I. EL SUPLICIO DE TÁNTALO O LA HEBRA INACCESIBLE
El panorama antes del ADN recombinante
TODOS hemos escuchado hablar del término ingeniería genética.
Los medios de información nos bombardean con descripciones de descubrimientos
asombrosos, y en ocasiones, plantean escenarios de desastre, propiciados por
una maligna intervención del hombre para alterar la naturaleza. En realidad,
si lo meditamos con cuidado, el ser humano siempre ha interferido con los procesos
naturales propios y de otras especies. ¿De dónde proviene, si no, la gran variedad
de razas caninas, por ejemplo? ¿Y no era un serio intrometimiento con el proceso
practicar repetidas sangrías al enfermo a la menor provocación? La administración
de antibióticos para salvar innumerables vidas humanas, ¿no significa una definitiva
interferencia con los procesos naturales? Lo que sí resulta claro es que la
capacidad de manipulación, no siempre basada en el conocimiento, está aumentando
continuamente. En particular, durante los últimos 15 a 20 años ha sido notable
un avance sin precedente en el campo de las ciencias biológicas. Esto se debe
fundamentalmente al surgimiento de las técnicas de ingeniería genética o ADN
recombinante.1![]()
Una característica importante de la ciencia biológica es que constituye una área de conocimiento relativamente nueva. ¿A qué se debe esto? ¿Cómo es, nos preguntamos, que en la era espacial y de las computadoras, no sabemos curar ni un catarro común? Parte de la respuesta estriba en la inmensa complejidad de cualquier sistema biológico. La célula más simple está constituida por cientos de miles de moléculas diferentes, que son entidades pequeñísimas, en continua transformación. Existe además un constante dinamismo en la composición del material viviente, mediado por interacciones moleculares de increíble sutileza, sujetas a una compleja y estricta regulación. Por esta razón, se requirieron instrumentos propios de la era tecnológica (por ejemplo, el microscopio), para empezar a atisbar en la estructura de estos sistemas. Se ha requerido del avance concertado de la física, la química, la ciencia de materiales, y del progreso económico para poder acceder a la tecnología necesaria para el estudio de los sistemas vivientes. Otro hecho destacado es que la investigación científica tiene la particularidad de ir acumulando preguntas que no es posible resolver con la tecnología del momento; pero en cuanto ésta progresa, se agolpan las soluciones de muchas preguntas y, rápidamente, surgen otras nuevas. Una de estas técnicas o metodologías, que establece un parteaguas en la capacidad de indagación sobre los seres vivos es, precisamente, la ingeniería genética.
Para entender cabalmente la importancia central de esta nueva metodología es útil revisar algunos conceptos básicos sobre las moléculas de la vida y lo que se conocía antes de que surgiera esa disciplina.
A través de varios miles de millones de años de evolución, la diversidad biológica nos resulta apabullante y asombrosa. Cada organismo vivo, desde un ser humano hasta una pequeña planta, una bacteria o una mosca, parece ser una invención única y diferente. Si observamos con cuidado, sin embargo, detectamos muchos elementos en común. La clasificación de los seres vivos en reinos, órdenes, géneros, etc., obedece precisamente a esta clara noción de que los organismos vivos se parecen unos a otros. En el nivel molecular, los seres vivos se parecen increíblemente. Es de la combinación y concierto de interacciones de los mismos tipos de moléculas que un ser vivo difiere de otro. Esto es similar al caso de las computadoras (especialmente los programas que corren en ellas), que pueden diferenciarse notablemente unas de otras, a pesar de estar constituidas por circuitos o instrucciones muy similares.
Las moléculas de la vida surgieron hace quizá 3 ó 4 mil millones de años. Sus características y sus interacciones fundamentales han sido alteradas muy poco en todo este tiempo. De manera similar al crecimiento y evolución de una ciudad, un ser vivo no se puede "reinventar" continuamente. La evolución ha ocurrido partiendo de lo que ya hay, con modificaciones paulatinas. En la Roma de hoy distinguimos calles que transitó Julio César; en nuestras células hay funciones moleculares afines surgidas hace 3 mil millones de años.
¿Cuáles son estas moléculas centrales, unificadoras? En lo que resta de este capítulo, me propongo describir algunas de ellas, pero antes debo hacer algunas aclaraciones al lector.
Para entender los fenómenos biológicos no se requiere, como en otras disciplinas (por ejemplo, la física o la química), el manejo de conceptos abstractos o el dominio de las matemáticas. Por otra parte, la complejidad de los sistemas biológicos sí necesita un vocabulario especial. Más aún, la amplitud y profundidad con la que comprenden hoy los biólogos la naturaleza es realmente impresionante. Confío en que las descripciones de naturaleza técnica, inevitables para hablar un lenguaje común y para que la presentación del material que nos ocupa no resulte trivial, sean accesibles al lector. Creo que la recompensa de cubrir los pasajes más rudos será una percepción más cabal de la belleza y relevancia de los recientes descubrimientos. Como dijera Albert Einstein: "Las cosas deben ponerse tan simples como sea posible. Pero no más simples."
El fenómeno de la herencia es discernible por cualquier observador, pero su fundamento químico-biológico no fue siquiera sospechado hasta épocas relativamente recientes. Por ejemplo, los genes eran entidades abstractas en los trabajos pioneros del monje austríaco Gregorio Mendel.
La historia de los avances del conocimiento y los experimentos cruciales que dieron origen a la identificación del material genético es extremadamente interesante e ilustrativa. Aunque desde principios de siglo se estudiaban preparaciones biológicas con la idea de identificar sustancias responsables de sus notables cualidades, no fue sino hasta los años cuarenta que los investigadores estadunidenses Avery, McLeod y McCarty sugirieron, cautelosamente, que el material hereditario podría estar contenido en la sustancia llamada ácido nucleico. Durante los siguientes cinco años, trabajando con sistemas de separación y análisis relativamente primitivos, los investigadores del campo de la genética bioquímica demostraron, de manera inequívoca, que tal era el caso.
El escenario estaba listo para el desarrollo de una de las investigaciones más relevantes en la historia de la biología. El multicitado descubrimiento de la estructura tridimensional del ácido desoxirribonucleico (ADN), realizado por Watson y Crick, y cuya culminación fue en 1953, merece siempre una mención especial.
Este trabajo muestra varios conceptos que son piedra angular en la investigación
biológica moderna. En primer lugar, los experimentos realizados se basaron en
los adelantos de la física. Se utilizaron rayos X y conceptualizaciones teóricas
para inferir, a partir de patrones de manchas, la estructura de la muestra de
ADN analizada (véase en el capítulo VI una descripción
más extensa sobre la técnica de cristalografía de rayos X). En segundo lugar,
se hace notar la convicción de que, de la forma tridimensional de las moléculas
biológicas se pueden obtener importantes ideas respecto a la función de las
mismas. En tercer lugar, se destaca la confianza de que hay elementos y principios
universalmente aplicables a todo ser vivo, cuando se hace el análisis a nivel
molecular. En su investigación, Watson y Crick reunieron información sobre las
propiedades de difracción de rayos X del material recientemente identificado
como portador de la herencia. Al utilizar también datos generados por los químicos
sobre la naturaleza básica de esta molécula, se dieron a la tarea de proponer
un modelo que fuera consistente con estas observaciones. Los resultados de sus
investigaciones fueron publicados en 1953 en la revista Nature, en una
sola página. Es así como hoy día sabemos que los genes están constituidos por
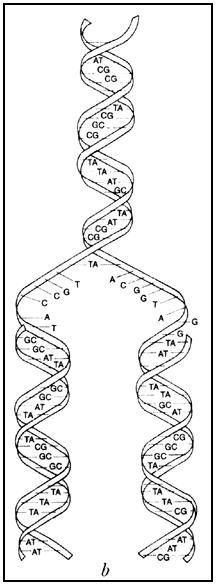
un polímero de entidades químicas, los nucleótidos arreglados en forma de escalera
de caracol, bautizado con el nombre de ácido desoxirribonucleico (ADN).
La estructura molecular de los ácidos nucleicos sugirió de inmediato algunas de sus notables propiedades (véase el recuadro I.1).
|
R
El ácido desoxirribonucleico o 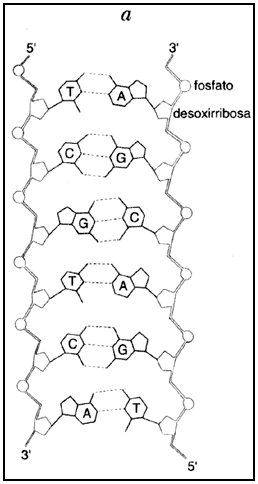
|
La complementariedad de las bases constituyentes de los ácidos nucleicos permite
almacenar y transferir información. La duplicación de estas moléculas se basa
precisamente en dicha propiedad. Si pensamos detenidamente en las características
fundamentales de las moléculas de ADN, observamos otros aspectos
muy interesantes: son moléculas químicamente monótonas, como inmensos rosarios
con sólo cuatro tipos de cuentas. El cromosoma de la bacteria más simple tiene,
por ejemplo, una longitud de alrededor de 600 mil pares de bases; los 23 distintos
cromosomas de una célula humana contienen mucho más ADN: ¡están
constituidos por aproximadamente 3 mil millones de pares de bases! Imaginemos
que miramos un collar de un millón de cuentas de cuatro tipos desde cierta distancia,
y que vemos otro, también de un millón de cuentas, arregladas en otro orden.
¿Cómo podríamos distinguir uno de otro? Sabemos que hay información muy valiosa
en el orden de esas cuentas, pero, ¿cómo tener acceso a ella? Antes del surgimiento
del ADN recombinante, la monotonía química del ADN
constituyó un obstáculo casi infranqueable para el progreso del conocimiento
en este campo.
A pesar de lo anterior; por medio de un gran número de experimentos, que no
consistían en la purificación de moléculas de ADN específicas,
ni su caracterización o análisis directo, se pudieron sentar las bases de su
funcionamiento fundamental. Así pues, sustentándose en pruebas de muchos tipos,
se estableció lo que se ha dado en llamar "dogma central de la biologia molecular"
(véase el recuadro 1.2).
|
R
La información contenida en la secuencia del 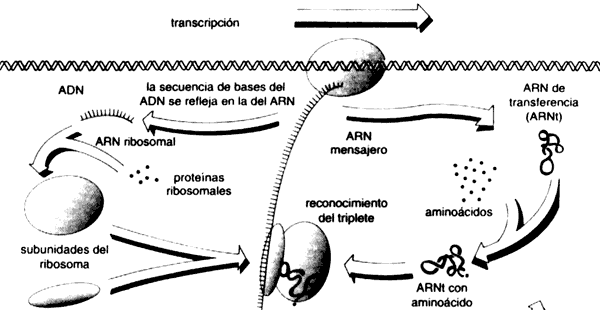
|
El concepto de que la información fluye de ADN a otra molécula
similar; el ARN, y de aquí hacia las proteínas, hace entrar en
escena a los otros actores protagónicos del conjunto de las macromoléculas biológicas:
las proteínas. La breve descripción del próximo inciso se refiere a ellas, pero
antes de iniciarlo, describiremos otra propiedad fundamental de los ácidos nucleicos:
su capacidad de hibridación. Las moléculas de ácidos nucleicos tienen la propiedad
de reasociarse si son separadas y, en general, de encontrar y asociarse con
otras moléculas cuya secuencia sea complementaria. Veamos con más cuidado este
concepto.
La complementariedad que pueden tener dos hebras de ácido nucleico se puede ilustrar si imaginamos una hebra con la siguiente secuencia: 5´CAGTGAATTCAATCGAT3´ y otra con la secuencia 5´ATCGATTGAATTCACTG3´ (los números 5´ y 3´ marcan los extremos de las hebras que corren en sentido antiparalelo, como se ilustra en el recuadro I.1). Estas dos especies moleculares son complementarias, es decir; si las colocamos una frente a otra, en sentido antiparalelo (tal y como se encuentran en las moléculas de ADN naturales), tenemos:
y observamos que frente a C (abajo de la C, en la representación mostrada), hay siempre G y viceversa, y frente a A, hay siempre T, y viceversa.
Podemos hacer dos consideraciones importantes respecto al ejemplo anterior. ¿Qué ocurriría si colocamos estas dos moléculas en un mismo tubo de ensayo? ¿Qué tan probable es encontrar una secuencia de ADN igual (o complementaria) a otra, dependiendo de su tamaño?
Respecto a la primera consideración se puede predecir que, bajo ciertas condiciones,
estas dos moléculas se unirían para formar una sola entidad: se aparearían o
hibridarían. Por lo que toca a la segunda pregunta, podemos hacer un cálculo
simple que nos indica que si tenemos cuatro símbolos, para un arreglo o "palabra"
de longitud 18, existen 418 combinaciones, es decir; unas 68 mil
millones. Si se compara con el lenguaje escrito, observamos que, en efecto,
las combinaciones de símbolos generan muy pronto secuencias únicas. No nos sorprendería
que, por ejemplo, una frase tal como "en Xochimilco mañana" (a pesar de ser
una frase corta, que tiene sentido y es correcta) no se encontrara en ninguno
de los libros editados por el Fondo de Cultura Económica. Similarmente, en todo
el genoma humano, la secuencia de ADN de nuestro ejemplo
podría no encontrarse. O si se encontrara una vez, sería muy poco probable que
hubiera una segunda. Esto quiere decir que la propiedad de hibridación confiere
a los ácidos nucleicos una capacidad de reconocimiento molecular altamente específica.
Aunque la propiedad de hibridación, o encontrar su complemento, de los ácidos nucleicos fue conocida desde hace mucho tiempo, no fue sino hasta el surgimiento del ADN recombinante que se pudo utilizar como herramienta para detectar secuencias de ADN o ARN (es decir; para aislar y cuantificar genes; véanse los capítulos III y VI).
Si observamos el proceso de expresión del material genético (véase el recuadro
I.2), observamos que, a partir de un código almacenado en moléculas químicamente
monótonas (ADN), se obtienen entidades moleculares individuales,
que resultan muy distintas unas de otras: las proteínas. Es notable
que en este proceso hay una correspondencia directa, lineal, entre la secuencia
de bases del ADN, y la secuencia de aminoácidos de una proteína
(las reglas de correspondencia se conocen como código genético (véase el recuadro
I.2). La diferencia entre estos dos polímeros biológicos estriba en que, en
los ácidos nucleicos los constituyentes o monómeros son sólo cuatro, y sus propiedades
fisicoquímicas son muy similares; en cambio, las proteínas están constituidas
por 20 tipos distintos de monómeros, con propiedades fisicoquímicas muy distintas.
La consecuencia de esta diferencia es que los ácidos nucleicos tienden a estructurarse
en largas hebras, cuyas propiedades generales son muy parecidas, independientemente
de la secuencia. Las proteínas, por su parte, son moléculas con gran personalidad
fisicoquímica.
Al estar constituidas las proteínas por diferentes aminoácidos, su secuencia determina que la hebra de una de ellas se pliegue en el espacio de manera específica. Así, aunque los genes que contienen las instrucciones (o codifican) para dos proteínas distintas son estructuralmente muy parecidos entre sí, las dos proteínas pueden tener estructura, propiedades y funciones totalmente distintas. Las proteínas se pliegan, obedeciendo a gran número de interacciones, cada una de magnitud relativamente pequeña, para formar estructuras específicas, características de cuya forma depende su función en última instancia (véase el capítulo VI).
Las proteínas son, pues, la herramienta de los genes, el brazo ejecutivo de la maquinaria celular; todo lo que existe en la biosfera resulta, en consecuencia, del trabajo concertado y regulado de las proteínas, dirigidas e interactuando con los genes y con el entorno.
Estas máquinas moleculares son capaces de llevar a cabo las más diversas funciones. Mediante la generación de polímeros con diversas secuencias, la evolución biológica ha producido proteínas capaces de transportar otras moléculas (por ejemplo, oxígeno en la hemoglobina de la sangre); de constituirse en materiales resistentes, flexibles o contráctiles (pelo, tendones, músculos); de transmitir información y alterar procesos (hormonas como la insulina, toxinas como la causante del tétanos o venenos de serpientes, arañas y alacranes); de transformar otras moléculas al catalizar reacciones químicas (enzimas).
Las enzimas son proteínas que merecen mención especial. Dentro de una célula viva existen miles de sustancias diferentes, que potencialmente pueden sufrir transformaciones químicas muy diversas. De hecho, sólo algunas de estas reacciones podrían ocurrir en condiciones suaves y a temperatura ambiente. La célula, sin embargo, sufre una constante transformación. Esto se debe al trabajo de las enzimas. Su capacidad catalítica consiste en que, al interaccionar con sus "moléculas blanco", es decir; aquellas a las que van a modificar (llamadas sustratos), facilitan transformaciones químicas específicas, sin alterarse ellas mismas en el proceso: una enzima cataliza o induce una transformación particular en miles de moléculas de sustrato sobre las que actúa en forma sucesiva. Así, de la acción concertada de las enzimas resulta un proceso continuo de transformación química, que es el responsable de la aparición de forma, función y adaptación al medio.
Es claro, entonces, que en cualquier fenómeno biológico las interacciones llevadas a cabo por las proteínas desempeñan un papel preponderante. De hecho, con gran intuición, en 1838, el químico holandés Gerardus J. Mulder fue quien dio nombre a estas sustancias utilizando la raíz griega protos, que significa: primordial o "de la mayor importancia". Resulta paradójico que quizá la mayoría de las veces, al escuchar la palabra proteína, pensamos en un componente de nuestros alimentos, y olvidamos la complejidad y la gran riqueza de funciones e información que encierran estas moléculas biológicas.
Carbohidratos, lípidos, esteroides, vitaminas, alcaloides, antibióticos...
Ciertamente, cuando hablamos de moléculas biológicas nos referimos a muchos compuestos que no son proteínas ni ácidos nucleicos. Gruesos tomos de bioquímica atestiguan la existencia de los muchos miles de moléculas y reacciones químicas que se llevan a cabo en cualquier ser vivo. Está por demás decir que una descripción de todas estas moléculas y sus funciones está mucho más allá del propósito de este libro y de mis capacidades. Creo, sin embargo, que no es arbitrario dedicar más espacio a explicar las propiedades de genes y proteínas, y mencionar solamente que hay innumerables compuestos esenciales que, organizados, transformados y reclutados por las proteínas, dan lugar al fenómeno de la vida.
SEPARACIÓN E IDENTIFICACIÓN DE LAS BIOMOLÉCULAS
Es usual que mis amigos, muchos de los cuales no son especialistas en ciencia, me pregunten con incredulidad: "bueno, ¿y cómo se ve a los genes?", "¿cómo saben que en un tubo de ensayo hay proteínas?", "¿cómo separan estos materiales, unos de otros?"
Como ya se mencionó anteriormente, es precisamente el avance metodológico el que permite responder preguntas y el que después capacita a formular otras nuevas. En la historia del desarrollo científico se puede observar la asombrosa capacidad del ingenio humano para intuir fenómenos que no ha podido observar directamente, y emitir avanzadas hipótesis respecto al funcionamiento de la naturaleza. Al mismo tiempo, se puede observar que es sólo mediante el uso de métodos, a veces extremadamente complejos, y de una tesonera experimentación, que estas hipótesis se pueden finalmente validar o descartar.
De la descripción de algunos de estos métodos se puede inferir, quizá, una sensación de lo que es una parte importante del diario quehacer de un científico experimental. De esta misma descripción podemos también deducir la noción fundamental de la importancia del surgimiento de las técnicas de ADN recombinante.
SEPARACIÓN Y ANÁLISIS DE LAS PROTEÍNAS
Imaginemos a los científicos alemanes Haus y Edward Buchner, quienes en 1897 investigaban las propiedades del proceso fermentativo, usando preparaciones derivadas de levaduras. A partir de un extracto de estos cultivos podían observar transformaciones sorprendentes, sin que hubiera ya células vivas. Se agregaba azúcar y, al cabo del tiempo, se notaba la aparición de alcohol. ¿Cómo podrían ellos identificar los componentes responsables de la reacción en la compleja muestra que tenían? En su época, los hermanos Buchner sólo contaban con pocas posibilidades, sin embargo, pudieron determinar que la reacción química que convertía el azúcar en alcohol dependía de dos "factores" presentes en el extracto. Uno de ellos se inactivaba al hervir la muestra, y otro, al dializarla (separar sus componentes por tamaño molecular de un lado u otro de una membrana). Conforme el fenómeno era estudiado, nuevas hipótesis se formulaban, y nuevos métodos se iban diseñando para contestar las preguntas formuladas. Tuvieron que pasar muchas décadas antes de que fuera claro lo que estaba ocurriendo, una vez que se acumularon conocimientos y nuevas metodologías. Estos experimentos iniciales, sin embargo, iban señalando el camino y pavimentándolo para el avance futuro.
Actualmente, en cambio, existen muy diversos y complejos métodos para separar y caracterizar biomoléculas. Los métodos modernos se basan en alguna propiedad molecular que, al interaccionar con el dispositivo experimental da origen a la separación. Ya mencionamos que el tamaño molecular se puede utilizar para realizar esta acción. En la actualidad disponemos de técnicas como la cromatografia de exclusión, la centrífugación y la electroforesis (véase el recuadro I.3) para la separación por tamaños. Afortunadamente, las proteínas son moléculas con tamaños muy variables y bien definidos. Una electroforesis del extracto de levadura mencionado revelaría un patrón de múltiples componentes, en el que se pueden ir identificando proteínas individuales.
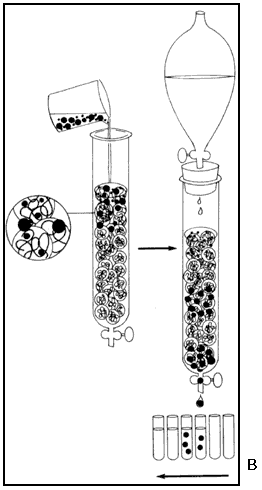
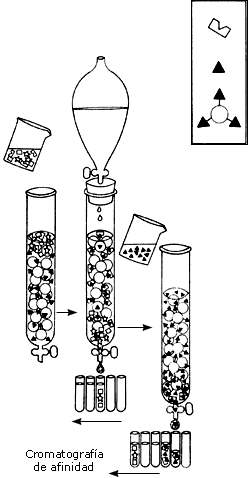
Otra importante propiedad que ayuda a la separación de las proteínas es su actividad. Cada proteína tiene una actividad particular que, en muchos casos, se puede medir. Si imaginamos que sometemos una muestra de saliva a un proceso de separación por tamaño molecular; y colectamos 100 fracciones en el proceso, ¿cómo sabemos en qué fracción está la actividad, por ejemplo, que degrada los azúcares? Si contamos con un método fácilmente visualizable, por ejemplo, una reacción química que origine un compuesto colorido, podemos ensayar esta reacción en cada uno de los tubos, y así conoceríamos el tamaño del componente activo de la saliva, y lo tendríamos parcialmente purificado.
De maneras similares a las descritas, se fueron identificando muchos de los componentes que constituyen a los seres vivos, y se les fueron asignando actividades. En los años setenta ya se disponía de la secuencia de aminoácidos de proteínas y de una descripción muy fina de su estructura, incluso en el nivel de resolución atómica (véase el capítulo VI).
Separación y análisis de los ácidos nucleicos
Como ya se mencionó, los ácidos nucleicos tienen apariencia de largas hebras de unidades básicas. Aunque, en principio, las técnicas de separación usadas para las proteínas también son aplicables para los ácidos nucleicos, aquí nos encontramos ante un problema: los genes no son entidades moleculares aisladas, con propiedades fisicas o químicas que los diferencien. Así, hasta la mitad de los años setenta, la estructura de los genes se había inferido sólo de manera general. Por medio de técnicas genéticas y de síntesis in vitro de polímeros de nucleotidos, se había podido definir el fundamento del almacenamiento, transmisión y expresión de la información genética; ¡pero no se había podido aislar o purificar; ni se conocía la secuencia de ningún gene en particular. Las técnicas de separación tenían una utilidad muy marginal, en tanto no se dispusiera de un ADN de tamaño manejable y naturaleza uniforme. El avance de este campo de la ciencia dependía críticamente de poder darle la vuelta a este problema. La sensación era, sin embargo, de desesperación al no ver una posible salida. Parecía no haber modo de tener acceso al genoma. La comunidad científica se encontraba sujeta al suplicio de Tántalo: sabía lo que buscaba y sabía que era extremadamente importante, pero no podía alcanzarlo.