II. LAS LLAVES DE LA BIBLIOTECA DE LA VIDA: LA NUEVA HERRAMIENTA DE LA INGENIERÍA GENÉTICA
La nueva herramienta: posibilidad de aislar, caracterizar y manipular los genes
LA INFORMACIÓN ya mostrada en los recuadros I.1 y I.2 sobre la
estructura y función básica de los genes se conocía desde finales de los años
sesenta. Más aún, la labor de los bioquímicos había producido un gran cúmulo
de conocimientos respecto de los conjuntos de reacciones químicas que ocurren
dentro de los seres vivos. Las vías metabólicas fundamentales para la generación
de energía y la biosíntesis de la mayoría de los compuestos esenciales para
la vida ya estaban establecidos.
Este conocimiento se basaba en la aplicación inteligente de métodos de ensayo y purificación de enzimas clave, y de la identificación y análisis de sus sustratos y productos. Al mismo tiempo, otras disciplinas como la inmunología y la genética, realizaban avances y eran actividades de gran complejidad y finura.
La biología molecular, sin embargo, se encontraba en un punto en el que su desarrollo se había hecho muy lento. Una vez sentadas las bases fundamentales de la replicación y expresión de la información genética, el siguiente paso era desentrañar; en forma detallada, los mecanismos de regulación de la expresión genética.
Todas las vías metabólicas, la expresión de anticuerpos, la actividad de los virus, en fin, todas las manifestaciones del fenómeno viviente, están, en última instancia, orquestados por la expresión ordenada y regulada de los genes, por medio de la producción de proteínas específicas.
El extraordinario proceso por el cual una sola célula, el huevo fecundado, da origen a un organismo complejo, es decir; las etapas de diferenciación celular; constituía un reto extremadamente atractivo, pero a la vez formidable. No se esperaba poder descifrarlo a un paso siquiera aceptable sin la capacidad de hacer una disección del genoma, de irlo analizando pieza por pieza.
Un descubrimiento clave inició el cambio dramático que conocemos hoy como ingeniería genética o ADN recombinante: en 1970 Hamilton Smith y Daniel Nathans descubren una enzima capaz de reconocer y cortar el ADN en secuencias específicas, que les valió el Premio Nobel de fisiología o medicina, compartido con Werner Arber, en 1978.
Este descubrimiento (consecuencia de un hallazgo accidental, en el que los investigadores profundizaron con gran sentido) dio origen a una sucesión de nuevos descubrimientos y potenció el desarrollo de toda una serie de otras disciplinas y metodologías. En este capítulo revisaremos los fundamentos de algunas de las más importantes.
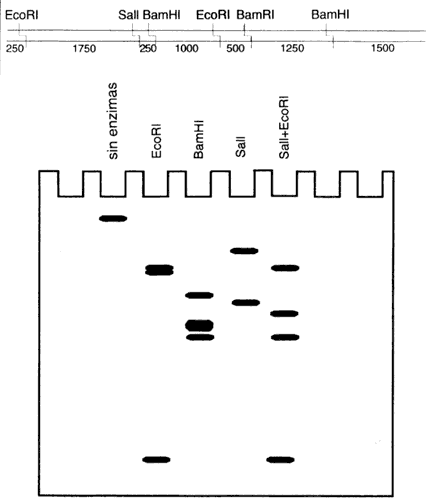
Muchos microorganismos producen enzimas que modifican y digieren o rompen el ADN. Estos sistemas, llamados de modificación-restricción, son análogos a un sistema inmune, los cuales probablemente evolucionaron como un mecanismo de protección de los microorganismos contra infecciones virales. En efecto, las bacterias, por ejemplo, son infectadas por virus, llamados bacteriófagos, que inyectan su propio ADN en la célula bacteriana para después controlar su maquinaria celular y redirigirla hacia la síntesis de sus propios componentes, dando como resultado final la ruptura de la célula y la liberación de cientos o miles de nuevos virus. En algunas bacterias el ADN propio está modificado en ciertas secuencias. Una enzima de modificación se desliza sobre la hebra de ADN, y cada vez que se topa con su secuencia blanco, por ejemplo GAATTC, introduce un pequeño grupo químico en la adenina (A) central. Otra enzima, la enzima de restricción, también se desliza en la hebra de ADN, y si encuentra la secuencia GAATTC, corta el ADN en esa posición. La enzima, sin embargo, no corta el ADN modificado, por lo que efectivamente es capaz de degradar el ADN extraño que puede entrar a la célula, sin alterar el ADN propio.
Hoy en día conocemos miles de diferentes enzimas de restricción, provenientes de otros tantos distintos microorganismos. Más de cien distintas secuencias pequeñas de ADN pueden ser rotas, específicamente, por medio del uso de la adecuada enzima de restricción.
Otra interesante propiedad de las enzimas de restricción es que, en general, reconocen secuencias palindrómicas, es decir; secuencias que son iguales si se leen en una dirección, o en la dirección contraria. Por ejemplo:
es una secuencia palindrómica. Cuando actúa la enzima que reconoce y corta
esta secuencia, llamada EcoRI, genera segmentos de ADN con
extremos que se proyectan fuera de la doble cadena:
Estos extremos se denominan cohesivos o pegajosos, porque tienden
a aparearse o hibridarse nuevamente. En realidad, cualquier extremo de ADN
generado por un corte con EcoRI puede hibridarse o aparearse con
otro extremo generado por la misma enzima.
Los descubridores de las primeras enzimas de restricción pronto se dieron cuenta
de que su acción sobre el ADN (comúnmente llamada "digestión")
producía un conjunto definido de diferentes segmentos. Esto es particularmente
fácil de detectar si la digestión se efectúa sobre una molécula pequeña; y tales
moléculas existen en la naturaleza. Por ejemplo, ciertos virus están constituidos
por genomas muy pequeños. El virus SV4O (que ocasiona cáncer en los simios)
contiene una molécula de ADN circular de unos 5 mil pares de bases.
Muchas bacterias portan pequeñas moléculas de ADN, llamadas plásmidos,
que llevan información accesoria a su cromosoma. Estas moléculas pueden
tener también sólo unos miles de nucleótidos. Si se somete el producto de digestión
de una de estas moléculas a una separación por electroforesis, después de utilizar
la enzima de restricción, se observa un patrón de bandas, que corresponde
a los fragmentos de ADN de tamaños correspondientes a la distancia
entre un sitio y otro. El principio del proceso es análogo a la separación de
proteínas (véase el recuadro II.1).
Súbitamente, el ADN dejó de ser una sustancia monótona y frágil,
donde purificar un segmento específico resultaba una tarea ardua o imposible.
Al poder generar segmentos específicos, con las enzimas de restricción, la molécula
de la vida quedó a merced del biólogo molecular. Es como si alguien jalara la
cinta de un casete de audio, la amontonara desordenadamente y luego nos pidiera
que separáramos el segmento que contiene una canción. Sería útil disponer de
unas tijeras que cortaran la cinta cada vez que apareciera la sucesión de notas
si, la, sol, do, re, mi, y después un ayudante acomodara los segmentos
resultantes, de acuerdo con su tamaño. O imaginemos lo dificil que seria identificar
un artículo de la enciclopedia, si está escrito sobre un listón de varios kilómetros
de largo. Cómo se facilitaría la tarea si nuestras tijeras encontraran automáticamente,
por ejemplo, la palabra "obtener", y ahí cortaran el listón. Luego nuestro ayudante
nos presentaría 20 ó 30 cajas, cada una conteniendo pedazos de listón
clasificados de acuerdo con su tamaño.
CONCEPTO DE CLONACIÓN MOLECULAR
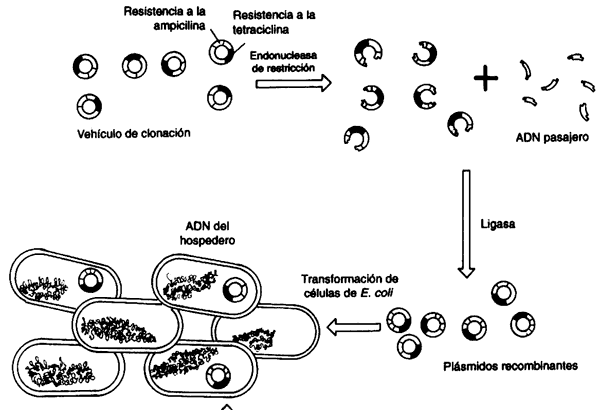
Como ya mencionamos, los fragmentos generados por las enzimas de restricción tienen además la propiedad de reasociarse unos con otros, por medio de sus extremos cohesivos. Cuando los investigadores de la genética bacteriana conocieron estas enzimas de reciente caracterización, a principios de los años setenta, se dieron cuenta de algo sumamente importante: si se reasocia un segmento de restricción proveniente de un organismo con otro segmento, generado por la misma enzima pero proveniente de otro organismo, se obtendría una molécula híbrida o quimérica, una molécula de ADN recombinante. De hecho, dado que el ADN de todos los organismos vivientes tiene una naturaleza química idéntica, no deberían existir limitaciones para recombinar el ADN de cualquier origen. Algunos investigadores intuyeron el monumental potencial de este concepto. Los laboratorios de Paul Berg, Stanley Cohen (en la Universidad de Stanford) y de Herbert Boyer (en la Universidad de California, San Francisco) se dieron así a la tarea de hacer que este concepto se convirtiera en realidad.
Para reasociar dos moléculas de ADN es necesario reconstituir
el enlace fosfodiéster (un enlace covalente, fuerte), y aquí entra en acción
otra enzima que había ya sido descrita para entonces: la ADN ligasa.
De esta forma podían tomarse las dos moléculas, reasociarlas (hibridizarlas)
y luego ligarlas, se obtiene una molécula continua, recombinante, la cual, sin
embargo, es algo inerte en tanto no sea introducida a una célula que la transcriba
y traduzca (véase el recuadro I.2). Además, algo muy importante:
esta molécula debe ser capaz de perpetuarse (replicarse dentro de la célula).
De otra manera se diluiría y perdería irremisiblemente después de que la célula
se dividiera. Finalmente, era necesario identificar las células en las que el
ADN recombinante se hubiera introducido y establecido.
Así tenemos los elementos esenciales de la técnica de ADN recombinante (véase el recuadro II.2):
- Obtención de fragmentos específicos de
ADN(enzimas de restricción). - Ligación (o reasociación covalente) de las moléculas para obtener hebras
continuas de
ADN(enzima ligasa). - Mecanismo para introducir al interior de células vivas el
ADNrecombinante (transformación). - Mecanismo para asegurar la replicación e identificación de la molécula recombinante dentro de la célula (vehículo molecular).
El primer experimento en el que se puso en práctica este concepto, realmente
simple, que se conoce como clonación molecular se logró al utilizar plásmidos
bacterianos como vehículos y ADN, también bacteriano, como pasajero.
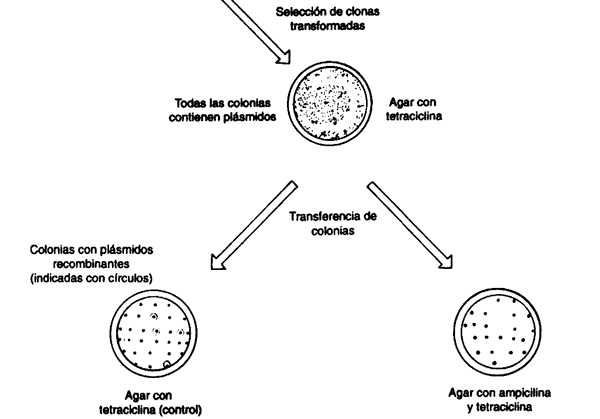
Uno de los primeros vehículos moleculares de donación, el plásmido denominado pBR322, fue construido por el doctor Francisco Bolívar, investigador mexicano que se encontraba trabajando en su postdoctorado, a mitad de los años setenta, en la Universidad de California, San Francisco. Este plásmido se ha usado innumerables veces y servido de base para la construcción de la mayoría de los vehículos más modernos de donación. Por la participación del doctor Bolívar en estas investigaciones, aunado a sus otras aportaciones a la ciencia mexicana e internacional, fue galardonado con el premio internacional Príncipe de Asturias en el área científica, en 1992, que otorga el gobierno español.
Consideremos ahora las consecuencias de un experimento de donación molecular.
Los segmentos de ADN que se encontraban dispersos en largas y monótonas
hebras de ADN, resultan ahora disociados en segmentos específicos,
y cada uno puede ser perpetuado independientemente. Disponemos ahora de una
biblioteca genómica o genoteca, que podemos mantener indefinidamente con sólo
mantener vivas las células bacterianas que albergan los plásmidos recombinantes.
Igualmente, una vez seleccionada una clona de la genoteca, pudimos cultivar
las células y obtener cantidades ilimitadas de ADN para su posterior
manipulación y caracterización. La elusiva purificación de genes específicos
se vuelve una realidad.
Como ya se mencionó, las repercusiones de estos primeros experimentos fueron visualizadas rápidamente, pues incluían también posibles riesgos y problemas, lo cual dio origen a un importante debate, iniciado por la propia comunidad científica (véase el recuadro II.3).
Durante la década siguiente, en un proceso continuo y creciente de desarrollo metodológico, se constituyó un verdadero arsenal de diferentes herramientas biológicas que aumentaron notablemente la capacidad de aislar; caracterizar y manipular los genes. (La descripción de algunas de las técnicas básicas más importantes se destaca en lo que resta del capítulo. En el capítulo siguiente se ven ejemplos de su aplicación y se describen otras técnicas importantes)
Desde la época de los experimentos pioneros en que se definió el código genético
fue necesario elaborar moléculas de ácidos nucleicos no naturales. Se inició
entonces el desarrollo de las técnicas para sintetizar o fabricar químicamente
el ADN. El desenvolvimiento de la química orgánica, que precedió
al de la biología molecular; había establecido desde tiempo atrás la naturaleza
química de los ácidos nucleicos. Se requirió, sin embargo, un periodo relativamente
largo para perfeccionar adecuadamente las técnicas para sintetizar el ADN
de manera práctica. Desde fines de los años sesenta, Har Cobind Khorana logró
la tarea titánica de elaborar químicamente un pequeño gene a partir de sus precursores
básicos (los nucleótidos A, G, C y T). Este trabajo hizo al doctor Khorana merecedor
del Premio Nobel de fisiología o medicina en 1968. Los siguientes quince años
fueron necesarios para que una técnica laboriosa, que requería la participación
de químicos especializados, se convirtiera en una tarea simple y automatizada,
al alcance de cientos de laboratorios del mundo. Actualmente, mediante síntesis
química se obtienen miles de fragmentos de ADN cada día. El factor
clave para la simplificación del procedimiento consistió en lograr la síntesis
en fase sólida, que es susceptible de ser adaptada a una máquina automática
(véase el recuadro II.4). Las limitaciones de esta técnica sólo permiten sintetizar
directamente fragmentos de ADN de un tamaño menor a unas 100 bases,
por lo que normalmente se les conoce como oligonucleótidos (o simplemente oligos).
De cualquier manera, utilizando las propiedades de hibridación del ADN
y de la enzima ADN ligasa, se pueden construir grandes trechos
de ADN de doble cadena.
Al disponer de oligos, cuya secuencia está definida por el investigador, se abren posibilidades para crear genes con completa libertad, y también para modificar, en principio sin limitación alguna, a los genes naturales (véase el recuadro II.4). (En secciones sucesivas se verán algunas mas de las múltiples aplicaciones de los oligos sintéticos.)
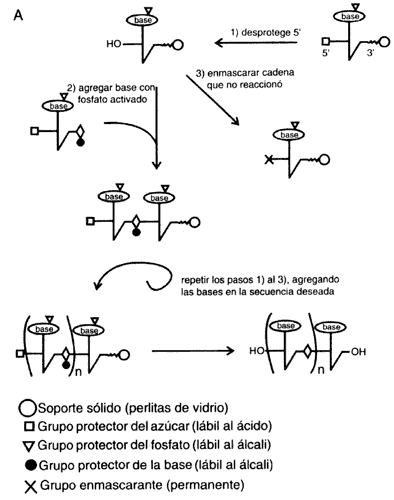
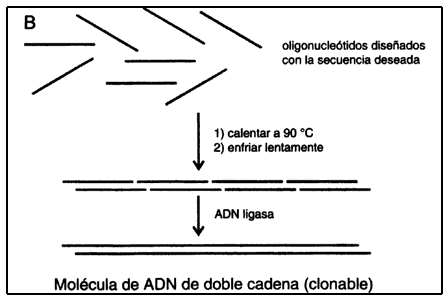
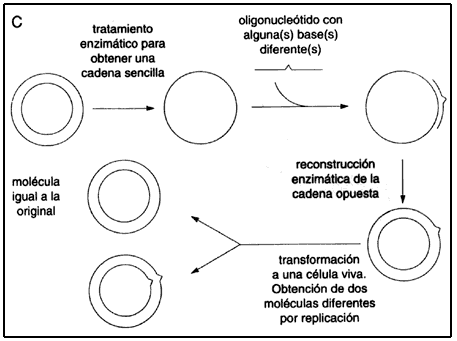
ENZIMAS ÚTILES PARA MANIPULAR EL ADN
Hasta el momento hemos mencionado a la ADN ligasa, una importante
enzima que permite ligar o unir moléculas de ADN. La investigación
sobre la fisiología de los ácidos nucleicos ha proporcionado, además, una serie
de enzimas que se utilizan ampliamente para manipular los ácidos nucleicos en
el tubo de ensayo, y que forman parte importante de las herramientas del ADN
recombinante. Cada una de estas enzimas, al igual que las endonucleasas de restricción,
cumple un papel dentro de la célula viva, para alterar el material genético
(por ejemplo, para repararlo, replicarlo, degradarlo, recombinarlo, etc.). En
el laboratorio se pueden usar estas enzimas, después de purificarlas de sus
fuentes naturales, para efectuar transformaciones útiles a los propósitos del
experimentador. Por ejemplo, la enzima transcriptasa reversa, que naturalmente
producen ciertos virus para invadir el genoma de sus células blanco (véase el
recuadro IV.3), se utiliza ampliamente para clonar genes a partir de sus ARN
mensajeros (véase el capitulo siguiente).
Dado que el propio desarrollo del ADN recombinante, ha potenciado,
a su vez el desarrollo de herramientas moleculares, hoy en día contamos con
un enorme número de enzimas y reactivos que permiten gran versatilidad en la
manipulacion del ADN.
Una consecuencia inmediata de la clonacion molecular es que permite disponer
de cantidades ilimitadas de segmentos específicos de ADN.
Debido a que el ADN pasajero se encuentra formando parte de un plásmido, es
factible separarlo fácilmente del resto del ADN celular, que se
encuentra en el cromosoma bacteriano, una molécula mucho más grande. A partir
de un cultivo de E. coli, se puede separar suficiente ADN plasmídico
para caracterizarlo. Una vez que se dispuso de genes individuales purificados,
el escenario estaba listo para la siguiente etapa.
Walter Gilbert, en la Universidad de Harvard, y Frederic Sanger, en Cambridge,
Inglaterra, se dieron a la tarea de desarrollar técnicas para determinar la
más importante característica del ADN, su secuencia de bases. Pocos
años después lograron su objetivo, y compartieron el Premio Nobel de química
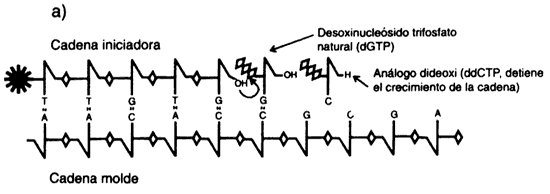
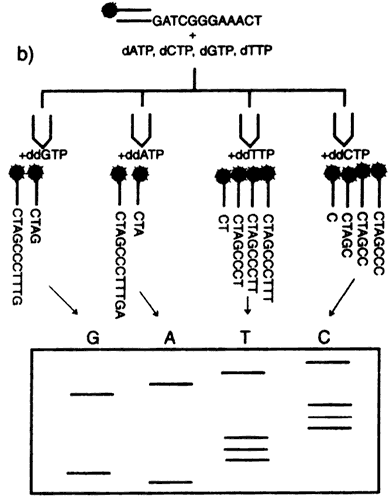
en 1980. Las técnicas de secuenciación actuales (véase el recuadro II.5) son
usadas abundantemente e, inclusive, han sido automatizadas. A la fecha, genes
de las más diversas fuentes han sido secuenciados, acumulando una base de datos
que representa cientos de millones de nucleótidos. Esta cantidad crece a paso
inexorable y se espera que lo haga cada vez más aceleradamente (véase la sección
sobre la secuenciación del genoma humano, en el capítulo IV).
LA REACCIÓN EN CADENA DE POLIMERASA
Otro de los avances metodológicos más importantes es la reacción en cadena
de polimerasa (PCR, siglas en ingles polymerase chain reaction).
Este procedimiento constituye un ejemplo sumamente interesante de la importancia
de la metodología en el desarrollo científico. También ilustra la manera como
ocurren muchos de los descubrimientos: a partir de intuiciones que integran
conocimientos que han estado disponibles durante cierto tiempo.
En efecto hacia 1985 —con el ADN recombinante ya plenamente
establecido como una técnica aplicada rutinariamente por cientos de laboratorios
en el mundo—, Kary Mullis, quien a la sazón trabajaba para la compañía Cetus,
en San Francisco, California, tuvo una súbita inspiración mientras manejaba
su auto y se dirigía hacia una cabaña en la que se disponía descansar el fin
de semana. Como muchos otros investigadores, Mullis se encontraba desarrollando
aplicaciones para los oligos sintéticos. Concibió entonces la idea de que combinando
el uso de los oligos con varios ciclos de replicación in vitro se podía
amplificar, es decir; obtener en gran cantidad, un segmento de ADN
específico (por ejemplo, un gene en particular).
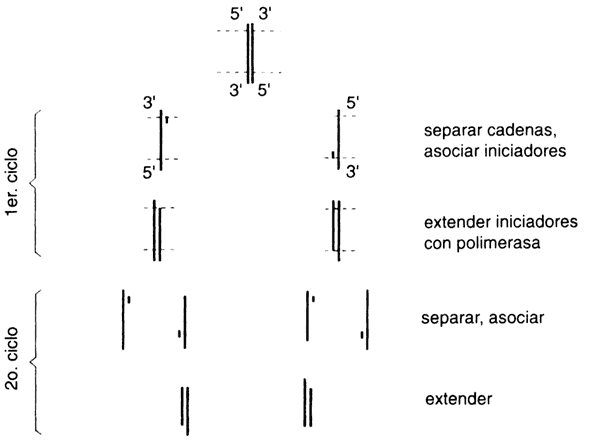
Pero veamos con más calma en qué consiste la idea (figura II.1). La enzima
ADN polimerasa participa en la replicación del ADN
(recuadro I.I). Para convertir una molécula de ADN de doble cadena
en dos moléculas idénticas, la enzima requiere que las cadenas de la molécula
inicial se separen, para así tener un molde disponible. Además, la enzima requiere
un segmento de ADN con un extremo libre, que sirve para cebar o
iniciar la reacción de replicación y los nucleótidos precursores. Si se colocan
estos sustratos en un tubo de ensayo, la ADN polimerasa puede incorporar
uno por uno los nucleótidos correspondientes, es decir, frente a una A en el
molde, adicionará T en la cadena creciente; frente a G, C, etc. En realidad,
éste concepto era perfectamente conocido y aplicado desde mucho antes de que
se concibiera la técnica de la PCR. Lo original de la idea de la
concepción de Mullis fue pensar qué sucedería si se aplica este procedimiento
en ciclos sucesivos, en una reacción en cadena. Los oligos sintéticos, en virtud
de su propiedad de hibridación de los ácidos nucleicos, definen los puntos de
partida para el copiado de las cadenas de ADN. Si se colocan dos
oligos cuya secuencia define los extremos de un segmento determinado de ADN,
se hacen hibridar con las hebras del molde, previamente separadas, y se someten
a una reacción con ADN polimerasa, de lo que resultarán dos moléculas
cuyos extremos están definidos por los oligos y que corren en sentido opuesto.
Las cadenas resultantes son complementarias entre si: existen ahora el doble
de moléculas con la secuencia que demarcan los oligos. ¿Qué pasa si ese proceso
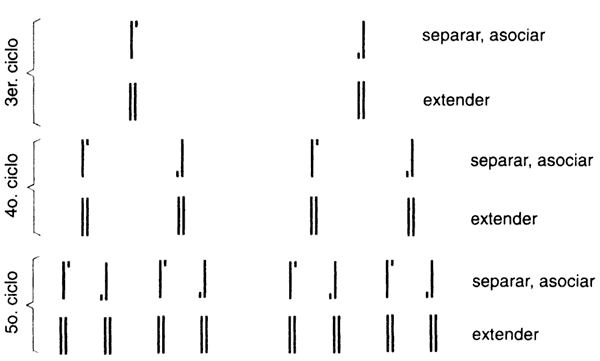
se repite, cíclicamente? ¡El resultado es lo que se denomina un crecimiento
exponencial! En el siguiente ciclo de separación de cadenas, hibridación con
los oligos y reacción con ADN polimerasa, se obtendrán cuatro moléculas
de la región entre los oligos. Los ciclos subsiguientes generarán 8, 16, 32,
64 moléculas, y así sucesivamente. Para comprender mejor las implicaciones de
un proceso exponencial, recordemos la fábula del inventor del ajedrez. Se cuenta
que un rey quedó tan asombrado de lo interesante que resultaba el juego, que
ofreció recompensar al súbdito que se lo enseñó. Este le pidió "solamente" un
grano de trigo por el primer cuadro del tablero, dos por el segundo, cuatro
por el tercero, y así sucesivamente. El rey, sin hacer cálculos, accedió de
buen grado. Por desgracia para el rey, todas las cosechas de trigo del mundo
fueron insuficientes para completar la solicitud del inteligente vasallo. Sólo
por el cuadro 30 se requerían 1 073 741 824 granos de trigo.
  |
Así, con la técnica de la PCR se pueden amplificar segmentos específicos
de ADN para crear muchos millones de moléculas a partir de unas
cuantas, o incluso de una sola. Para esto se requirió adaptar el concepto básico
haciendo uso de ADN polimerasas provenientes de microorganismos
termófilos (que crecen a temperaturas de más de 70 grados en manantiales termales).
De otra manera, la enzima se inactivaba cada ciclo, pues se requiere aplicar
calor para separar las moléculas del ADN molde. Hoy día un ciclo
puede tomar cinco minutos o menos, por lo que el proceso de amplificación se
lleva a cabo en menos de dos horas (normalmente 20 ó 30 ciclos son suficientes).
Nuevamente nos encontramos ante un concepto bellamente simple. Lo sorprendente es que los elementos técnicos y conceptuales necesarios para esta invención estuvieron disponibles por varios años antes de que Kary Mullis los desarrollara.
La aplicación de la PCR, en sí misma, constituye un avance revolucionario en la investigación biológica moderna. Uno de los más espectaculares ejemplos que ilustran la potencia de esta técnica es el rescate y secuenciación de ADN prehistórico (realidad que sustenta la historia de ciencia ficción Parque Jurásico de Michael Crichton). (En capítulos siguientes consideraremos otras aplicaciones en las que la PCR desempeña un papel indispensable.)
OTRAS TÉCNICAS IMPORTANTES EN BIOLOGÍA EXPERIMENTAL MODERNA
En los capítulos que siguen hablaremos de las muchas maneras en que los investigadores aplican métodos específicos para lograr sus objetivos de aislar, caracterizar y manipular genes y sus productos. Cada problema requiere echar mano de las técnicas más modernas y avanzadas. Efectivamente, en la práctica, la labor de un científico está fuertemente condicionada por las técnicas experimentales de que dispone y puede manejar.
Así, entre el gran número de técnicas que no mencionamos en este capitulo, algunas harán su aparición cuando se describan aplicaciones especificas. Observaremos cómo se utiliza reiteradamente la propiedad de reconocimiento molecular, de asociarse específicamente tras moléculas. ¡Realmente el científico moderno tiene las herramientas para encontrar una aguja en un pajar! Trataremos de resaltar también cómo los avances de tecnologías que provienen de disciplinas diferentes son incorporadas ávidamente por los biólogos. Su uso les permite prosperar en las respuestas a las preguntas que se están formulando.