VIII. MÁS ALLÁ DEL MÉTODO
La cladística no es la panacea... pero... ahora tenemos una metodología explícita que nos permite evaluar hipótesis de monofilia. Sistemática y paleobiología (1985) NILES ELDREDGE y MICHAEL NOVACECK No ha habido ningún debate más intenso en el seno de la teoría evolutiva en la última década que el desafío planteado por los cladistas a los sistemas tradicionales de clasificación. El problema surge de la complejidad del mundo, no de la confusión del pensamiento humano, aunque ésta también ha hecho su habitual contribución. Dientes de gallina y dedos de caballo (1984) STEPHEN JAY GOULD No tenemos forma de evaluar científicamente nuestros métodos. Puede parecer paradójico sugerir que la biología comparada, o cualquier ciencia, debe adoptar métodos sin que pueda ser capaz de evaluar su eficacia. Pero el hecho es que usamos nuestros métodos en un intento de resolver los problemas. Si ya conocemos las soluciones correctas a esos problemas, fácilmente podemos evaluar y elegir entre varias metodologías en competencia: aquellos métodos que consistentemente ofrezcan las soluciones correctas obviamente serán preferidos. Pero, desde luego, si ya conociéramos las soluciones correctas, no tendríamos la necesidad de los métodos. Sistemática y biogeografía (1981) GARETH NELSON y NORMAN PLATNICK La idea de un método que contenga principios científicos, inalterables y absolutamente obligatorios que rijan los asuntos científicos, entra en dificultades al ser confrontada con los resultados de la investigación histórica. En ese momento nos encontramos con que no hay una sola regla, por plausible que sea, ni por firmemente basada en la epistemología que venga, que no sea infringida en una ocasión o en otra. Llega a ser evidente que tales infracciones no ocurren accidentalmente, que no son el resultado del conocimiento insuficiente o de una falta de atención que pudieran haberse evitado. Contra el método (1981) PAUL FEYERABEND
OCURRE frecuentemente que investigaciones más profundas y descubrimientos de cierta relevancia revolucionan el conocimiento acerca de los objetos o fenómenos bajo estudio; esto acontece a tal grado, que nos obliga a reevaluar los métodos en aplicación, las unidades naturales utilizadas y nuestras concepciones de tales entidades.
El origen, las interrelaciones y sus líneas de desarrollo son puestos nuevamente en la mesa de las discusiones; así, se inicia un nuevo debate sobre las implicaciones que los nuevos hallazgos producen.
En nuestro caso, los estudios en biología molecular han permitido el análisis del origen de varios grupos relativamente antiguos y poco complejos, que posiblemente fueron de los primeros organismos existentes en la Tierra, tales como una variedad de bacterias, virus, algunos organelos celulares: la mitocondria y el cloroplasto, y hasta la misma investigación del origen de los ácidos nucleicos —desoxirribonucleico (DNA) y ribonucleico (RNA)— que se encuentran en todos los seres vivos, lo cual ha conducido a un serio examen de sus orígenes, interrelaciones y evolución.
Los temas de estudio en esa área de investigación se encuentran en la frontera del conocimiento actual y no es fácil transmitir algo de lo que apenas se tienen algunas respuestas. Sin embargo, es necesario dar cuenta de algunas de éstas, pues nos conducirán a las nuevas avenidas de la ciencia que hoy y en un futuro cercano se construirán.
Simbiosis. Una teoría de los setenta que ahora se tiene bien apoyada con evidencia de biología molecular, es la que plantea el origen de los organelos en la célula "típica" o eucarionte: membrana nuclear bien definida y citoplasma con organelos.
En cualquier texto de biología del nivel secundario se acepta ya la teoría endosimbiótica propuesta por Lynn Margulis, la cual afirma que organelos como la mitocondria y el cloroplasto en algún tiempo fueron bacterias de vida libre que paulatinamente se integraron como parte de un consorcio celular. Dentro de ésta, endosimbióticamente, evolucionaron al especializarse y perder varias de sus propiedades originales; sin embargo, la existencia de DNA ha permitido el estudio de sus interrelaciones y posibles orígenes, por lo cual se sabe de sus relaciones y origen bacterianos. Es decir, la organización entre varias bacterias con distintas propiedades metabólicas condujo a la formación de los primeros eucariontes.
Es difícil que un suceso evolutivo complejo, como el origen de la célula con núcleo, haya ocurrido en varias ocasiones en la filogenia de la vida; a pesar de esto, no es posible dejar de cuestionarnos acerca de su origen "no monofilético", lo cual pone en tela de juicio en cierto modo nuestras ideas de monofilia o historia de descendencia única para todos los organismos. No obstante, la alternativa de varios sucesos pudiese quedar abierta.
Las células eucariontes representan la asociación de varias entidades con historias de descendencia distintas —células procariontes— que se han integrado en una sola, a partir de la cual ha descendido el resto de los eucariontes.
Una hipótesis alternativa, en la que tal evento pudiera haber acontecido varias veces y a distintos tiempos durante la historia terrestre, causaría un cisma mayor en nuestras concepciones y dificultaría el análisis filogenético de tales casos. Para fortuna de la investigación biológica, no existen datos que sugieran la posibilidad de tal hipótesis, de modo que por ahora hay cierta seguridad en la concepción monofilética de los eucariontes.
De modo contrario, si la hipótesis alternativa señalada tuviera posibilidades, el método inventado por Hennig no podría aplicarse con plena confianza y certidumbre, ya que primero se tendría que examinar la generalidad de su aplicación. Esto es debido a que el problema no es sólo el evento evolutivo reconocido por Margulis, sino que si estas endosimbiosis exitosas se hubieran repetido, entonces nuestras intenciones de encontrar grupos de organismos con historias de descendencia únicas se convierten en irrelevantes o, cuando menos, nuestro problema se vuelve más complejo. Por ello, es muy importante saber qué tan extendido está ese fenómeno de endosimbiósis, para aplicar nuestro método a las entidades correctas, sean organismos completos o parte de éstos.
Es imprescindible reconocer que los organismos no existen de modo natural con independencia de otros; nuestros conceptos de ecosistema y de comunidad nos previenen de esto y nos obligan a pensar en los sistemas de relaciones que mantienen los organismos. Algunas de éstas significan relaciones interdependientes, que son producto de muchos millones de años de evolución. Con base en lo anterior podemos explicarnos casos como el de los líquenes, que debieran considerarse como una asociación endosimbiótica entre una alga y un hongo, es decir, dos entidades cuyos componentes tienen ancestros distintos, pero que para funcionar como organismos (líquenes) necesitan integrarse, haciéndose complementarios. Por supuesto, también pueden existir de manera independiente, como individuos que pertenecen a especies de taxa bastante diferentes.
Como el caso anterior, también pudiera considerarse, en cierto modo, la íntima asociación de algunas bacterias fijadoras de nitrógeno (Rhizobium) con varias plantas leguminosas; por ejemplo: el frijol (Phaseolus). En éste y en muchos otros casos de parasitismo, la dependencia o interdependencia de la relación decide la existencia y la descendencia de los organismos comprendidos. Cada elemento que se integra a la asociación tiene una historia; el patrón de historia coevolutiva puede examinarse para cada constituyente. En otras palabras, cada elemento tiene una genealogía reconocible.
La aplicación del método hennigiano no tiene una aplicación universal a todos los organismos, pues requiere que éstos representen historias de descendencia únicas, de lo contrario la búsqueda se hace intrascendente. Su aplicación exige el reconocimiento de los posibles eventos endosimbióticos ocurridos en la formación de organismos durante la historia de la vida en la Tierra.
No es una debilidad del método, sino simplemente que se deben admitir otros hechos ecológicos y evolutivos, los cuales nuestro método no puede abordar libremente. Los procedimientos hennigianos no fallarían si se aplicaran a los constituyentes del organismo endosimbiótico si lo que se quisiera fuera encontrar las líneas de relación y descendencia evolutiva de los organelos con sus antecesores de vida libre. Igualmente, nuestro método no falla si se aplica a cualquier caso de organismos con historias monofiléticas, e incluso, podría servir para reconocer patrones reticulados.
Plásmidos y episomas. Desde la década de 1960 se descubrió la presencia de DNA de origen distinto dentro de las células procariontes y asociado a su cromosoma bacteriano. En algunos casos se supo que eran de origen viral, en otros casos se les supuso un origen bacteriano a partir de otras células de la misma especie. Cualquiera de ambas hipótesis planteaba el hecho de un movimiento horizontal del material genético entre los organismos.
Con tal fenómeno al descubierto se reconocía la posibilidad de que el material genético no se heredara o moviera sólo de padres a hijos, de ancestro a descendiente, es decir verticalmente, sino que también había otro movimiento posible: el horizontal.
Se planteaban varias incógnitas: ¿Qué tan extendido se hallaba este fenómeno entre los seres vivientes? ¿La migración horizontal de DNA se presentaba sólo en organismos procariontes? ¿Era posible esperar tales movimientos horizontales entre especies distintas? ¿Cualquier clase de información del DNA podía moverse entre las especies? Para esta clase de preguntas existen ahora algunas respuestas parciales, las cuales han pueso en advertencia la aplicación generalizada del método, que no parece ser apropiado para casos de movimiento de caracteres entre los descendientes.
No se sabe aún la extensión y la magnitud de tales fenómenos, aunque se sabe que existen. En algunos casos virales están más o menos bien reconocidos, pero todavía se ignora mucho en este terreno y se requiere mucha más investigación sobre esta clase de tópicos. Las implicaciones para la historia evolutiva pueden ser significativas y, al menos, nos podrán prevenir sobre el uso indiscriminado de algunos caracteres que —supuestamente— se planteaban como superiores para el análisis filogenético.
En el caso de los virus, que casi siempre están asociados a determinados grupos de especies, pudiera tratarse de entidades coevolucionadas con los grupos a los que están asociados, del mismo modo que los líquenes; sin embargo, otros casos de movimiento horizontal, si los hubiera entre eucariontes, serían de gran interés para el análisis de su genealogía.
El fenómeno genealógico de los seres vivos, que genera el patrón de referencia para la biología, puede verse obstruido metodológicamente debido a que el método hennigiano basado en el análisis de caracteres —fundado en el axioma de descendencia vertical de los caracteres—, no da cuenta de fenómenos de movimiento horizontal.
Hoy se acepta ya el hecho de que no toda la genealogía de los organismos es divergente, sino que en muchos casos hay cierto grado de reticulación, que puede ser mucho más pronunciado en organismos procariontes. Sin embargo, la posible migración de caracteres considerados sinapomórficos en una hipótesis de descendencia por analizar, previene en la aplicación indiscriminada del método hennigiano.
Recuérdese que el medio para descubrir hipótesis de descendencia únicas es mediante el análisis de los caracteres del modo planteado por Hennig, pero éste funciona bajo el axioma de que los caracteres se mueven verticalmente, de padres a hijos; si se viola esta suposición nuestro método cae en dificultades, pues se requiere saber qué tipo de caracteres pasan por movimiento horizontal para evitar su uso en el análisis de caracteres.
Fenómenos inexplorados. Como los dos casos anteriores, numerosos fenómenos más se han dado a conocer respecto a los ácidos nucleicos, de gran significancia para aspectos taxonómicos, ya sea para el reconocimiento de especies, el análisis filogenético o la clasificación de organismos.
La investigación sobre la regulación de la expresión del DNA, su duplicación, constancia, repetitividad y modificación, así como muchos otros aspectos, ha puesto al descubierto una amplia gama de hechos de importancia potencial en taxonomía.
A pesar de todos estos descubrimientos, la historia de la taxonomía ha dejado a sus practicantes el entendido de que no hay que perder de vista la interrelación de nuestras tareas fundamentales: 1) la clasificación de organismos con base en su genealogía, y 2) la distribución de las propiedades, caracteres o atributos entre los organismos. Ambas tareas se interrelacionan aún más debido a los fenómenos macromoleculares que se han descubierto en tiempos recientes. Se requiere clasificar organismos que pertenecen a especies que se crean a partir de un proceso de ancestría-descendencia con modificación, donde los ácidos nucleicos son fundamentales pero no son los únicos. Como caracteres, no están bien conocidos aún, pues por los resultados recientes se considera que todavía no termina la larga cadena de descubrimientos espectaculares.
El uso de la adjetivación respecto a los caracteres, es decir, la aplicación de la terminología y las consecuencias de ésta a los caracteres, no es del todo posible en los ácidos nucleicos y por ende se dificulta la aplicación del método.
Bajo esta perspectiva, puede observarse que el método y sus posibilidades no
están en entredicho; éste tiene un uso más o menos bien delimitado, pero lo
que se cuestiona es la utilización indiscriminada de los ácidos nucleicos con
un conocimiento aún inadecuado de ellos.
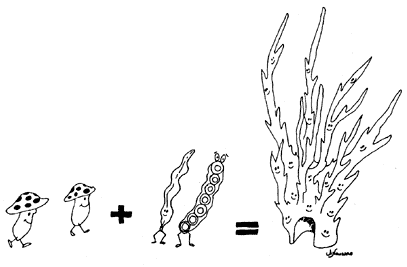
Los líquenes son una asociación entre un alga y un hongo; dos entidades
cuyos componentes tienen ancestros distintos, pero que para funcionar como organismos
necesitan integrarse, haciéndose complementarios...
Bajo otra perspectiva puede advertirse que el cuestionamiento al que conducen los descubrimientos recientes de movimiento horizontal del DNA apunta hacia la idea de que los caracteres se heredan de ancestro a descendientes, pero se ignora el movimiento genético entre organismos de especies distintas, con lo cual el método debiera caer en desuso y, tal vez, sería mejor regresar a la idea de semejanza total de los feneticistas, ya que la posibilidad de reconocer la genealogía de los organismos no es más que una quimera.
Mientras hay más resultados, la práctica consiste en el empleo del método hennigiano para caracteres morfológicos, sean micro o macro, externos o internos, de juveniles o de adultos; y por otra parte, el análisis de caracteres macromoleculares (ácidos nucleicos y proteínas). Ambos deben ser congruentes y una incongruencia indica datos erróneos o bien método o decisiones arbitrarias, pues a pesar del dilema planteado por el movimiento horizontal del DNA hay una sola historia de los hechos de interrelación filogenética entre los organismos, la cual está por descubrirse.
Por el momento, muchos biólogos moleculares no están preocupados por este problema metodológico, y con frecuencia establecen interrelaciones mediante métodos hoy en desuso o de bajo poder de resolución entre los taxónomos modernos. Aunque siendo justos en la crítica, muchos taxónomos "modernos" practican métodos anticuados e improcedentes para los problemas que los ocupan. Nos espera aún una polémica incesante, plena de vigor y creatividad, sobre todo si se recuperan las enseñanzas de la historia de la taxonomía y si se tratan de hacer preguntas de relevancia a los nuevos problemas planteados.
Sin menospreciar el debate internacional de estos problemas científicos, pues éstos determinarán los quehaceres próximos, tenemos ante nosotros una situación harto compleja. Queremos ser partícipes creativos de la ciencia, pero en México nos espera la resolución de problemas más preocupantes, pues somos testigos del decaimiento de la taxonomía y de su falta de madurez en la última década. La causa de esto ha sido también la falta de madurez de la ciencia mexicana, pero sin duda alguna también lo ha sido la incomprensión de otros colegas científicos y la carencia de medios y recursos para su desarrollo.
Señalo con convicción que se requiere hacer un gran esfuerzo institucional y colectivo para construir un gran museo nacional de historia natural, a través del cual se pueda hacer realidad el cultivo y la difusión de una ciencia fundamental para el desarrollo de la biología, una disciplina indispensable para un conocimiento adecuado de los recursos naturales y culturales de la nación.