetividad.
El sistema tradicional intenta equilibrar los dos tipos de información,
pero a menudo caé en una confusión imposible porque realmente ambos entran
en conflicto [...] Así, los cladistas compran su objetividad potencial
a cambio de 'ignorar' una información biológicamente importante, y los
tradicionalistas se arriesgan a la confusión y la subjetividad, al intentar
un equilibrio de dos fuentes de información legítimas aunque a menudo
dispares. ¿Qué se puede hacer? |

Grupos hermanos son aquellos que provienen de un ancestro común inmediato.
TIPOS DE CARACTERES EN FILOGENIA Y SU JERARQUÍA
Desde tiempos posdarwinianos y hasta la nueva sistemática, los caracteres utilizados para determinar ancestría común fueron los homólogos, aquellos que por definición provenían de una misma ruta de descendencia. Los caracteres convergentes provenían de ancestros distintos y, por ende, no servían para reconocer historia común de descendencia. Los caracteres que variaban de modo paralelo presentaban una enorme dificultad para distinguirlos de los convergentes, pero de cualquier modo la procedencia a partir de ancestros distintos los eliminaba, como evidencia útil a la definición de descendencia común.
Los feneticistas interpretaban los caracteres homólogos como producto de la similitud general que identificaban y evaluaban, y no como los gradistas la entendían, en el sentido tradicional de ancestría común. ¿Era la homología —así entendida— suficiente para descubrir grupos monofiléticos que permitieran reconocer la columna vertebral de la filogenia, esto es, la genealogía? La respuesta era no, pues se quería reconocer un tipo de homologías que ayudaran a descubrir una ancestría común inmediata, no cualquier tipo de ancestría. Se requería precisión en la postulación de posibles ancestros y los hermanos descendientes; había hipótesis generales e hipótesis particulares de descendencia genealógica, habría entonces homólogos para cada tipo de hipótesis. Se imponía efectuar una evaluación de los caracteres como indicadores del nivel de ancestría que documentaban en las relaciones filogenéticas de los organismos. La distribución de los homólogos y su jerarquía era la clave.
¿Pero qué eran los homólogos en la perspectiva de cambio evolutivo de los caracteres
en las especies? ¿Qué relación tenían respecto a los nuevos caracteres y las
novedades evolutivas? ¿Cómo era la ruta evolutiva de los caracteres? ¿Qué relevancia
filogenética podía tener el que se compartieran homólogos primitivos en lugar
de homólogos recientes? ¿Cuáles eran los caracteres que demostraban ancestría
común inmediata? Las respuestas a estas preguntas llevaron a la invención de
nuevos conceptos en análisis filogenético con sus correspondientes términos;
del mismo modo que los taxónomos numéricos habían denotado a sus conceptos con
nuevas palabras, lo mismo hicieron los hennigianos. Es lo que naturalmente ocurre
en la ciencia con el surgimiento de nuevas teorías y métodos; inventar y aprender
nuevos términos para los mismos conceptos, aunque sea en otro contexto teórico,
es algo que causa mucho disgusto incluso entre los científicos, pues eso fue
una de las críticas más sonadas hacia la sistemática cladista.

El razonamiento que siguió Hennig es relativamente sencilío, si se toma en
cuenta que la especiación es una transformación de los organismos en la que
los descendientes se tornan específicamente diferentes de los progenitores.
En el caso más simple, cuando una especie se divide y produce otra especie,
por lo menos hay un cambio o transformación de un carácter en la especie descendiente,
esto es, un carácter a cambia en a' (a ![]() a'). Si a su vez surge otro fenómeno de especiación
simple en la especie que es portadora de a, puede esperarse un cambio
de a' en a" (a'
a'). Si a su vez surge otro fenómeno de especiación
simple en la especie que es portadora de a, puede esperarse un cambio
de a' en a" (a' ![]() a"), pero si a' no se transforma en a",
un carácter b puede transformarse en b', incluso supóngase que
b estaba presente en la especie original que portaba el carácter a,
pero que sólo se transformó después del cambio a
a"), pero si a' no se transforma en a",
un carácter b puede transformarse en b', incluso supóngase que
b estaba presente en la especie original que portaba el carácter a,
pero que sólo se transformó después del cambio a ![]() a'. En la figura siguiente se ilustran estos cambios:
a'. En la figura siguiente se ilustran estos cambios:
En el ejemplo, el carácter pasa por una sucesión de estados diferentes desde
a hasta a" y de b en b', lo cual forma dos series
de transformación de estados de carácter que pueden escribirse así: a
![]() a'
a' ![]() a" y b
a" y b ![]() b'. Los estados de carácter originales en estas
series de transformación se denominan primitivos o plesiomorfos (a y
b), mientras que los subsiguientes se denominan derivados o apomorfos
(a', a" y b'). Estos conceptos sólo son utilizados en un sentido relativo,
esto es, cuando hay una serie de transformación conocida en la que los caracteres
se pueden comparar: a es plesiomorfo de a', pero a' es plesiomorfo
frente a a".
b'. Los estados de carácter originales en estas
series de transformación se denominan primitivos o plesiomorfos (a y
b), mientras que los subsiguientes se denominan derivados o apomorfos
(a', a" y b'). Estos conceptos sólo son utilizados en un sentido relativo,
esto es, cuando hay una serie de transformación conocida en la que los caracteres
se pueden comparar: a es plesiomorfo de a', pero a' es plesiomorfo
frente a a".
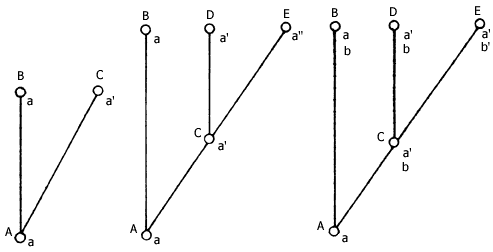
La explicación de este hecho y la posesión común de estos caracteres como resultado de la herencia, es lo que Hennig reconoció para el análisis filogenético. Vuélvase al ejemplo y adviértase que las especies B y D comparten el carácter b, por lo que se llama simplesiomorfo a un grupo unido por homologías primitivas. Las especies D y E están unidas con base en las apomorfías a' a", que son sinapomorfas respecto al carácter a', y es un grupo unido por caracteres derivados. No obstante, puede señalarse que a" es un carácter autapomórfico, como el de aquellos caracteres derivados que son exclusivos a un grupo o a una especie.
Con base en esta línea de razonamiento Hennig distinguió dos tipos de homologías: las plesiomórficas y las sinapomórficas; sólo las homologías sinapomórficas podían definir grupos monofiléticos. Obsérvese que si se sustrae a la especie E del grupo aquí ilustrado, como en el caso de las aves y los cocodrilos, se eliminaría a uno de los descendientes de un grupo monofilético, que sólo quedaría definido por el carácter b, que es una simplesiomorfía; por esa razón se determinó que un grupo parafilético está definido por simplesiomorfías. Era un grupo que, como puede advertirse, aunque sólo incluía taxa de una sola historia de descendencia, no contenía todas las especies que compartían esa historia.
Un carácter o atributo que aparecía por primera vez en un grupo de especies
se llamaba autapomorfía o novedad evolutiva, era un carácter derivado de un
ancestral. A partir de ahí, la autapomorfía, si se heredaba a las especies descendientes,
tomaba el papel de sinapomorfía respecto al carácter original; los cambios subsecuentes
en dos o más eventos de especiación convertían a la autapomorfía en un carácter
relativamente primitivo, esto es, una simplesiomorfía. La secuencia era la siguiente:
autapomorfía ![]() sinapomorfía
sinapomorfía ![]() simplesiomorfía, una secuencia que se determinaba en forma
relativa. No existen caracteres apomorfos sin relación a otro que le antecede
y es simplesiomorfo, en una misma línea de descendencia de homólogos. Los caracteres
autapomorfos, simplesiomorfos y sinapomorfos no existen por sí solos, sino en
relación con otros en una misma ruta de descendencia con modificación.
simplesiomorfía, una secuencia que se determinaba en forma
relativa. No existen caracteres apomorfos sin relación a otro que le antecede
y es simplesiomorfo, en una misma línea de descendencia de homólogos. Los caracteres
autapomorfos, simplesiomorfos y sinapomorfos no existen por sí solos, sino en
relación con otros en una misma ruta de descendencia con modificación.
Se había puesto al descubierto que la semejanza o parecido en los seres vivos podía deberse a sinapomorfías, simplesiomorfías y convergencias (también reversiones y paralelismo). Tres rutas evolutivas distintas que explicaban el parecido en los atributos de los organismos. Cuando el parecido era resultado de convergencias evolutivas significaba que se reunían grupos con ancestros distintos, por lo que se denominaban grupos polifiléticos, esto es, de origen en distintas ramas, el mismo caso para reversiones y paralelismo.
Sin embargo, el método era aún incompleto, pues faltaba aclarar cómo se determinaba
el origen de una serie de transformación en el caso de conocer sólo la relación
de caracteres en nuestro ejemplo a ![]() a'
a' ![]() a", sin
las flechas correspondientes que señalasen el orden de los cambios.
a", sin
las flechas correspondientes que señalasen el orden de los cambios.
¿Cómo se determinarían las homologías? La definición teórica era impracticable, pues si se definían como toda similitud debida a un antecesor común, no se daría una verdadera solución, pues no se conocían a los antecesores independientemente del análisis de las similitudes.
Las reglas o consejos para determinar la homología usadas desde tiempos de Darwin y Haeckel podían ser usadas para plantear hipótesis de relación genealógica: 1) sucesión estratigráfica o paleontológica; 2) sucesión del desarrollo ontogenético; 3) correlación de series de transformación, y 4) anatomía comparada (posición morfológica, calidad de parecido y similitud por formas intermediarias). ¿Pero cómo examinar estas hipótesis sin llegar a razonamientos circulares?
En nuestro ejemplo, todas las especies y caracteres estaban contenidos en un conjunto sin referencia a otros. Confiamos en los caracteres a y b como los más primitivos, ¿pero qué ocurriría si nuestro carácter a' o b' se encontrase en una especie cercana a B, D o E?, nuestra serie de transformación de caracteres estaría equivocada y nuestras conclusiones acerca de la ancestría-descendencia de las especies no serían las correctas.
Precisamente, este elemento de análisis —aparentemente tan sencillo— fue el que Hennig introdujo para examinar las hipótesis de parentesco. Las hipótesis de ancestría-descendencia entre los organismos podían probarse de una manera independiente a las reglas o consejos que Darwin daba para generarlas, y así podía evitarse el razonamiento circular; el análisis de la similitud era el examen de la distribución de los caracteres homólogos en los taxa. Esta regla se conoció como regla del grupo externo y al método general que propuso Hennig se le llamó método de posición filogenética. ¿Acaso se había alcanzado al fin el método natural que con tanta ansia se había perseguido a través del tiempo? Cuando menos se había inventado un método consecuente con las ideas evolutivas prevalecientes, un método extraordinariamente persuasivo y convincente, y la genealogía era la mejor consecuencia de las relaciones de ancestría-descendencia entre los organismos.
Lo que se ha mostrado hasta aquí es que las relaciones de ancestro-descendiente sólo pueden documentarse o demostrarse con base en las especies descendientes, esto es, con base en la similitud compartida que demuestre una hermandad de especies. Son las hermanas las que presentan las sinapomorfías, que son el elemento de demostración en el problema.
Otro aspecto que hay que observar es que ahora los fósiles no funcionan como la prueba dura de las relaciones genealógicas. Los fósiles, como cualquier otro tipo de especies, cualquiera que sea la evidencia que la apoye, deben probar su posición filogenética. La idea de que un fósil siempre es un ancestro, un eslabón perdido, tiende a ser reconsiderada con la posibilidad de que se trate de un grupo hermano extinto, de acuerdo con el método de análisis cladístico mostrado aquí. Un fósil prueba relaciones cronísticas, aunque nunca muestra edades absolutas, sólo mínimas.
Aquellos que insistían e insisten en que la filogenia incorpora varios componentes tan importantes como la genealogía pueden tener una respuesta más detallada de acuerdo con el conocimiento que se ha alcanzado en teoría evolutiva: el proceso anagenético, el que causa la divergencia de los caracteres o disimilitud, puede ser independiente al proceso cladogenético, la especiación; aunque también puede causar la generación de nuevas especies al avanzar simultáneamente de modo gradual, por medio de la acumulación de cambios.
¿HACIA UN NUEVO CONCEPTO DE ESPECIE?
El taxónomo, biogeógrafo y evolucionista E. Wiley, discípulo en el departamento de ictiología del Museo de Historia Natural de Nueva York en la década de 1970, es hoy investigador en la Universidad de Kansas. Ha sido un incesante estudioso de la biología comparada y propuso una revisión del concepto de especie hace escasos años. Señaló que, con toda probabilidad, no hay tema en sistemática y evolución que haya consumido más papel que los trabajos sobre la naturaleza y la definición de la especie.
La controvertida palabra 'especie' en biología ha pasado por uno y mil debates, no obstante sigue siendo la misma. El problema del origen de las especies y la conceptualización de lo que es una especie, es desde cierta perspectiva un mismo problema sobre el que han girado cientos de disputas en la historia de la biología.
De cualquier manera, el concepto de la especie es igualmente problemático que la mayoría de los conceptos en la ciencia. Aquel que dice tener una idea de la objetividad absoluta en la ciencia tiene una concepción ahistórica y fundamentos desviados del quehacer científico; a menudo se trata de personas plenas de fe, en busca de una religión a la moda.
Las líneas anteriores quieren decir que nadie debe creer la última y definitiva palabra acerca de la especie, pues no hay finalismos en la ciencia. Debe esperarse una retroalimentación entre el estudio de la generación de especies las teorías de los procesos biológicos y el concepto de especie mismo, como ha ocurrido siempre.
El concepto de especie más popular en los últimos cincuenta años es concebido como un conjunto de poblaciones que integran una comunidad reproductiva separada de otras comunidades reproductivas por mecanismos de aislamiento, es decir, propiedades morfológicas, fisiológicas, genéticas, conductuales y otras, que impiden a las poblaciones de distintas especies que pierdan su identidad mediante la fusión o una hibridación poblacional completa.
Este concepto carece de una relación con el factor tiempo y el proceso de herencia de los caracteres; por ende, carece de aspectos de relación de ancestría-descendencia de las poblaciones. Al estar fundamentado en las relaciones reproductivas y, por consiguiente, en términos de sexualidad como vehículo que permite el flujo del material hereditario, deja de lado aspectos más generales en la biología de las poblaciones. Es un concepto que hace énfasis únicamente en los contenidos genéticos de las poblaciones, y no toma en cuenta los aspectos filogenéticos.
De Aristóteles a Linneo, de Darwin a Mayr, siempre se ha deseado tener un concepto universal de la especie aplicable a todos los seres vivos, en función de los hechos y teorías más lógicos, aceptados y disponibles hasta ese momento. Entonces, la proposición de un nuevo concepto de especie debería pretender una aplicabilidad que considere el parámetro tiempo, la herencia de los caracteres e incluya organismos que no presentan sexualidad. En este caso debiera considerar el flujo horizontal de material hereditario, no el vertical de padres a hijos, sino el de cualquier otro organismo que es capaz de incorporar material hereditario a otro organismo.
Un nuevo concepto de especie debe probar que, dado un grupo de organismos cualquiera, se puede comprender o no a una especie. Debe permitir el trabajo con especies reconocidas en función de sus propiedades genéticas, fenéticas, espaciales, temporales, ecofisiológicas y conductuales. También debe tomar en cuenta las rutas posibles de generación de especies.
A partir de una definición del paleontólogo George Gaylord Simpson, Wiley efectuó la siguiente: una especie es un solo linaje de poblaciones de organismos ancestro-descendientes que mantienen su identidad de otros linajes y tienen sus propias tendencias evolutivas y destino histórico. Linaje significa una sola o una serie de poblaciones pequeñas que comparten una historia común de descendencia no compartida por otras poblaciones, e identidad es un conjunto de cualidades que una entidad posee como producto de su origen y su capacidad para permanecer distinta y aislada evolutivamente de otras entidades. Puede fácilmente advertirse que es un concepto muy abstracto y difícil.
Una crítica constante a los conceptos de especie proviene de aquellos biólogos que trabajan con conjuntos de seres vivos ausentes de reproducción sexual, en donde la cohesión de los organismos en las poblaciones en una misma especie puede deberse a otros mecanismos distintos; en nuestra definición parece que otra vez ocurre lo mismo. Sin embargo, observe el lector que tal vez no se trata de lo inadecuado del concepto de especie evolutiva, sino que puede deberse a nuestras limitaciones de observación y método. Con las técnicas de estudios de macromoléculas (proteínas y DNA), su sucesión evolutiva y los métodos de comparación cuantitativa tal vez sea factible resolver este problema práctico.
Para terminar este capítulo, es importante referir una implicación lógica del concepto de especie evolutiva en relación con la especiación. Los paleontólogos a menudo clasifican un mismo linaje de poblaciones ancestro-descendientes como especies distintas. Este problema fue reconocido por Mayr y Simpson. La contradicción es clara cuando se reconoce que a menor cantidad de fósiles (datos) es más fácil la decisión de separar a las especies paleóntológicas; a mayor cantidad de fósiles tendríamos un continuo que haría más arbitraria nuestra decisión de división de especies. La ausencia de objetividad en estos procedimientos ha provocado que cada vez más biólogos rechacen a las paleoespecies subdivididas arbitrariamente.