V. LOS NEUROTRANSMISORES
DEFINIMOS A UN NEUROTRANSMISOR como una sustancia producida por una célula nerviosa capaz de alterar el funcionamiento de otra célula de manera breve o durable, por medio de la ocupación de receptores específicos y por la activación de mecanismos iónicos y/o metabólicos.
Aquí tenemos que imaginar las posibilidades de un neurotransmisor. La sustancia
es capaz de estimular o inhibir rápida o lentamente (desde milésimas de segundo
hasta horas o días), puede liberarse hacia la sangre (en lugar de hacia otra
neurona, glándula o músculo) para actuar sobre varias células y a distancia
del sitio de liberación (como una hormona), puede permitir, facilitar
o antagonizar los efectos de otros neurotransmisores. O también puede activar
otras sustancias del interior de la célula (los llamados segundos mensajeros,
véase la figura VII.2, p. 113)
para producir efectos biológicos (p. ejem., activar enzimas como las fosforilasas
o las cinasas). Y además, una misma neurona puede tener efectos diferentes sobre
las estructuras postsinápticas, dependiendo del tipo de receptor postsináptico
presente (p. ejem., excitar en un sitio, inhibir en otro e inducir la secreción
de una neurona en un tercero).
Para todas estas posibilidades se han usado términos como el de neuromodulador, neurorregulador, neurohormona o neuromediador. Aunque el uso de términos diferentes puede ayudar a definir acciones y contextos de comunicación intercelular, aquí utilizaremos el de neurotransmisor, pues hablamos simplemente de intercambio de información, de transmisión de señales, de uniones funcionales entre células.
¿Cómo se reconoce a un neurotransmisor? ¿Cómo saber que una neurona produce una sustancia que afecta a otra? Los criterios para identificar a una sustancia como neurotransmisor son semejantes a los que mencionamos cuando hablamos del sistema nervioso autónomo (básicamente, la acetilcolina y adrenalina). Antes de tratar estos puntos es necesario decir que las técnicas de las que actualmente disponemos para abordar estos problemas siguen siendo relativamente burdas. Querríamos saber lo que sucede en una fracción de la célula que mide millonésimas de milímetro y los instrumentos que tenemos sólo detectan porciones más grandes. A medida que estos instrumentos se acercan más a lo pequeño, requieren que el objeto a examinar sea lo más puro (o concentrado) posible, si es que queremos distinguirlo de otros objetos (o moléculas) igualmente diminutos. Ya sea con electrodos pequeñísimos o con procedimientos de purificación y enriquecimiento de "jugos" cerebrales, estamos alterando la forma y funciones originales. A pesar de estas limitaciones, ha sido posible averiguar muchos elementos de la función sináptica. Se han utilizado técnicas de citoquímica y de fraccionamiento subcelular con bastante éxito, las cuales han permitido aislar estos componentes y así estudiarlos. Recordemos nuevamente los criterios:
a) Se debe demostrar la presencia del transmisor en las terminales presinápticas y en las neuronas de donde estas terminales provienen.
Hay varios implícitos en estas condiciones. Veamos algunos de ellos.
Si decimos que una sustancia debe estar presente en algún sitio, significa que su distribución y concentración son particulares. Varios transmisores fueron descubiertos al detectarlos en el tejido nervioso en concentraciones particularmente elevadas. Se pensó que si se concentraban tanto en un solo sitio, algo tendrían que ver con la función de tal sitio. Si decimos que allí se localiza y concentra, entonces pensaríamos que la sustancia en cuestión también debería producirse (sintetizarse) en la neurona que suponemos libera ese transmisor. Esto quiere decir que también tendríamos que identificar los componentes celulares necesarios para su fabricación (enzimas, precursores, metabolitos, etc.), para su transporte (si es que se producen en el soma neuronal para ser liberados a nivel de las terminales) y para su procesamiento una vez liberados (en este caso, la recaptura del neurotransmisor, que constituye uno de los mecanismos de inactivación).
Finalmente, si seccionamos o lesionamos una vía o núcleo neuronal, esperamos que el transmisor en cuestión desaparezca del sitio donde se encuentran sus terminales.
b) El transmisor debe liberarse de la terminal presináptica por estimulación nerviosa. Aquí se incluyen los procesos necesarios para esta liberación, como la existencia de transportadores del transmisor desde el citoplasma al sitio de liberación, lo cual implica a moléculas que interactúan con el esqueleto celular (una red de estructuras que dirige el tránsito de sustancias al interior de la célula) y otras que permiten que la membrana celular pueda abrirse para expulsar el neurotransmisor: Sabemos que para que estos procesos se realicen es necesario el calcio, y por lo mismo, los canales iónicos por los cuales este ion penetra a la terminal.
c) Identidad de acción. Ésta ha sido considerada el criterio principal para tratar a una sustancia como neurotransmisor. Podemos enunciarlo de otra manera: los efectos de la sustancia en cuestión, cuando ésta se aplica al sitio de estudio, deben ser idénticos a aquellos producidos por la estimulación de la terminal presináptica.
El investigador pretende demostrar que la sustancia propuesta como transmisor produzca los mismos cambios iónicos que la estimulación sináptica directa; (sea con electricidad o químicos). Sin embargo, este tipo de estudios requiere registrar intracelularmente la terminal o neurona postsináptica por largo tiempo e, idealmente, contar con una sustancia que antagonice específicamente al transmisor natural. Si el antagonista bloquea los efectos tanto de la estimulación eléctrica como los de la sustancia en cuestión, a dosis semejantes, entonces podremos decir que existe identidad de acción.
Al aplicar diversas técnicas de marcado neuroquímico se han identificado vías nerviosas que tienen neurotransmisores específicos, a partir de neuronas que los sintetizan y envían sus proyecciones hacia lugares distantes del sistema nervioso. Gracias a técnicas que utilizan anticuerpos dirigidos a las enzimas de síntesis de los diversos neurotransmisores, los cuales se conjugan con otras sustancias que fluorescen o que se colorean, se ha podido determinar el curso de las fibras de estas neuronas. En la figura V.I. se ilustran algunas de ellas.
Estudios recientes indican que una misma terminal puede contener varios tipos de transmisores, los cuales pueden liberarse juntos o independientemente.
Un ejemplo de identificación de un transmisor lo constituye la acetilcolina en la unión nervio-músculo. La estimulación del nervio motor libera acetilcolina; existen los mecanismos para su síntesis, almacenamiento y recaptura a nivel de la terminal; el músculo contiene receptores específicos para la acetilcolina; los efectos de la estimulación nerviosa o los de la aplicación externa de esta sustancia dan lugar a los mismos cambios iónicos a nivel de la fibra muscular (la postsinapsis, en este caso) y existen las enzimas necesarias para su metabolismo. En el caso del SNC el problema se complica por la gran densidad de neuronas y sobre todo de terminales, además del factor que mencionábamos de coexistencia de varios transmisores en la misma terminal. A pesar de estas limitaciones, existen pruebas acerca de varias sustancias que permiten que las consideremos como neurotransmisores centrales. Hagamos una revisión de las principales.
Ya vimos el papel que tuvo la acetilcolina en la transmisión neurohumoral (los
experimentos de Loewi en los años 20). A pesar de que conocemos esta sustancia
desde hace mucho tiempo, no se le ha podido investigar en detalle, a nivel central,
por falta de técnicas adecuadas. Está bien establecido que la acetilcolina es
el transmisor a nivel de la unión neuromuscular y en muchas áreas del SNA.
La distribución y concentración de la acetilcolina en el SNC hizo
pensar que también allí podría tener una función. Y finalmente, el efecto neurológico
de varias drogas que se sabía interactuaban con la acetilcolina sugería que
se podría tratar de un transmisor. En los años 50, John Eccles y sus colaboradores
demostraron que un tipo particular de neurona de la médula espinal, la célula
de Renshaw, era sensible a antagonistas colinérgicos y a la misma acetilcolina.
La célula de Renshaw es inervada por motoneuronas (las neuronas que excitan
fibras musculares), así sabemos que en esa sinapsis se libera acetilcolina (recordando
el llamado Principio de Dale, que postula que una neurona libera el mismo
neurotransmisor en todas sus terminales). Hasta ahora es de los pocos ejemplos
de sinapsis centrales donde se reúnen casi todos los criterios necesarios para
considerar a esta sustancia como neurotransmisor, a pesar de que se ha demostrado
que la acetilcolina se encuentra en otras áreas del SNC y que muchas
regiones cerebrales son sensibles a esta sustancia.
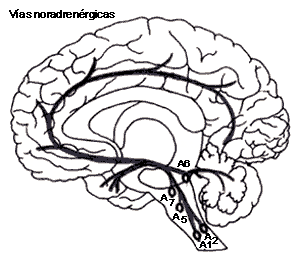
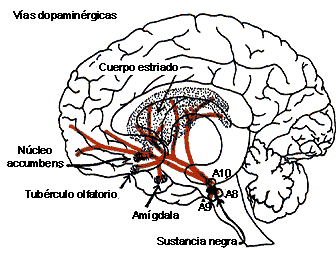
FIGURA V.IA. Las vías de los neurotransmisores:
noradrenérgicos (en negro) y dopaminérgicos (en amarillo). Las letras A indican
la localización de grupos neuronales que sintetizan el neurotransmisor.
La acetilcolina se elabora a partir de la colina, cuyo origen en general es
la dieta, y de la acetil-coenzima A, que proviene de la glucosa a través de
varios pasos metabólicos que ocurren en las mitocondrias. La enzima que une
estas dos moléculas para producir acetilcolina es la colina-acetiltransferasa
(véase la figura V. 2.). Las enzimas que
destruyen a la acetilcolina se llaman acetilcolinesterasas. Se ha visto que
existen varias colinesterasas, y que diversas áreas cerebrales pueden contener
niveles diferentes de ellas.
La primera vía colinérgica demostrada a nivel del SNC fue la que
se forma con fibras colaterales del axón de las motoneuronas espinales (llamadas
colaterales recurrentes, porque salen del mismo axón y retornan en dirección
al cuerpo neuronal) hacia la célula de Renshaw. Esta célula, al activarse por
estas recurrentes, inhibe a la motoneurona, constituyendo así un circuito de
retroalimentación negativa. A niveles superiores ha sido más difícil hacer los
mapas de las vías colinérgicas, por la ausencia de marcadores de las mismas.
Hace años se utilizaron técnicas histoquímicas para hacer que la acetilcolinesterasa
reaccionara con ciertos colorantes, y así señalar su presencia. Actualmente
se utilizan anticuerpos contra la enzima que participa en la síntesis de la
acetilcolina, la colina-acetiltransferasa, asociados a otras moléculas que pueden
marcarse para ser vistas por microscopía. La inervación colinérgica central
se distribuye ampliamente, sea por medio de interneuronas (neuronas contenidas
en un núcleo, y que no envían sus prolongaciones axónicas fuera de él) o por
vías largas que se ramifican. Entre estas últimas están las fibras que nacen
del núcleo basal de Meynert (llamado magnocelular en la rata), localizado a
lo largo de la porción basal del cerebro anterior y que, por una parte, envía
prolongaciones a la corteza cerebral en forma difusa y, por la otra, a grupos
de neuronas situados en el tallo cerebral que se proyectan hacia estructuras
como el tálamo, la formación reticular y los núcleos cerebelosos y vestibulares,
además de hacia varios nervios craneales (como el vago, del que ya hablamos).
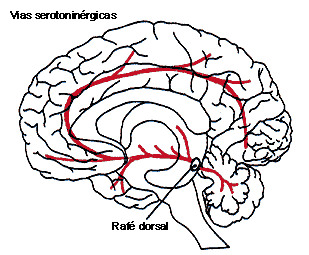
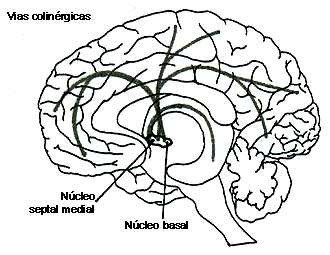
FIGURAV.IB. Las vías de neurotransmisores:
serotoninérgicos (en rojo) y colinérgicos (en verde). Se indican los principales
núcleos de origen de dichas vías.
Existen neuronas que responden a la acetilcolina en muchas partes del cerebro, y de acuerdo con la región que se estudie, este neurotransmisor puede tener efectos excitadores o inhibidores. Los receptores colinérgicos han sido divididos en dos tipos: los muscarínicos y los nicotínicos. Estos términos se refieren a los efectos de la muscarina, sustancia proveniente de un hongo (Amanita muscaria) que tiene efectos similares a los de la nicotina, contenida en el tabaco, y de la acetilcolina. La muscarina, en general, estimula los receptores colinérgicos, mientras que la nicotina primero los estimula y después los bloquea.
Como vimos, existen dos familias de receptores colinérgicos: los muscarínicos y los nicotínicos. Hasta la fecha, se han descrito cinco subtipos de receptores muscarínicos (que llamaremos M1 a M5). La ocupación de todos ellos produce respuestas relativamente lentas (de 100 a 250 milisegundos de duración), mediadas directamente por receptores ionotrópicos (canales de K+, Ca2+ o Cl-) o por segundos mensajeros (la familia de proteínas G). Dependiendo del tipo celular participante, se obtendrán respuestas excitatorias o inhibitorias. La identificación de estos subtipos de receptor ha sido posible gracias a que se cuenta con antagonistas de algunos de ellos, y a técnicas de biología molecular por medio de las cuales se han aislado cadenas de aminoácidos particulares de cada subtipo.
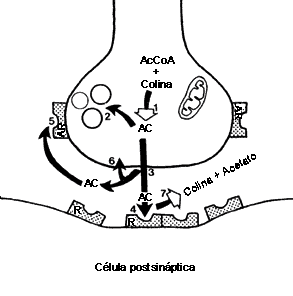
FIGURA V. 2. La sinapsis colinérgica. Esquema
de una sinapsis que sintetiza, acumula y libera acetilcolina. El neurotransmisor
proviene de la conversión del aminoácido precursor: la colina, junto con la
acetil-coenzima A (AcCoA), a través de la enzima colina-acetilasa (I),
hacia acetilcolina (AC). Esta puede almacenarse en vesículas (2)
o liberarse directamente (3). Una vez fuera de la terminal sináptica,
la acetilcolina puede ocupar sitios receptores (R) en otra célula
(4), en ella misma —autorreceptores, AR—(5),
recaptarse (6) o metabolizarse —por colinesterasas—(7)
hacia colina y acetato.
Habíamos mencionado la utilidad que tuvo el axón gigante de calamar en los estudios sobre la generación y la conducción del impulso nervioso. En el campo de la transmisión colinérgica se tuvo la suerte de contar con otra preparación marina para investigar, esta vez, el receptor colinérgico. Se trata de la llamada electroplaca. Se encuentra en peces y anguilas eléctricas y corresponde a la placa neuromuscular de los vertebrados. En este caso, el nervio motor también libera acetilcolina, la cual interactúa con su receptor. Sólo que en lugar de producirse una contracción muscular, hay una descarga eléctrica. En estos animales las descargas sirven no sólo para defenderse o atacar, sino también para comunicarse y para marcar territorios. La gran ventaja que ofrecen estas electroplacas es su abundancia de receptores colinérgicos. Y gracias a esta abundancia se pudieron obtener preparaciones ricas en receptor en las cuales estudiar su estructura y función. A partir de experimentos bioquímicos y electrofisiológicos se ha visto que la acetilcolina puede liberarse no sólo a partir de vesículas sinápticas, sino también directamente del citoplasma de la terminal. El grupo de Y. Israel, en Francia, ha descrito la presencia de una proteína citoplásmica que se encarga de liberar la acetilcolina al medio extracelular. La han llamado mediatóforo.
En cuanto al receptor nicotínico, aislado del órgano eléctrico, se ha podido marcar y aislar por el descubrimiento de sustancias contenidas en venenos de cobras de la India (bungarotoxina). Gracias a ellos, y a técnicas inmunológicas, se han identificado al menos cuatro subtipos de receptor, de acuerdo con su localización en el organismo.
Mecanismos y funciones colinérgicas
Se ha relacionado a la acetilcolina con funciones mnésicas (las ligadas a la memoria), así como en la transmisión del dolor, el calor y los sabores. También en la regulación de los movimientos voluntarios y el control del ciclo sueño-vigilia. Muchas de las pruebas que originaron estas hipótesis funcionales se obtuvieron por el uso de agonistas (sustancias que imitan el efecto de la sustancia en cuestión) y antagonistas colinérgicos. Por ejemplo, la oxotremorina y la arecolina, agonistas muscarínicos, producen temblor que se bloquea con atropina, antagonista muscarínico por excelencia. La nicotina también produce temblor, pero no puede antagonizarse con atropina. Esto sugiere que ambos tipos de receptor (muscarínico y nicotínico) participan en el temblor. También se ha visto que microinyecciones de agonistas colinérgicos en el tallo cerebral pueden afectar el ciclo sueño-vigilia. De estos aspectos funcionales hablaremos más adelante, cuando tratemos las drogas utilizadas en la enfermedad de Parkinson y de fármacos que inducen sueño (los hipnóticos).
En ciertas enfermedades neurológicas, claramente se han identificado anticuerpos contra el receptor colinérgico del músculo esquelético, como en casos de pacientes con miastenia gravis. En otros padecimientos cerebrales, el papel de la acetilcolina es menos claro, aunque de acuerdo con los efectos de agonistas y antagonistas, se ha propuesto que participa en padecimientos como la corea de Huntington, y las enfermedades de Parkinson y Alzheimer. De estas alteraciones hablaremos más adelante.
Mencionemos finalmente, que muchos insecticidas y algunos gases utilizados en situaciones de guerra deben sus acciones a los efectos antagónicos irreversibles de la acetilcolinesterasa.
Estás sustancias pertenecen al grupo de las catecolaminas, que también incluyen a la dopamina.
Antes mencionamos que en el SNA han sido utilizados los extractos de glándula suprarrenal para producir respuestas fisiológicas (de allí el término adrenalina). No fue sino hasta 1946 cuando se identificó el verdadero transmisor de los nervios de la división simpática del SNA: la noradrenalina.
A diferencia de la acetilcolina, las catecolaminas muestran una distribución bastante desigual en el sistema nervioso, es decir, hay áreas donde son muy abundantes y en otras son muy escasas.
En los años 60, dos grupos de investigadores suecos descubrieron que si se exponían tejidos nerviosos congelados al vapor de formaldehído caliente (entre 60 y 80°C) se lograba que las catecolaminas emitieran fluorescencia. De esta manera, por primera vez en la historia de la neurotransmisión, se pudo avanzar en la delimitación de vías y en aspectos morfológicos de las neuronas catecolaminérgicas. Se vio que una sola neurona podía tener terminales hasta de 10 a 20 cm de distancia del soma, y que éstas mostraban varicosidades (al microscopio aparecían cómo un rosario) ricas en fluorescencia.
Estudios más detallados mostraron después, que las tres principales catecolaminas (adrenalina, noradrenalina y dopamina) se distribuían en forma diferente en el interior del sistema nervioso, y que en ciertas áreas eran más abundantes unas que otras.
Actualmente conocemos con bastante detalle cómo se forman las catecolaminas
en el interior del cerebro, en las células cromafines (de la glándula suprarrenal)
y en los nervios y ganglios del sistema simpático. A partir del aminoácido l-
tirosina, la enzima tirosina-hidroxilasa (TH) lo convierte
en DOPA (dihidroxifenilalanina) y ésta se transforma, por la DOPA-descarboxilasa,
en dopamina, ésta a su vez puede transformarse, en aquellas células que contengan
la enzima dopamina-b-hidroxilasa (DBH),
en noradrenalina. La noradrenalina puede convertirse en adrenalina por otra
transferencia de metilos, a cargo de la fenil-etanol-amina-N-metiltransferasa
(PNMT). La noradrenalina, a su vez, inhibe a la tirosina-hidroxilasa,
funcionando así como señal de interrupción de la síntesis. A este mecanismo
se le conoce como "inhibición por producto final". Estas vías metabólicas se
ilustran en las figuras V.3 y V.4.
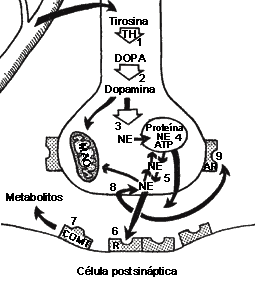
FIGURA V.3. La sinapsis noradrenérgica. Esquema
de una sinapsis que sintetiza, acumula y libera noradrenalina o norepinefrina
(NE). El neurotransmisor proviene de la conversión del aminoácido
precursor, la tirosina, a través de varios pasos enzimáticos, hasta noradrenalina:
la tirosina-hidroxilasa (TH) convierte la tirosina en DOPA
(I); la DOPA- descarboxilasa la convierte en dopamina
(2), y la dopamina -b- hidroxilasa en
noradrenalina (3). Ésta puede almacenarse junto con otras proteínas
sinápticas y con ATP (4) para de allí liberarse, directa
o indirectamente (5). Una vez liberado, el neurotransmisor puede
ocupar receptores postsinápticos (6), metabolizarse por la enzima
catecol -O- metiltransferasa (COMT) (7), recaptarse
(8) para su eventual reutilización u ocupar autorreceptores (AR)
(9).
La tirosina-hidroxilasa parece ser el paso limitante de todas estas reacciones. Es decir, se trata del paso enzimático crítico en la síntesis de las catecolaminas. Si queremos aumentar sus niveles, tendremos que aumentar los volúmenes de la enzima. Estos cambios dependen, por un lado, de la cantidad de enzima y del sustrato (la molécula sobre la cual actúa la enzima), de la disponibilidad de los llamados cofactores (como las vitaminas, que funcionan como "ayudantes" de las enzimas) y, por otro, de la intensidad y patrón de activación nerviosa.
Las catecolaminas se almacenan en vesículas que se transportan desde el cuerpo celular hasta las terminales. La liberación del neurotransmisor parece efectuarse no solamente en éstas, sino también en las varicosidades de las fibras catecolaminérgicas. Muchas de esas varicosidades no están asociadas a terminales postsinápticas, por lo que se ha sugerido que puede haber liberación "extrasináptica" del neurotransmisor. Esta liberación podría contribuir a la existencia de niveles cambiantes de catecolaminas en el espacio extracelular, con posibilidad de inducir efectos a distancia y sobre un gran número de estructuras cerebrales. Es lo que se ha llamado "transmisión de volumen" en el sistema nervioso y, aunque aún está sujeto a controversia, se trata de un interesante concepto que podría tener relación con estados cerebrales "difusos" como el sueño, la vigilia o la atención selectiva.
La liberación de catecolaminas se regula por la existencia de autorreceptores en la terminal presináptica, los cuales responden a la concentración del neurotransmisor en la sinapsis. Los nervios adarenérgicos parecen tener varios tipos de autorreceptores. Unos responden a las mismas catecolaminas, mientras que otros lo hacen a diferentes neurotransmisores. Esto agrega posibilidades de regulación del tránsito sináptico y ofrece también oportunidades de intervención farmacológica.
En lo que respecta al metabolismo de las catecolaminas, tanto la dopamina como la noradrenalina se degradan por la monoaminooxidasa (MAO) y la catecol-O-metiltransferasa (COMT), en ácido homovanílico (HVA) y ácido dihidroxifenilacético (DOPAC). Ambos metabolitos pueden cuantificarse en el líquido cefalorraquídeo u orina para tener un índice de actividad catecolaminérgica.
Quizás el lector se encuentre un poco abrumado con tantos nombres y detalles. Pero si se quiere entender, al menos un poco, lo que algunas drogas hacen en el cerebro, es mejor tener a la mano la información necesaria. Un par de ejemplos: a sujetos con Parkinson, que como veremos tienen deficiencia de dopamina cerebral, se les administra no sólo su precursor (la L-DOPA), sino también inhibidores de la descarboxilasa (la enzima que, como acabamos de ver, la destruye), para aumentar su efecto. Los inhibidores de la MAO se utilizan clínicamente como antidepresivos.
Como consecuencia del desarrollo de agonistas y antagonistas catecolaminérgicos
específicos se ha podido establecer la existencia de varios tipos de receptores
adrenérgicos. Clásicamente se les ha dividido en dos familias: los a
y los b-adrenérgicos. Ahora se sabe que al interior
de estas familias existen otros subtipos. Así, se conocen hasta la fecha tres
tipos de receptores b (I, 2 y 3), cuatro
a- I (A, B, C, y D) y tres a-2
(A, B y C), de acuerdo con sus efectos sobre las llamadas proteínas G, aunque
este número puede aumentar. Las proteínas G representan una familia de segundos
mensajeros (véase la figura VII.2)
que traduce la señal dada por la ocupación del receptor membranal al lenguaje
neuronal intracelular a través de la activación o inhibición enzimática. Así
tendremos proteínas G estimuladoras y proteínas G inhibidoras.
En el SNC, se ha asociado la presencia de receptores a-
I, a-2 y b-I
con neuronas, y los b-2 con la glía
y células vasculares.
Los receptores adrenérgicos están sujetos a varios tipos de regulación. Su número puede aumentar o disminuir, lo mismo que su sensibilidad a fármacos (p. ejem., después del uso crónico).
Se ha podido determinar la distribución de fibras y cuerpos celulares con contenido
catecolaminérgico. Es un sistema de transmisión notable: se origina en áreas
muy circunscritas del tallo cerebral y envía ramificaciones a todas las áreas
del cerebro, cerebelo y médula espinal que se han estudiado. Esta proyección
amplia hace que sus influencias sean generalizadas (véase
la figura V.I).
Las principales fibras noradrenérgicas nacen de dos sitios principales del tallo cerebral: el locus coeruleus y el área tegmental lateral.
El locus coeruleus se encuentra en la porción más inferior del tallo cerebral. En el ser humano está constituido por aproximadamente 12 000 neuronas en cada lado del cerebro. Estas neuronas dan lugar a cinco haces de fibras principales, que llegan al tálamo, hipotálamo, hipocampo, bulbo olfatorio, y muchas otras áreas, para eventualmente terminar en la corteza cerebral.
A partir del momento en que se identificó y aisló la enzima que convierte la noradrenalina en adrenalina (la PNMT), se crearon anticuerpos contra ella para así localizarla en el sistema nervioso. Se encontraron dos grupos principales de neuronas que contienen esta enzima (y que, por tanto, pueden elaborar adrenalina) también a nivel del tallo cerebral inferior y lateral. Estas células están asociadas a centros de regulación autónoma de funciones respiratorias, cardiovasculares y viscerales, por una parte, y por otra, a estructuras hipotalámicas más anteriores.
Hasta hace relativamente poco tiempo, se pensaba que la dopamina era sólo un producto intermedio del metabolismo de las catecolaminas. Sin embargo, al observarse que la distribución cerebral de la dopamina y la noradrenalina eran francamente diferentes y que la primera era mucho más abundante que la segunda, se le empezó a considerar más seriamente como un neurotransmisor aparte. El refinamiento de las técnicas anatómicas (p. ejem., histofluorescencia) mostró que gran parte de la dopamina cerebral se concentraba en los ganglios basales (véase el capítulo II), concluyendo entonces que esta sustancia podría tener algo que ver con el control del movimiento y patologías como la enfermedad de Parkinson.
La dopamina, como el resto de las catecolaminas, se sintetiza a partir de la
l- tirosina, que debe ser transportada hacia el cerebro a través de la barrera
hematoencefálica hasta la neurona dopaminérgica. Allí, la enzima tirosina-hidroxilasa
la transformará en 1-dihidroxifenilalanina (L-DOPA), y la DOPA-descarboxilasa
a dopamina. Si queremos aumentar los niveles cerebrales de dopamina es necesario
aumentar la concentración de DOPA, que normalmente es bastante
baja. No se obtiene tal efecto aumentando los niveles de l- tirosina, que ya
de por sí son relativamente elevados.
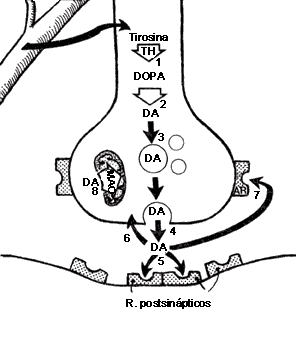
FIGURA V.4. La sinapsis dopaminérgica. La dopamina
(DA) se sintetiza a partir de la tirosina, a través de los mismos
pasos enzimáticos que la sinapsis noradrenérgica: la tirosina-hidroxilasa (TH)
convierte la tirosina en DOPA (I); la DOPA-descarboxilasa
la convierte en dopamina (2). La DA puede almacenarse
(3) para de allí liberarse (4). Una vez liberado el
neurotransmisor puede ocupar receptores postsinápticos (5), metabolizarse,
recaptarse (6) u ocupar autorreceptores (AR) (7).
Dentro de la terminal, la DA puede metabolizarse por la monoamino-oxidasa
mitocondrial (8).
Se han descrito tres sistemas dopaminérgicos principales en el cerebro:
a) El sistema nigro-estriado, donde los cuerpos celulares se hayan localizados en la sustancia nigra y sus axones proyectan hacia el neoestriado (núcleos caudado y putamen). Se considera parte del llamado sistema extrapiramidal.
b) El sistema mesolímbico y mesocortical, que se origina en el área tegmental ventral del mesencéfalo, y envía sus axones hacia estructuras estriatales, límbicas y corticales, y
c) El sistema tuberoinfundibular, con fibras relativamente cortas que nacen en el hipotálamo (núcleo arcuato y periventricular) y terminan en la hipófisis (lóbulo intermedio) y la eminencia media.
Existen también interneuronas dopaminérgicas en la retina, el bulbo olfatorio y el hipotálamo.
Hay varias formas de clasificar a los receptores dopaminérgicos (de hecho a todos los receptores de interés farmacológico). Una de ellas es por su localización: a) receptores de las células no dopaminérgicas: por definición, receptores postsinápticos, y b) receptores de células dopaminérgicas, también llamados autorreceptores, pues responden al mismo neurotransmisor liberado por la neurona.
Los receptores dopaminérgicos postsinápticos se han clasificado en dos grandes
grupos, dependiendo de sus efectos en una enzima llamada adenilato-ciclasa.
Esta enzima es parte de la familia de los segundos mensajeros (véase
la figura VII. 2), y está encargada de aumentar los niveles
intracelulares del adenosín monofosfato cíclico (AMPc),
compuesto que sirve para activar sistemas enzimáticos ligados más directamente
a los efectos biológicos. Así, los receptores D1 estimulan
la adenilato-ciclasa, mientras que los D2 la inhiben. El receptor
D1 es aproximadamente lo veces menos sensible a la dopamina que el
D2, y como veremos después, estas diferencias de sensibilidad se
correlacionan con la potencia de drogas tranquilizantes.
Ambos tipos de receptores han sido encontrados en las áreas de proyección dopaminérgica, aunque es posible que se localicen en células diferentes. En el estriado la activación de los receptores dopaminérgicos disminuye la actividad de la vía estriado-nigral (la que regresa al sitio de origen de la vía dopaminérgica nigroestriada), constituyendo así un sistema de retroalimentación negativa. Es importante mencionar que la exposición prolongada a agonistas o antagonistas dopaminérgicos puede producir cambios importantes en la sensibilidad del receptor.
En relación con los autorreceptores, éstos pueden existir en cualquier nivel de la neurona dopaminérgica para regular su actividad. Así, los localizados en la región somatodendrítica disminuyen la frecuencia de generación de potenciales de acción, mientras que la estimulación de los autorreceptores a nivel de la terminal sináptica inhibe la síntesis y liberación del neurotransmisor. Ambos tipos de autorreceptores son, en su mayoría, del tipo D2 (por tanto, varias veces más sensibles al neurotransmisor que los D1).
Existen diferencias farmacológicas tanto entre los receptores D1 y D2 como entre los autorreceptores.
Mencionemos, para terminar, que técnicas recientes de neurobiología molecular han permitido identificar subtipos de receptor dopaminérgico. Así, se han descrito cuatro subtipos del receptor D2 y dos subtipos del D1. Quizá la lista se extienda en el futuro.
Como veremos después, los receptores dopaminérgicos participan en gran número de efectos farmacológicos, incluyendo los de agentes tranquilizantes, antidepresivos, antiparkinsonianos y estimulantes y en patologías neurológicas y psiquiátricas serias, como la enfermedad de Parkinson ya mencionada, la esquizofrenia y en fenómenos de adicción a drogas.
Desde el punto de vista histórico, la serotonina (cuyo nombre químico es 5- hidroxitriptamina o 5-HT) ha sido el neurotransmisor que más ha influido en el campo de la neuropsiquiatría. La mayoría de los llamados alucinógenos posee efectos serotoninérgicos, además de cierto parecido estructural con la serotonina misma. Cuando se detectó la presencia de la 5-HT en el cerebro aparecieron las teorías que relacionaban a este neurotransmisor con varias formas de enfermedades mentales.
Aunque existe serotonina en todo el cuerpo, ésta no atraviesa la barrera hematoencefálica,
por lo que el cerebro produce la propia. La síntesis depende del aporte de un
aminoácido, el triptófano, proveniente de la dieta (por lo que sus niveles cerebrales
dependen, en parte, de los alimentos). El siguiente paso en la síntesis de serotonina
es la hidroxilación (adición de un grupo OH) del triptófano, para
dar lugar al 5-hidroxitriptófano. La enzima responsable de esta reacción es
la triptófano hidroxilasa. Existe la p-clorofenilalanina que bloquea justamente
a esta enzima, y que ha sido de gran utilidad en investigación. La administración
de esta sustancia puede disminuir el contenido cerebral de serotonina en casi
80%. Estas manipulaciones se han hecho para investigar las funciones en las
que participa este neurotransmisor, y que estudiaremos más adelante.
La serotonina se obtiene por descarboxilación del 5-hidroxitriptófano, reacción que sucede rápidamente, a medida que el precursor inmediato se encuentra disponible.
La serotonina se metaboliza por medio de la monoaminooxidasa (MAO) y el producto detectable de este catabolismo es el ácido 5-hidroxi-indolacético (5HIAA).
Existen varios mecanismos por los cuales se regula la síntesis, liberación y metabolismo de la serotonina, y varios de ellos son sensibles a drogas de uso clínico.
Vías serotoninérgicas centrales
Por medio de técnicas de histofluorescencia (mismas que se utilizan para la
visualización de las catecolaminas) se han podido identificar núcleos serotoninérgicos
en el interior del sistema nervioso (véase la figura V.I).
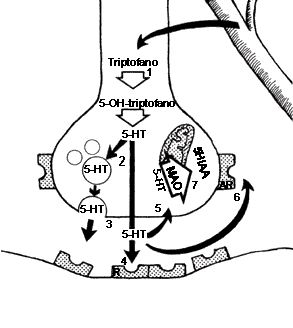
FIGURA V.5. La sinapsis serotoninérgica. La
serotina (5-hidroxitriptamina) (5-HT) se sintetiza a partir del
triptófano, el cual se convierte en 5 - hidroxitriptófano por la enzima triptófano-hidroxilasa.
La 5-HT puede almacenarse en vesículas (2) y/o liberarse
(3). Una vez liberada, puede ocupar receptores postsinápticos (4),
recaptarse (5), ocupar autorreceptores (6) o metabolizarse
por la MAO mitocondrial (7) hacia ácido 5-hidroxiindolacético
(5HIAA).
Sin embargo, cuando se aplican a la 5-HT, la sensibilidad de estas técnicas es mucho menor que con las catecolaminas, por lo que ha resultado difícil el mapeo de las fibras serotoninérgicas. Para ello se ha tenido que tratar previamente a los animales con altas dosis de triptófano, además de con inhibidores de la MAO, buscando aumentar al máximo los niveles de la amina.
Así, se ha observado que las células serotoninérgicas se concentran en la parte media del tallo cerebral, agrupándose en nueve núcleos principales, conocidos como complejo nuclear del rafé. A partir de estos núcleos nacen fibras que llegan a prácticamente todo el sistema nervioso (ganglios basales, hipotálamo, tálamo, hipocampo, sistema límbico, corteza cerebral, cerebelo y médula espinal). Los núcleos más anteriores (en animales) proyectan hacia las partes más rostrales (hacia adelante), mientras que las más posteriores envían sus fibras hacia las áreas del tallo cerebral y la médula. A través de estas proyecciones, la serotonina participa en el control de los estados de sueño y vigilia, el ánimo, las emociones, el control de la temperatura, la dieta, la conducta sexual, algunos tipos de depresión, conducta suicida y ciertos estados alucinatorios inducidos por drogas.
Los receptores serotoninérgicos
La respuesta obtenida después de estimular los núcleos serotoninérgicos puede consistir, por un lado, en una inhibición (hiperpolarización membranal) provocada por aumento de la permeabilidad (conductancia) al K+, o por el otro, en aumento de la frecuencia de disparo (así se le llama al incremento en la ocurrencia de los potenciales de acción) por disminución de la conductancia al K+. A partir de estos estudios fisiológicos y de otros farmacológicos en los que se han empleado diferentes antagonistas, se ha sugerido la existencia de varios subtipos de receptores a la serotonina.
Se han descrito tres tipos principales de receptor: el 5-HT1, el 5-HT2 y el 5-HT3. Y éstos, a su vez, se han subdividido en cuatro subtipos del 5-HT1 (de la A a la D), dos del 5-HT2 (A y B) y, hasta ahora, uno del 5-HT3. De ellos, la mayoría son postsinápticos, pero al menos dos de ellos (el 5-HT1B y el 5-HT1D) pueden ser autorreceptores, modulando la liberación del neurotransmisor. La ocupación de receptores postsinápticos produce sus efectos a través de segundos mensajeros ligados a la fosforilación de moléculas intracelulares y, en algunos casos, por acoplamiento con canales iónicos de calcio.
El sistema nervioso contiene gran cantidad de aminoácidos extremadamente activos para lograr el funcionamiento neuronal. Durante muchos años no se sabía si estas sustancias eran activas en sí o sólo representaban precursores de proteínas (recordemos que todas las proteínas están hechas de aminoácidos). Ahora sabemos que estas pequeñas moléculas son las principales responsables de la conducción nerviosa rápida en el sistema nervioso.
Aminoácidos inhibidores: el GABA y la glicina
El GABA es el neurotransmisor inhibidor predominante del SNC
en su parte supraespinal (grosso modo, la porción intracraneal). En los
años 50 y gracias a técnicas neuroquímicas más sensibles, se observó que el
GABA (g- aminobutirato) no sólo estaba
en el cerebro, sino que además era el órgano que más GABA contenía.
El GABA se forma a partir de otro aminoácido también abundante en el cerebro: el 1- glutamato. Paradójicamente, este precursor es, a su vez, neurotransmisor, pero esta vez excitador.
La enzima que hace esta conversión es la glutamato-descarboxilasa (GAD),
de la que se han encontrado dos formas diferentes; ambas producen GABA
y están codificadas en diferentes genes; sin embargo, no sabemos aún el porqué.
La GAD necesita para su funcionamiento de vitamina B6
(fosfato de piridoxal).
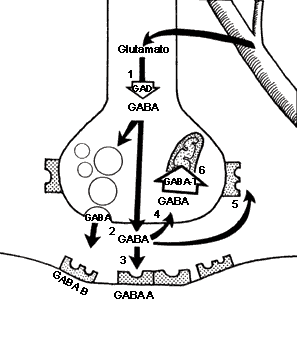
FIGURA V.6. La sinapsis GABAérgica.
El ácido y aminobutirato (GABA) se sintetiza
a partir del glutamato a través de una descarboxilasa (1), la glutamato-des-carboxilasa
(GAD). El GABA puede liberarse hacia el espacio sináptico
directamente o desde almacenes vesiculares (2). Una vez fuera de
la terminal, el GABA puede ocupar receptores postsinápticos (3),
los cuales se han clasificado en tipo A (GABAA) o el
tipo B (GABAB). El aminoácido puede recaptarse (4),
ocupar autorreceptores (AR), que usualmente son tipo B (5),
o metabolizarse por la transaminasa del GABA (GABA-T)
(6).
El GABA ha satisfecho los criterios requeridos para considerarlo como neurotransmisor en la unión neuromuscular de crustáceos, como el acocil (lo cual resulta una buena indicación de que también puede serlo en mamíferos, porque si no ¿dónde quedaría la evolución?) En esta preparación, el GABA produce los mismos efectos que los de la estimulación del nervio correspondiente y la potencia para inducir inhibición producida por extractos de nervio se correlaciona con el contenido de GABA del extracto. Finalmente, tanto el GABA como la estimulación del nervio producen un potencial inhibitorio (una hiperpolarización) por aumento de la conductancia al cloro. Ambos efectos pueden ser bloqueados por el mismo antagonista, la bicuculina.
Las neuronas GABAérgicas muestran una distribución difusa, lo que sugiere que funcionan como interneuronas. Existen, sin embargo, algunas vías GABAérgicas algo más largas como la estriadonigral y la cerebelo-vestibular.
Existen numerosas sustancias que interactúan con receptores GABAérgicos. Todas las que interfieren con su funcionamiento producen aumento de la excitabilidad cerebral hasta el punto de producir crisis convulsivas.
Sustancias que producen sueño (los barbitúricos), o que se utilizan como ansiolíticos
(las benzodiazepinas), actúan en buena parte porque favorecen la transmisión
GABAérgica (véase la figura XVIII.I).
Se han descrito dos tipos de receptor del GABA: el GABAA y el GABAB. El agonista específico para el primero es el muscimol, y el antagonista la bicuculina. Para el receptor GABAB, el agonista específico es el baclofén y el antagonista el faclofén (o el saclofén). Como dijimos, la ocupación del receptor GABAA por un agonista produce aumento de la permeabilidad membranal al cloro. En cambio, la activación del receptor GABAB da lugar a la activación de segundos mensajeros de la familia de las proteínas G.
El otro neurotransmisor inhibidor de importancia, particularmente en el tallo cerebral y la médula espinal, es la glicina. Su efecto es similar al del GABA: hiperpolarización (inhibición) por aumento de la conductancia al cloro. Esta inhibición puede ser antagonizada por la estricnina, otra sustancia convulsivante.
Aminoácidos excitadores: el glutamato y el aspartato
Estas sustancias se encuentran particularmente concentradas en el sistema nervioso, y ejercen potentes efectos excitadores sobre la actividad neuronal. Durante la última década se ha producido muchísima información relativa a la neurobiología de la transmisión glutamatérgica, gracias al desarrollo de sustancias con propiedades agonistas y antagonistas de los diferentes subtipos de receptor del glutamato. Algunas de ellas se han aislado del reino vegetal y muestran potentes efectos despolarizantes: el ácido quiscuálico, obtenido de semillas, el ácido iboténico, aislado de hongos, y el ácido kaínico, proveniente de algas marinas. Este último es cerca de 50 veces más potente que el glutamato mismo, y su inyección intracerebral produce destrucción selectiva de cuerpos neuronales. Este efecto neurotóxico ha sido utilizado a nivel experimental para inducir lesiones en sistemas de los cuales queremos averiguar su función. Al eliminarlos y estudiar los déficits que aparecen, podemos inferir el tipo de función en los que participan. La ventaja de este tipo de lesiones neuroquímicas sobre las eléctricas (en las que se produce una destrucción localizada mediante corriente eléctrica) radica en que las neurotoxinas afectan únicamente los cuerpos celulares del área, ya que los axones y las fibras nerviosas son relativamente resistentes al kainato, mientras que la corriente eléctrica afecta todos los elementos del área en cuestión. Por otra parte, se ha relacionado esta neurotoxicidad, que induce pérdida neuronal, con la patología de la epilepsia y enfermedades cerebrovasculares. En ambos casos, se ha detectado aumento de la concentración extracelular de glutamato después de crisis convulsivas o accidentes isquémicos (por falta de irrigación sanguínea) o hipóxicos (por falta de oxígeno). Parte de las pruebas de estos hallazgos radican en el uso de antagonistas del receptor del glutamato, que han mostrado proteger a las neuronas de este tipo de patologías.
Finalmente, se ha relacionado al glutamato con un tipo de memoria, representado por el fenómeno conocido como potenciación a largo plazo, a nivel de la sinapsis. Todos estos factores han contribuido a estimular la investigación sobre los aminoácidos excitadores.
Hasta la fecha, se han descrito al menos cinco subtipos de receptor del glutamato. Tres de ellos se han definido por los efectos excitatorios (despolarizantes) de agonistas específicos: N-metil-D-aspartato (NMDA) kainato y quiscualato (o AMPA, el nombre de otro agonista más específico) y por los de sus antagonistas específicos. Un cuarto receptor, el del I-2.-amino-4-fosfonobutirato (AP4) que parece representar a un autorreceptor inhibidor. Y un quinto receptor, activado por el ácido transamino-ciclopentano-dicarboxílico (ACPD) y que constituye un receptor metabotrópico, pues tiene efectos sobre el metabolismo de los derivados fosfatados intracelulares.
Las técnicas modernas de neurobiología molecular han permitido obtener información sobre las características fisicoquímicas del receptor así como de sus interacciones con otras sustancias. Se ha visto, por ejemplo, que la glicina, aminoácido con propiedades inhibidoras (como vimos antes), a concentraciones muy bajas, facilita los efectos del NMDA (excitadores), y que drogas como la ketamina (agente anestésico) y la fenciclidina (droga que produce alucinaciones), son antagonistas del receptor al NMDA.
Dada la ubicuidad de los receptores del glutamato, ha resultado difícil establecer con precisión vías nerviosas que utilicen preferentemente a este aminoácido como neurotransmisor; pero existen pruebas de que gran número de fibras cuya estimulación eléctrica produce excitación a nivel de las estructuras a las que proyecta, son de carácter glutamatérgico. El aspartato, otro aminoácido relacionado, podría tener también sus vías específicas, así como efectos particulares y separables de los del glutamato.
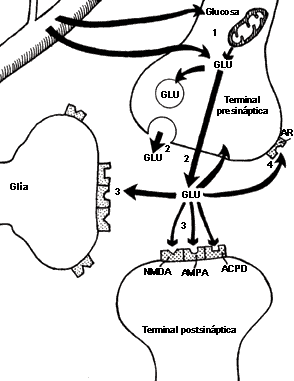
FIGURA V.7. La sinapsis glutamatérgica. El
glutamato (GLU), aminoácido excitador por excelencia, se capta
directamente de la sangre y el espacio extracelular o através de glucosa y la
conversión metabólica en la terminal presináptica (I). Desde allí
puede liberarse directamente o desde almacenes vesiculares (2).
El GLU puede ocupar receptores postsinápticos neuronales o gliales
(3) de tres tipos diferentes, denominados de acuerdo con la sustancia
que interactúa con ellos en forma más específica: los receptores al NMDA
(N-metil-D-aspartato), los no NMDA (sensibles al AMPA)
y los metabotrópicos, sensibles al ácido transamino-ciclopentano-dicarbixílico
(ACPD). El aminoácido también podría interactuar con autorreceptores.
(AR) (4).
Recordemos al lector que un péptido está formado por una cadena de aminoácidos. A su vez, los péptidos forman proteínas. Esta secuencia se controla desde el núcleo de la célula.
Los llamados neuropéptidos constituyen varias familias de moléculas que han mostrado ejercer efectos particulares a nivel del sistema nervioso (aunque muchos de estos péptidos se descubrieron en el intestino). Mencionemos que el sistema gastrointestinal contiene tantas neuronas como el cerebro, las cuales producen los mismos neurotransmisores que las neuronas centrales.
Las neuronas secretoras de péptidos difieren de las productoras de transmisores aminoácidos: estos últimos se forman mediante una o dos reacciones enzimáticas a partir de precursores que provienen, en general, de la dieta. El producto de estas reacciones se almacena en la terminal nerviosa hasta el momento de su liberación. Una vez que ésta ocurre, el transmisor es recaptado por la terminal para ser reutilizado.
A diferencia de estas células, las neuronas liberadoras de péptidos los sintetizan
en el cuerpo celular (en los ribosomas) y siempre a partir de precursores mucho
más grandes. O sea, a partir de moléculas mucho más largas que el neuropéptido.
Estos precursores, o prohormonas, son fraccionados después por enzimas
específicas, en fragmentos más pequeños, algunos de los cuales serán los neuropéptidos
que se liberarán por la terminal (véase la figura
X.I. como un ejemplo). Tanto la prohormona como sus fragmentos
pueden tener efectos biológicos (y, por tanto, receptores) diferentes. Estos
fragmentos se transportan después (por flujo axonal) hasta las terminales, donde
se pueden liberar solos o junto con otro neurotransmisor de tipo aminoácido.
En esta "coliberación" participa el calcio.
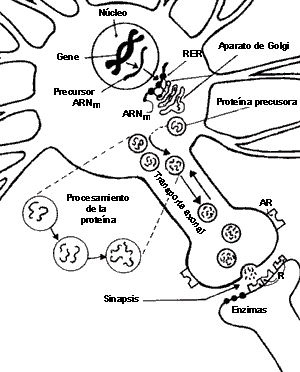
FIGURA V.8. La sinapsis peptidérgica. En éste
esquema se ilustra la síntesis, el procesamiento, el transporte y la secreción
de neuropéptidos. En el núcleo de la célula los genes codifican la formación
de ARNm (ácido ribonucleico mensajero), el cual, en
el retículo endoplásmico rugoso (RER) y el aparato de Golgi, se
convierte en una proteína precursora, que a su vez se procesa en los almacenes
vesiculares al tiempo que se transporta hacia las terminales sinápticas. Este
transporte axonal puede ser hacia delante (anterógrado) o hacia el cuerpo celular
(retrógrado). Finalmente, la neurona libera péptidos (cadenas de aminoácidos)
en la terminal presináptica, que representan fracciones determinadas de la proteína
precursora. Una vez liberados, interactúan con receptores propios o ajenos,
o se degradan enzimáticamente.
Los efectos postsinápticos son, en cuanto a mecanismos íntimos, similares a los de los neurotransmisores "clásicos", esto es, a través de la ocupación de receptores ionotrópicos (canales iónicos) o metabotrópicos (por segundos mensajeros). Algunas asociaciones transmisor-péptido encontradas en el sistema nervioso son las siguientes:
GABA-somatostatina, acetilcolina-colecistoquinina o péptido vasoactivo intestinal (VIP), noradrenalina-somatostatina o sustancia P o encefalina o neuropéptido Y, dopamina-neurotensina o colecistoquinina (CCK), adrenalina-neuropéptido Y o neurotensina, serotonina-sustancia P o encefalina.
Existen varias familias de péptidos, algunos más parecidos entre sí que otros. Unos funcionan por sí solos, mientras que otros modulan el efecto de aminoácidos: el VIP acentúa la respuesta de neuronas corticales a concentraciones muy pequeñas (subóptimas) de noradrenalina.
Encontramos otros neuropéptidos que pueden ser considerados aparte: la colecistoquinina (CCK), somatostatina, angiotensina, péptido relacionado con el gene de la calcitonina (CGRP), factor liberador de la corticotropina, etc. Esta lista seguramente aumentará en los años por venir.
No podemos tratar en detalle cada uno de estos neuropéptidos. Abordaremos en otros capítulos aquellos que se han relacionado con efectos farmacológicos definidos (como los péptidos opioides y los efectos de la morfina). Sólo se enumerarán los miembros más conocidos de cada una de las cinco grandes familias de neuropéptidos considerados hasta ahora:
b) Taquiquininas (que incluye la sustancia P, la kassinina, la eledoisina y la neuroquinina A).
c) Péptidos relacionados con el glucagon (que incluye el VIP,
la secretina, la hormona liberadora de la hormona de crecimiento —GHRH1-24—,
etcétera.)
d) Péptidos relacionados con polipéptidos pancreáticos (que incluye al neuropéptido Y, entre otros).
e) Péptidos opioides (que incluye las prohormonas proopiomelanocortina, la proencefalina, la prodinorfina y sus derivados, como las endorfinas y las encefalinas).
HISTAMINA, PURINAS, PROSTAGLANDINAS
La histamina se ha relacionado clásicamente con los fenómenos alérgicos. A nivel periférico, una reacción alérgica puede producir la aparición de urticaria, comezón, enrojecimiento de la piel, constricción bronquial, etc. Estas reacciones alérgicas pueden ser disminuidas con antihistamínicos, agentes farmacológicos que muestran efectos a nivel del sistema nervioso. Este hecho, aunado al de la imposibilidad para la histamina que circula por la sangre de atravesar la barrera hernatoencefálica, hizo pensar que esta sustancia podría ser un neurotransmisor que se producía en el cerebro. La detección neuroquímica de la síntesis de histamina por las neuronas, junto con la particular distribución de estas neuronas histaminérgicas en el sistema nervioso ha apoyado esta sospecha. La histamina se concentra particularmente en el hipotálamo. De allí, estas células envían sus fibras en forma difusa a todo el sistema nervioso, tal como lo hacen las demás neuronas aminérgicas.
Se han descrito tres tipos principales de receptores a la histamina. El H1, descrito en neuronas, glía y células vasculares, es el más prominente y parece actuar por medio de la movilización de calcio intracelular. Los receptores H2, están relacionados directamente con la adenilato-ciclasa, y los H3, los más sensibles a la histamina, se concentran a nivel de los ganglios basales y el bulbo olfatorio.
En esta familia de moléculas se encuentran los nucleótidos de adenosina. La adenosina ha sido encontrada virtualmente en todas las sinapsis en las que se le ha buscado. Sus principales efectos electrofisiológicos muestran una tendencia a inhibir la liberación de transmisores, pero también se le han descrito efectos postsinápticos, que incluyen desde la interrupción de la liberación en las ardillas, actividad anticonvulsiva (se le ha llamado el anticonvulsivo endógeno), aumento del flujo sanguíneo cerebral e interacciones con el receptor de las benzodiazepinas. Se han localizado al menos dos subtipos de receptor: los P1, que estimulan a la adenilato-ciclasa (y por lo tanto, aumentan la concentración intracelular del AMP cíclico) y que son más sensibles a la adenosina que al ATP. Y los P2, más sensibles al ATP que a la adenosina y cuya activación estimula la producción de prostaglandinas.
A los receptores purinérgicos también se les ha clasificado en relación con los efectos de la adenosina sobre la adenilato-ciclasa en A1, que la estimulan, y en A2, que la inhiben.
Son derivados del ácido araquidónico, consideradas —como las purinas— más como moduladores que como transmisores. Las prostaglandinas, y sustancias relacionadas (prostaciclina y tromboxano), se forman por medio de la ciclooxigenasa, una enzima presente en virtualmente todas las células del organismo (curiosamente, esta enzima es inhibida por la aspirina, y esta inhibición representa parte del efecto terapéutico, analgésico y antinflamatorio de este medicamento).
Existen varias prostaglandinas. En el cerebro se ha demostrado la existencia de prostaglandinas de la serie E y de la F (PGE y PGF, respectivamente), en las que cada serie tiene varios miembros. A nivel del hipotálamo, intervienen en la producción de fiebre inducida por bacterias o toxinas.
Para terminar, mencionemos que pueden existir otras moléculas que contribuyen en la comunicación entre células, ya sea neuronas o glía. La descripción relativamente reciente de los potentes efectos de gases como el óxido nítrico (NO) o el monóxido de carbono (CO) a nivel de la sinapsis hacen pensar que la lista de neurohumores aumentará en el futuro. A estos agentes, junto con el ácido araquidónico, se les ha denominado "terceros mensajeros", pues son capaces de transmitir información en "sentido contrario", es decir, desde la terminal postsináptica a la presinapsis. Son moléculas que seguramente mostrarán su participación en fenómenos nerviosos en un futuro cercano.

![[Anteior]](../img/prevsec.gif)
![[Previo]](../img/parntsec.gif)
![[Siguiente]](../img/nextsec.gif)