IV. LA SINAPSIS: ESTRUCTURA Y FUNCIÓN
AL SITIO DE COMUNICACIÓN entre dos neuronas se le conoce como sinapsis. No se trata de un contacto directo, puesto que existe una separación infinitesimal entre las dos células, sino del punto en el que las dos células muestran, con el microscopio electrónico, áreas especializadas identificables tanto a nivel de la membrana celular como del interior y donde ocurre la transferencia de información entre dos células nerviosas. En el caso de la célula que "envía" la señal, nos referimos a la terminación presináptica (axonal). La neurona que recibe esa información representa la porción postsináptica (dendrítica). La parte distal del axón muestra un engrosamiento en forma de botón, en cuyo interior podemos encontrar mitocondrias (para el aporte de energía) y pequeñas vesículas que contienen moléculas de neurotransmisor (que discutiremos más adelante). Al otro lado hay dendritas con forma de espina, a las que la terminación axónica puede asociarse, ya sea en su parte terminal (cabeza) o en la unión con la dendrita principal (cuello). En muchos casos podemos identificar esta porción postsináptica por la presencia de una capa más densa localizada justo al lado opuesto de la presinapsis, como veremos más adelante. Este espesamiento o densidad postsináptica puede contener las sustancias receptoras que interactúan con los neurotransmisores liberados desde la presinapsis.
Existen varios tipos de sinapsis: por una parte las llamadas químicas, que ocuparán la mayor parte de nuestra atención aquí, pues en ellas actúan los fármacos que nos interesan. Existen también las sinapsis eléctricas que representan sitios donde las membranas de las dos neuronas están casi juntas. Es decir, no se observa (todo esto con el microscopio electrónico) ninguna hendidura o brecha entre las células. En estas sinapsis, el impulso nervioso pasa de una célula a otra manteniendo su forma eléctrica, sin pasar por una transformación de fuerzas químicas. En este último caso, se habla de transmisión neurohumoral. Anatómicamente, podemos referirnos a sinapsis axodendríticas, aquellas en las que el impulso nervioso parte del cuerpo celular y viaja hacia la periferia, para establecer comunicación a nivel de las dendritas (que son las que hemos mencionado); asimismo, podemos hablar de sinapsis axoaxónicas, axosomáticas (de un axón al cuerpo de una neurona) e incluso dendrodendríticas.
Los impulsos nerviosos provocan respuestas en el músculo liso (el de los vasos y vísceras), en el cardiaco, el esquelético, las glándulas exocrinas (las que vacían su contenido al exterior) y en las porciones postsinápticas de las neuronas por medio de la liberación de neurotransmisores químicos específicos. El descubrimiento de esta forma de neurotransmisión química modificó profundamente nuestra concepción sobre el funcionamiento del cerebro y estableció las bases para la comprensión de los mecanismos íntimos de los efectos farmacológicos. Vale la pena ver un poco en detalle el proceso seguido por los científicos para descubrir los diferentes pasos de la transmisión nerviosa.
La historia parece haber comenzado alrededor del año 1700, cuando Luigi Galvani notó que las ancas de rana que colgaban de ganchos de cobre en un balcón de fierro de vez en cuando se contraían solas, como si todavía estuvieran vivas. A partir de estas observaciones y de experimentos posteriores, Galvani concluyó que la fuerza que hacía mover los músculos y que existía en el sistema nervioso era electricidad, y no los "espíritus animales", idea propuesta más de mil años antes por Galeno. A partir de ese momento se utilizó la electricidad en experimentos fisiológicos para estimular todo tipo de tejido, y algo más de un siglo después se trataba de detectar y registrar (recordemos los experimentos de Einthoven), con galvanómetros de cuerda, los impulsos cardiacos, y que fueron los precursores de la electrocardiografía.
Entre 1898 y 1901, Lewandowsky por un lado, y Langley por otro, notaron en forma independiente la similitud entre los efectos de la inyección de extractos de glándulas suprarrenales y los de la estimulación eléctrica de nervios simpáticos. Pocos años después (1905), Elliot, alumno de Langley, propuso que la estimulación eléctrica de los nervios simpáticos producía la liberación de pequeñas cantidades de adrenalina, la hormona encontrada en las glándulas suprarrenales. También observó que este órgano permanecía sensible a la adrenalina aun después de que los nervios hubieran degenerado. Pensó entonces que existían sustancias receptoras de tipo excitador o inhibidor en el órgano efector, y que la acción de la adrenalina dependía de cualquiera de estas dos sustancias predominaba. En 1907, Dixon notó la impresionante similitud entre los efectos del alcaloide muscarina y los de la estimulación del nervio vago y propuso que este nervio liberaba una sustancia parecida a la muscarina para producir sus efectos. Ese mismo año Hunt describía los efectos de la acetilcolina y otros ésteres de la colina. En 1914 sir Henry Dale investigó las propiedades farmacológicas de la acetilcolina, y se sorprendía de la fidelidad con la que esta sustancia reproducía las respuestas a la estimulación eléctrica de nervios parasimpáticos. Observó además que la acetilcolina actuaba durante poco tiempo, por lo que propuso la existencia de otra sustancia (una enzima) que fraccionaba a la molécula para terminar su acción.
En 1921 Otto Loewi realizó un experimento crítico: el líquido con el que había perfundido un corazón de rana estimulado eléctricamente, y a través del nervio vago, lo hizo circular por un segundo corazón no estimulado. Dicha estimulación produce usualmente una lentificación de la frecuencia cardiaca. Loewi notó que el líquido del primer corazón perfundido producía lentificación de la frecuencia del segundo corazón y dedujo que se había liberado alguna sustancia por la estimulación del nervio vago a partir del corazón estimulado. Denominó a esta sustancia vagusstoff y cinco años más tarde mostró que se trataba de la acetilcolina.
También en 1921, Walter Cannon reportó que la estimulación de los nervios simpáticos del hígado producía la liberación de una sustancia parecida a la adrenalina que aumentaba la presión arterial y la frecuencia cardiaca. A esta "simpatina" de Cannon se le identificó años más tarde como la noradrenalina, que es la adrenalina sin un grupo metilo. Así se desarrollaron los criterios para identificar el fenómeno de la transmisión neurohumoral:
I) La demostración de la presencia de una sustancia biológicamente activa y de las enzimas necesarias para su elaboración (síntesis) en el sitio analizado.
2) La recuperación de la sustancia a partir de perfusados (baños en donde se mantiene el tejido) de estructuras que reciben algún tipo de nervio después de periodos de estimulación de los nervios de la misma y, a la inversa, de su ausencia cuando no se estimula.
3) La demostración de que el compuesto, administrado exógenamente, reproduce los mismos efectos de la estimulación eléctrica.
4) La demostración de que tanto la respuesta a la estimulación eléctrica como aquella producida por la administración de la sustancia se afectan de la misma manera ante un mismo fármaco.
Por largo tiempo se pensó que cada nervio liberaba un solo tipo de neurotransmisor, pero se ha visto que una misma neurona puede liberar varios tipos de sustancias neuroactivas. Neuronas localizadas en varias partes del cerebro son capaces de liberar neurotransmisores como la acetilcolina y también otros de carácter peptídico (es decir, formados por aminoácidos).
Veamos un poco los pasos que participan en la neurotransmisión neurohumoral, pues éstos resultan de particular importancia en farmacología. Muchos fármacos deben sus efectos a acciones sobre etapas particulares de la neurotransmisión.
Para que se efectúe la neurotransmisión es necesario que haya conducción del impulso nervioso. Hablamos de conducción cuando nos referimos al paso del impulso nervioso a través de un axón o de una fibra muscular. En contraste, el término transmisión significa el paso del impulso nervioso a través de una sinapsis o de una unión neuroefectora, o sea, cuando una célula diferente a la que conduce o produce la impulsión nerviosa es activada (así sea para inhibirse).
Mucho de lo que sabemos actualmente sobre la transmisión del impulso nervioso se lo debemos a los trabajos de Hodgkin y Huxley, en Inglaterra. Estos investigadores, trabajando justo antes y después de la Segunda guerra mundial, aprovecharon el calamar para sus experimentos. La gran ventaja que este animal representa es que posee algunas neuronas de gran tamaño que tienen axones visibles a simple vista (esto significa cientos de veces el tamaño de un axón de mamífero). Fue en este axón gigante de calamar donde el grupo inglés realizó sus estudios.
Como habíamos dicho antes, el interior de una neurona (incluyendo su axón) tiene menos cargas positivas que el exterior, produciéndose una diferencia de voltaje o potencial eléctrico en ambos lados de la membrana neuronal (aproximadamente -70 milésimas de voltio). Esto es, se encuentra polarizada, siendo el polo negativo el interior de la célula y el positivo el exterior. Este potencial de reposo se debe a que existen casi 40 veces más iones del potasio al interior de la célula en relación con el exterior, además de que la membrana neuronal es altamente permeable a este ion. Por otra parte, el sodio (Na+) y el cloro (Cl-) tienen altas concentraciones en el medio extracelular, pero la membrana es mucho menos permeable a estos iones que al potasio (K+). Estas diferencias (gradientes) de concentración a uno y otro lado de la membrana son mantenidas por la presencia de bombas (para meter o sacar iones) asociadas a la membrana. Son sistemas enzimáticos asociados a sustancias productoras de energía (la trifosfatasa que produce adenosín trifosfato (ATP), compuesto de donde se obtiene gran parte de la energía que necesita la célula para todo tipo de funciones), que transportan en forma activa las partículas con carga eléctrica.
Cuando una fibra nerviosa es estimulada (despolarizada) se inicia un impulso nervioso o potencial de acción. Éste tiene dos fases: una fase inicial producida por la entrada rápida de iones de sodio al interior de la célula, a través de canales de la membrana del axón que son sensibles al voltaje de la misma. La rápida entrada de estas cargas positivas hacen que el valor negativo del interior de la célula en la región estimulada, disminuya rápidamente hacia la positividad. La segunda fase del potencial de acción ocurre por la apertura retardada de canales potásicos que hacen que este ion salga de la célula (recordemos que normalmente los iones de K+ están mucho más concentrados al interior que al exterior), contribuyendo así a una mayor despolarización (la falta de polarización significa la ausencia de una diferencia de potencial, o sea, un valor cercano o igual a cero voltios) de la membrana, pero también a una inactivación de los canales de sodio. Este último fenómeno ya anuncia la repolarización membranal.
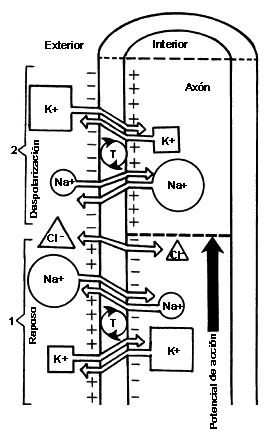
FIGURA IV.I. El potencial de acción: bases
iónicas. Aquí se muestra el corte de una fibra nerviosa donde se ilustran las
diferencias de concentración de iones dentro y fuera del citoplasma: en estado
de reposo (I) el sodio (Na+) tiene mayor concentración
fuera de la célula, mientras que el potasio (K+) es más abundante
en el interior de la célula. Estas diferencias de concentración iónica producen
un desequilibrio eléctrico: el interior de la célula es más negativo que el
exterior. Esta electronegatividad, causada por la presencia de canales iónicos
y bombas o transportadores (T) que mueven los iones de un lado a otro de la
membrana, hace que la célula sea excitable. Cuando llega el impulso nervioso
(cabeza de flecha en la porción media de la figura), la polaridad se invierte
pues el Na+ entra rápidamente a la célula, al tiempo que sale el
K+, produciendo una despolarización: la carga de la membrana pasa
de negativa a positiva (2). La repolarización (el retorno al estado
de excitabilidad previo o de reposo) se logra cuando las bombas membranales
(T), las cuales funcionan por la energía proveniente de la conversión de ATP
en ADP, sacan el Na+ y vuelven a introducir el K+
(de nuevo, al estadio ilustrado en I). Éste es el proceso participante
en la excitación. En la inhibición el ion cloro (Cl-) desempeña un
papel importante, aumentando su concentración intracelular.
El movimiento de corriente eléctrica alrededor del sitio despolarizado hace que los canales iónicos situados en la vecindad también se activen, produciendo una cascada de excitación membranal, y de esta manera, la propagación del impulso nervioso a todo lo largo de la fibra.
Éste es el mecanismo básico por el que un potencial de acción se produce, el fonema fundamental del cerebro, la letra mayúscula del lenguaje neuronal. Cualquier sustancia que afecte estos procesos puede ser mortal. Existen venenos que deben su acción mortífera justamente a sus acciones sobre estas etapas de la producción del impulso nervioso. La tetrodotoxina, extraída de la glándula del pez globo, y la saxitoxina, proveniente de la almeja, bloquean la primera fase del potencial de acción (el aumento de la permeabilidad al Na+). La batracotoxina, producida por glándulas de la piel de una rana sudamericana, produce parálisis por su efecto sobre estos mismos canales, aunque esta vez para activarlos en forma sostenida. Otros venenos de serpiente o alacrán actúan sobre los mismos mecanismos iónicos (es decir, activación, inactivación y sus correspondientes: inactivación de la activación o activación de la inactivación).
Ya sea a una glándula, una fibra muscular o una sinapsis, la llegada del impulso nervioso produce una serie de eventos pre, trans y postsinápticos sensibles a la acción farmacológica. Veamos qué sucede en el interior de cada uno de estos compartimientos, para luego examinar sus interacciones.
Al compartimiento presináptico llega el potencial de acción y allí se produce la conversión de la señal eléctrica en señal química, la cual vuelve después a recuperar sus propiedades eléctricas. Es aquí donde, dependiendo del tipo de neurona, las moléculas del neurotransmisor se elaboran, o si lo hacen en el cuerpo neuronal, maduran para su liberación hacia la hendidura sináptica. En este último caso se trata, generalmente, de péptidos que se sintetizan en el soma y que son transportados por el flujo axonal (el movimiento de sustancias a través del axón) anterógrado (hacia la periferia) hasta la terminal sináptica. Las sustancias que se transportan hacia el soma neuronal lo hacen por flujo axonal retrógrado.
El neurotransmisor puede almacenarse en vesículas sinápticas, pequeños reservorios globulares que contienen receptores en su pared exterior y permiten que el neurotransmisor se libere en sitios específicos de la terminal presináptica. Se ha hecho la analogía de la terminal presináptica como un espacio donde las vesículas sinápticas, así como las mitocondrias y otras estructuras subcelulares, están flotando. Sólo en un lugar determinado de este espacio se localizan los sitios por donde el neurotransmisor puede liberarse hacia el exterior. Como si las vesículas fueran huevos que sólo pudieran acomodarse en los huecos de sus cajas, que se encuentran en el piso de este espacio. Y sólo a través de los huecos de estas cajas se puede descargar el contenido hacia el exterior. En este caso, los huecos tienen receptores que reconocen los componentes de la cáscara del huevo.
La presencia de estas vesículas y de receptores en sus membranas, y de moléculas
de el neurotransmisor significa que existe todo lo necesario en el interior
de la terminal para sintetizar todos estos componentes, y de mecanismos para
la regulación de esta síntesis y de la liberación sináptica. La terminal presináptica
tiene autorreceptores que le informan sobre los niveles del neurotransmisor
en el exterior de la terminal. Si sus niveles son elevados, la terminal puede
fabricar o liberar menos. Si éstos son bajos, puede hacer lo contrario. La activación,
pues, de los autorreceptores puede tener efectos de estimulación o, generalmente,
de inhibición de la liberación sináptica. La membrana de la terminal presináptica
también posee moléculas transportadoras del mismo neurotransmisor que ellas
liberan. Es un mecanismo de recaptación que sirve no sólo para ahorrar neurotransmisor
al reutilizarlo, sino también contribuye a limitar sus efectos postsinápticos.
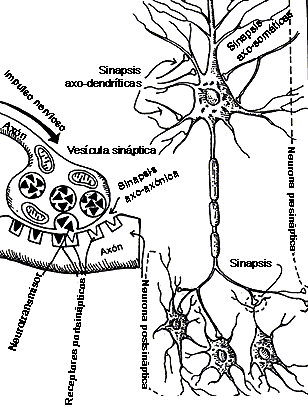
FIGURA IV.2. La sinapsis. Es el sitio donde
una célula nerviosa se comunica con otra. Aquí se ilustran los sitios posibles
de contacto: en el cuerpo celular (sinapsis axosomáticas), en las dendritas
(sinapsis axodendríticas) o en el axón mismo, como en las sinapsis axoaxónicas
(figura de la izquierda). El impulso nervioso, al llegar a la sinapsis, provoca
la liberación del neurotransmisor a partir de vesículas sinápticas, que actúa
en los receptores postsinápticos.
Cuando el potencial de acción llega a la sinapsis, se produce la entrada del ion calcio (Ca2+), que hace que las vesículas se fusionen con la membrana celular y liberen su contenido al exterior. Este proceso se conoce como exocitosis. Normalmente hay vesículas que están liberando neurotransmisor todo el tiempo, produciendo los llamados potenciales miniatura en la postsinapsis. Lo que hace el potencial de acción, mediante o ayudado por el aumento de calcio intracelular, es provocar la liberación del contenido de cientos de vesículas al mismo tiempo.
Junto con el neurotransmisor se liberan otras sustancias proteicas que también contribuyen a los efectos postsinápticos, quizá con una acción cuya duración es mucho más prolongada que la del neurotransmisor mismo, probablemente con efectos tróficos sobre otras células (los efectos tróficos son aquellos que favorecen la sobrevivencia, la diferenciación y el crecimiento celular).
Como en el caso del potencial de acción, existen fármacos cuyos efectos se deben a estos mecanismos. Así, todas las sustancias que interfieran con la entrada de calcio a la terminal presináptica (porque actúan sobre los canales iónicos de este ion) se comportarán como antagonistas de la liberación del neurotransmisor. Todas aquellas drogas que estimulen autorreceptores inhibitorios harán lo mismo. Y a la inversa, los fármacos que aumenten la entrada de calcio o inhiban los autorreceptores, producirán una estimulación de la liberación del neurotransmisor.
EL COMPARTIMIENTO POSTSINÁPTICO
Cuando el neurotransmisor liberado por la presinapsis alcanza la membrana postsináptica
se combina con receptores específicos allí localizados. Entonces pueden suceder
tres cosas: a) aumentar la permeabilidad a cationes (usualmente el Na+,
a veces el Ca2+), lo que produce una despolarización, llamado
potencial postsináptico excitador (PPSE) o, en el caso del
músculo esquelético, potencial de placa motriz; b) aumentar la permeabilidad
membranal a aniones (moléculas cargadas negativamente, como el cloro), lo que
producirá una estabilización del potencial de membrana o incluso una hiperpolarización,
es decir, un potencial postsináptico inhibidor (PPSI). En
otras palabras, el potencial de reposo conservara sus valores normalmente negativos
o incluso los aumentará (véase la figura IV.I.);
y c) aumentar selectivamente la permeabilidad a iones de K+.
Este aumento de la permeabilidad provoca que el K+ salga de
la célula (pues es allí donde se encuentra más concentrado), lo que conduce
a una hiperpolarización o estabilización de la membrana, o sea, a un PPSI.
De esta manera, un neurotransmisor puede excitar la membrana postsináptica (generando
un PPSE) o inhibiría (con un PPSI).
Mucho de lo que conocemos sobre la sinapsis lo hemos averiguado gracias al estudio de la unión neuroneuromuscular, llamada también placa motriz, esto es, el sitio donde el nervio hace contacto con el músculo; el neurotransmisor que allí se libera es la acetilcolina. Sabemos que justo en este sitio existe una acumulación de receptores de esta sustancia. La ocupación del receptor muscular da lugar a la contracción de las fibras musculares. El bloqueo del mismo produce parálisis. En la actualidad sabemos que el nervio motor no sólo produce contracción muscular, sino que también provee al músculo de factores necesarios para su conservación (factores tróficos). En ausencia de ambos el músculo se atrofia (las fibras musculares van muriendo) en forma irreversible.
Como vemos, los efectos netos producidos por un neurotransmisor se deben a flujos iónicos pasivos, de acuerdo con los gradientes de concentración de los mismos. Los cambios de la permeabilidad de los canales a través de los cuales estos iones se desplazan están regulados por receptores especializados que se localizan en la membrana postsináptica. Existen muchos tipos de receptores. Los que hemos mencionado hasta ahora se conocen como receptores ionotrópicos, por su afinidad o relación con iones. Existen otros receptores llamados metabotrópicos, por tener relación con el metabolismo celular, es decir, con moléculas presentes en el interior del compartimiento postsináptico encargados de funciones intracelulares (versus las puramente membranales de los ionotrópicos). Frecuentemente, la ocupación de estos receptores da lugar a movilización del calcio, el cual activará diversas enzimas. Esto lo detallaremos más adelante.
Cuando decimos receptor nos referimos a proteínas. Se trata de moléculas compuestas por cadenas de aminoácidos que forman enlaces entre sí. Al establecer estos enlaces se crean cavidades y protuberancias, que pueden constituir receptores para sustancias que tengan la forma correspondiente (como un listón que se dobla sobre sí mismo varias veces). Es como si el neurotransmisor fuera una llave que pudiera abrir o cerrar una cerradura, y ésta sería una proteína que acepta sólo un tipo de llave. Si la llave adecuada entra bien en la cerradura, ésta podrá activarla. Si esta llave sólo entra un poco, impedirá que otra llave entre y, por tanto, actuará como un antagonista.
Recientemente se ha observado que los receptores ionotrópicos se organizan
en familias de proteínas con semejanzas estructurales y pequeñas diferencias
que les permiten interactuar con diferentes transmisores. En esta familia encontramos
los receptores a la acetilcolina, al g- aminobutirato
(GABA), a la glicina y a otros aminoácidos que veremos en detalle
más adelante.
El neurotransmisor termina su efecto cuando su concentración disminuye. Esto puede lograrse por la recaptación del neurotransmisor en la terminal presináptica, como habíamos visto, por ataque enzimático a nivel de la hendidura sináptica o por su captación por las células gliales, que consideraremos como parte del tercer compartimiento sináptico: el transináptico.
EL COMPARTIMIENTO TRANSINÁPTICO
Consideramos este compartimiento como el espacio formado por la glía y el medio extracelular. El espacio extracelular contiene, además de los iones que mencionamos, otras sustancias, como hormonas, factores tróficos, péptidos, etcétera.
En sus membranas la glía contiene receptores para todas estas sustancias, así como transportadores que pueden captarlas activamente hacia el interior de la célula, donde serán metabolizadas. Además, la glía secreta sustancias que permiten a las neuronas crecer y extender sus terminaciones (factores tróficos y trópicos, respectivamente) hacia el espacio extracelular. La glía se encarga también de formar cicatrices, en casos de lesión y desempeña un papel importante en funciones inmunológicas en el interior del sistema nervioso. Finalmente, la glía contribuye con la función de barrera hematoencefálica. En la actualidad conocemos pocas drogas que actúen específicamente con las células gliales. Sabemos, por el contrario, que existen enfermedades que afectan exclusivamente a esta población celular, y son tan graves como las correspondientes a las neuronas (la esclerosis múltiple es una de ellas). El día que descubramos sustancias que sean específicas para este tipo de células, podremos actuar contra padecimientos como los tumores gliales o interferir con la cicatrización anormal que puede ocurrir en casos de traumatismos al sistema nervioso.

![[Anteior]](../img/prevsec.gif)
![[Previo]](../img/parntsec.gif)
![[Siguiente]](../img/nextsec.gif)