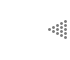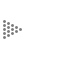IX. VIAJANDO POR EL MUNDO
PARA que una planta se reproduzca con éxito, una vez formadas
sus semillas, es necesario que se distribuyan o dispersen a lugares donde puedan
germinar y establecerse. De este modo, una vez que la planta se establece, la
información genética de los progenitores queda en el ambiente. A su vez, cuando
esa nueva planta forme semillas, su información genética estará en las semillas
que produzca y si logran establecerse quedará en el ambiente. Así, las características
genéticas de una planta, que incluyen el color de las flores, la intensidad
del aroma y su resistencia a plagas, entre otras muchas, se irán transmitiendo
de padres a hijos y estarán presentes en el ambiente, pues siempre habrá plantas
con esas características.
Cada especie tiene sus propios requerimientos de germinación (capítulo VI), por lo que no cualquier sitio es adecuado. Las características idóneas varían según la especie. Los diferentes modelos o estrategias de dispersión de las semillas quizá sean resultado de las fuerzas que la selección natural ha ejercido sobre ciertas características de las semillas, las cuales incrementan las probabilidades de que lleguen a un sitio adecuado para germinar y establecerse.
Al analizar el modelo que sigue un individuo para depositar sus semillas (sombra
de las semillas), se observa que la mayoría está concentrada cerca de la planta
que las produjo, y conforme aumenta la distancia, va disminuyendo su número
(Figura IX.1). La distancia a la que se deposita el mayor número de semillas
y la distancia máxima que alcanzan con respecto a la planta adulta varía entre
las especies. Por ejemplo, en el caso de las semillas dispersadas por el viento,
la distancia que alcanzan es mayor que en el caso de las semillas dispersadas
por un ratón.
Figura IX.1. Patrón de sombra de las semillas dispersadas. El número mayor
de semillas cae alrededor de la planta que las produce y conforme aumenta la
distancia van dispersándose cada vez en menor número (tomado de Poole y Cairns,
1940, Bakker, 1960 y Roe, 1967 en Harper, 1977). Es importante hacer notar las
diferentes distancias a las cuales las semillas se dispersan y que aparecen
en el eje horizontal (eje X) de las graficas.
Como se había mencionado anteriormente, los sitios adecuados para germinar y establecerse varían según la especie. Hay semillas que requieren de luz y fluctuaciones de temperatura para germinar, por lo tanto, los sitios adecuados serán aquellos en los que exista un claro sin vegetación, donde no haya una cubierta de ramas y el Sol incida directamente en el suelo, y además, donde no haya depredadores. Por consiguiente, las semillas de esta especie tendrán que dispersarse hasta zonas donde haya claros, tal vez a cierta distancia de la planta que las produjo, o bien sería necesario que hubieran sido producidas en suficiente número como para que existiera una buena probabilidad de que llegara a los pequeños claros que existen dentro del propio bosque. En este ejemplo los sitios adecuados están más o menos retirados de la planta progenitora, por lo que la selección natural sobre el modelo de dispersión habrá favorecido el que pocas semillas se mantengan cerca de la planta productora y que mas bien se dispersen abarcando una distancia grande. En este caso también la selección natural habrá favorecido la producción de numerosas semillas de tamaño pequeño que fácilmente puedan ser transportadas por el viento y en suficiente número para alcanzar los claros dispersos entre la vegetación.
Una característica importante de los sitios adecuados para germinar y establecerse es que no haya depredadores de las semillas (capítulo X). Éstos pueden ser desde roedores que se lleven las semillas a su madriguera hasta insectos que pongan sus huevecillos en las semillas para que las larvas se desarrollen alimentándose de ellos. En general para un depredador es mucho más sencillo encontrar las semillas cuando están juntas o cercanas entre sí; por lo tanto, la probabilidad de que las semillas que se dispersan en alto número alrededor de la planta sean depredadas es mayor que para las semillas que estan más alejadas y más aisladas entre sí.
El transportar y alejar a las semillas de la planta progenitora generalmente requiere de un agente externo de dispersión como es el agua, viento o animales. En ocasiones las semillas presentan una primera fase de dispersión que involucra a un agente que las separa a corta distancia de la planta progenitora y una segunda fase de dispersión, a veces con otro agente, que les permite separarse a mayores distancias.
La dispersión de las semillas es uno de los capítulos más interesantes en la vida de la planta. En esta fase la naturaleza ha desplegado una enorme creatividad de formas, situaciones y respuestas. La variedad es inmensa y sería imposible relatar todos los casos. Con la idea de poder percibir estas respuestas múltiples, presentaré a lo largo del texto varios ejemplos de cómo las semillas logran dispersarse.
1) Dispersión por viento (anemocoria)
Dentro de esta categoría caen tanto las semillas muy pequeñas (menores a 0.05 mg, como las semillas de orquídeas) como aquellas que tienen estructuras especiales que les permiten usar las corrientes de viento para dispersarse.
Como ya se mencionó anteriormente, una de las estrategias más sencillas es el tener un tamaño pequeñísimo, con una proporción superficie/volumen grande. Estas semillas se producen en enormes cantidades y se transportan en las corrientes de viento igual que el polvo. Este modo de dispersión es característico de muchas orquídeas, parásitas y saprófitas. Entre las principales familias que presentan este tipo de dispersión están las orquídeas, droseráceas, nepentáceas, escrofulariáceas, etc. El tamaño pequeño priva a la semilla de tener reservas, por lo que frecuentemente está relacionado con la dependencia nutricional de otro organismo (capítulo VIII).
Una de las familias cuyas semillas típicamente se dispersan por viento es la de las compuestas. Presentan frutos de una sola semilla llamados aquenios que constituyen la estructura que se dispersa. Frecuentemente tienen apéndices que les permiten ser transportados por las corrientes de viento y que también actúan como una especie de paracaídas que les permite depositarse en el suelo. A estas estructuras se les llama pluma (pappus, vilano) y una semilla puede tener desde una hasta varias, lo cual le da un aspecto peludo (Figura IX.2(a)). Estas semillas pueden transportarse a mayores distancias que las semillas aladas. Comparando semillas sin ninguna estructura especial de dispersión con semillas con apéndices tipo pluma, se encontró que las primeras se separan sólo de uno a tres metros de la planta progenitora, mientras que las segundas lo hacen entre 40 (Asclepias syriaca) y 80 metros (Apocynum sibiricum).
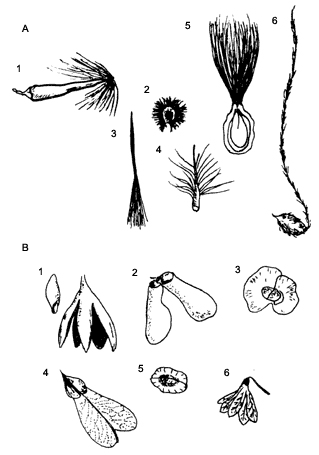
Figura IX.2. A) Semillas con los diferentes tipos de estructuras en forma
de plumas (pappus o vilano) que ayudan a las semillas a dispersarse con el viento.
1. Platanus occidentails, 2. Bellotia mexicana, 3. Rhabdadenia
biflora, 4. Tamarix parvifolia, 5. Asclepias curassavica, 6.
Clematis spp. B) Semillas con los tipos de alas que las ayudan a dispersarse
usando el viento y a planear durante el descenso. 1. Fruto y semilla de la Cedrela
odorata, 2. Heteropteris macrostachya, 3. Hiraea affinis, 4.
Thinovia myriantha, 5. Urvillea almacea, 6. Petrea bracteata.
Estas estructuras también permiten que la semilla, al separarse de la planta progenitora, disminuya su velocidad de caída y se incremente la probabilidad de que las corrientes de viento la transporten. Entre los factores que pueden afectar la distancia a la que son transportadas (forma de la sombra de semillas) están la forma y tamaño de la semilla, la altura de la planta que las produce y por lo tanto a la cual se liberan, la velocidad del viento y la densidad de la vegetación que las rodea. Este último constituye un aspecto importante en la dispersión de las semillas. Algunas plantas como las gramíneas producen sus frutos en el extremo de tallos alargados que sobresalen de la vegetación, exponiéndolos así a las corrientes de aire.
En ocasiones una testa o bien alguna cubierta del fruto, está separada y suelta de modo que ayuda a formar una estructura globosa que permite la dispersión por viento. Se encuentran frutos globosos por ejemplo en el género Cardiospermum de las sapindáceas y en algunas rosáceas.
Ocasionalmente el fruto puede tener cierto control sobre la distancia a la que las semillas se dispersan. En las umbelas de Daucus carota las semillas se encuentran en la punta de los pedicelos que sirven de unión con la planta progenitora. Los frutos reaccionan a la humedad relativa del aire extendiendo los pedicelos. Así, una umbela que responde rápidamente a la humedad relativa, extiende o abre los pedicelos, y las semillas quedan más expuestas y fácilmente pueden ser desprendidas y transportadas por un viento suave, por lo que estas semillas se dispersan rápidamente y a distancias cercanas. En cambio, una umbela que responde más lentamente, tiene las semillas menos expuestas y requiere de un viento más intenso para desprenderlas por lo que se dispersarán a distancias mayores.
Estos apéndices pueden tener formas muy variadas (Figura IX.2). Además de capacitar a la semilla para flotar en el aire, pueden tener otras funciones como la de orientar a la semilla durante su caída de modo que aterrize en el suelo en cierta posición específica (como en el caso de pastos de las estepas de los géneros Stipa y Aristida), ahuyentar a algún depredador o bien permitirle más movimiento ya estando en el suelo. A veces también incrementan la superficie de la semilla permitiéndole flotar en corrientes de agua (Bombax munguba).
Otro apéndice importante para la dispersión por viento son las alas (Figura IX.2(b)). Ayudan tanto a la flotación de la semilla como a su orientación. Algunas alas son enormes, como por ejemplo las de Dipterocarpus grandiflorus, cuyas semillas miden 6 cm de ancho y el ala 14 cm de largo; Macrozanonia macrocarpa, una cucurbitácea trepadora de selvas, cuyas alas laterales se extienden a ambos lados de la semilla y entre ambas abarcan 15 cm. Hay semillas con una sola ala llamadas samaras que permiten una propulsión lateral, como las del Acer y las de algunas leguminosas (Dalbergia, Pterocarpus). Este tipo de alas son muy comunes y también pueden llegar a ser muy grandes como la de Centrolobium robustum, planta brasileña en que el ala mide 17 cm. Otras especies presentan varias alas, lo cual les permite rotar y cuando son planas deslizarse en el aire. Entre las más llamativas están las de la bombacácea del género Cavanillesia del Amazonas, en la que las alas llegan a medir hasta 9 cm de ancho.
Existen especies en las cuales toda la planta o parte de la planta es lo que se dispersa. Son especies como Agrostis hemialis que habitan en ambientes con vegetación abierta donde corre el viento libremente, como en algunos desiertos o estepas, por ejemplo. En estas especies la planta entera o una parte se desprende y rueda empujada por el viento. Las distancias a las que se dispersan las semillas de estas plantas son mayores (9 m) cuando las semillas permanecen adheridas al fruto y ruedan con toda la planta que cuando caen directamente del fruto (2 m) antes de que la planta empiece a rodar. Otra especie cuya infrutescencia también rueda es Spinifex squarrosus, que habita las playas de la Isla de Java, al igual que otras especies de este mismo género que viven en las estepas de Australia.
Las semillas dispersadas por viento son distribuidas en el ambiente separadas entre sí, ya que son transportadas "de una en una". Por tanto es poco probable que las plántulas de la misma especie germinen cerca y compitan entre sí por recursos.
Durante las tormentas, la velocidad del viento aumenta considerablemente, dispersando las semillas a distancias mayores. Se han registrado distancias de varios kilómetros para algunas especies como:
| Abies |
7 |
kilómetros |
| Pinus sylvestris |
2 |
kilómetros |
| Betula |
1.6 |
kilómetros |
| Acer |
4 |
kilómetros |
| Fraxinus excelsior |
0.5 |
kilómetros |
| Tussilago |
14 |
kilómetros |
| Populus |
30 |
kilómetros |
| Senecio congestus |
200 |
kilómetros |
Un ejemplo claro de la dispersión por viento es la colonización de islas carentes de vegetación. Después de la erupción volcánica que destruyó la vida de Krakatoa, las plantas han iniciado la colonización; entre las principales se encuentran especies anemócoras. Resalta una de las pocas especies arbóreas de la familia de las compuestas, Vernonia arborea, la cual alcanza alturas hasta de 25 metros.
A través de la dispersión de semillas por animales se ha establecido una dependencia mutua entre las plantas y los dispersores. En muchos casos esto ha llevado a una coevolución entre ambos, por ejemplo, entre los frugívoros especialistas. En el caso de las aves, hay desde exclusivamente frugívoras (que solamente se alimentan de frutos) hasta frugívoras ocasionales, que son principalmente insectívoras y sólo de vez en cuando se alimentan de frutos.
Hay distintos grados de relación entre las plantas y sus dispersores y distintos mecanismos que a continuación se exponen:
a) Dispersión externa por animales (ectozoocoria). Muchas semillas o frutos tienen adaptaciones que les permiten adherirse al cuerpo de los animales. A este mecanismo de dispersión se le denomina ectozoocoria. Este tipo de semillas tiene apéndices en forma de ganchos o sustancias pegajosas con las que se adhieren al pelambre o a las plumas de los animales. A veces las semillas son transportadas en el lodo que se pega a las patas de las aves acuáticas y en estos casos por lo general no presentan ningún tipo de apéndice especializado. Darwin fue uno de los primeros investigadores en detectar este mecanismo de transporte. Colectó el lodo de las patas de una perdiz y lo mantuvo húmedo; obtuvo 82 plántulas. Posteriores investigadores han encontrado que son muchas las especies dispersadas por este mecanismo (Juncus, Carex, Cyperus, Alisma, etc.). Otros investigadores han colectado el lodo de la suela de zapatos, obteniendo un total de 43 especies, entre ellas varias ruderales, o sea especies que habitan principalmente en zonas perturbadas, como las orillas de caminos. Entre estas especies está la Plantago lanceolata, Urtica dioica, Bellis perennis y varias especies de pastos, siendo Poa annua una de las más comunes.
Las lorantáceas son plantas parásitas que viven sobre los árboles. Las raíces de estas especies se adhieren a las ramas del hospedero y se alimentan de las sustancias producidas por la planta a la que parasitan. Frecuentemente están cubiertas por una sustancia pegajosa que les permite adherirse al plumaje de aves. Cuando los pájaros descansan en las ramas, se limpian las plumas con el pico o se frotan contra la corteza, depositando ahí las semillas. De esta manera, la dispersión es eficiente y la planta se asegura de que por lo menos parte de sus semillas quedarán sobre las ramas de árboles hospederos. La eficiencia de la dispersión dependerá de la distancia a la que el ave transporte la semilla. Algunos ejemplos de estas semillas son Arceuthobium y Phoradendron.
Uno de los casos más llamativos es el de las semillas de Pisonia, árbol tropical de la familia de las nictagináceas, cuyas semillas son tan pegajosas que pueden llegar a cubrir casi por completo a un ave, afectando su capacidad de movimiento. Entre los animales que transportan semillas en el lodo de las patas están las aves, el ganado y los animales que se acercan a charcas de agua y se embarran el pelo de lodo. El hombre también desempeña este papel y transporta a las semillas tanto a cortas como a largas distancias. En general, este mecanismo de transporte es poco específico en relación con qué especie vegetal es acarreada por qué especie animal.
Como ya se mencionó, numerosas especies tienen ganchos o sustancias pegajosas que les permiten adherirse a los animales y en muchas ocasiones esto hace que la relación planta-animal sea más específica. Trabajos hechos en África, en particular en los matorrales de Kenia, demostraron que una especie de liebre (Lepus capensis) tenía hasta diecisiete especies de semillas adheridas al pelo. Estos investigadores realizaron un interesante experimento en donde compararon qué tan comunes eran estas especies en la vegetación y no encontraron coincidencia; es más, descubrieron que algunas de las especies comunes en la vegetación no estaban presentes en el pelaje. La conclusión a la que llegaron fue que algunos tipos de apéndices se adhieren más fácilmente a ciertos tipos de pelaje, y de ahí el grado de especificidad entre el vector animal y la planta (Figura IX.3 (A y B)). La altura de la planta también es un factor importante.
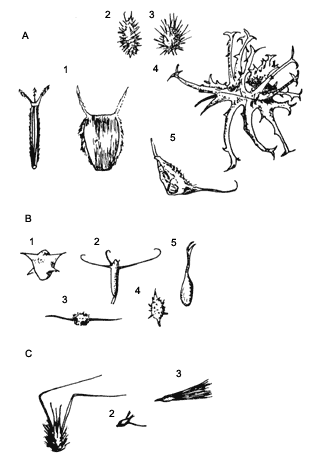
Figura IX.3. A) Semillas con ganchos que se adhieren a la ropa así como
al pelaje de los animales. Bidens 1, Xanthium 2, Cenchrus 3,
Harpagophytum 4, Proboscidea 5. B) Ganchos que les permiten anclarse
al sustrato: Trapanatans 1, Trapella sinensis 2, Blyxa echinosperma
3, Ceratophyllum demersum 4, Zoostera marina 5 (tomado de
Van der Pijl, 1976). C) Estructuras que les permiten desplazarse y modificar
su posicion una vez sobre el suelo. 1 Centaurea cyanus, 2 Trifolium
squarrosum, 3 Aegilops ovata.
En el chaparral mediterráneo, que es un tipo de matorral que se establece en climas secos, se encontró que 55 especies de plantas se adherían a los borregos y sólo 24 a las cabras. Para saber qué tanto tiempo permanecían adheridas, se pintaron semillas y se adhirieron artificialmente a los animales. Después de seguir la investigación durante varios días se encontró que los borregos retenían más semillas y por más tiempo que las cabras. La mayoría de las semillas se caía después de ocho días, pero las semillas de una especie (Medicago minima) se mantuvieron sobre los borregos hasta dos meses.
b) Dispersión por hormigas (mirmecocoria). Las hormigas son un grupo importante de dispersores ya que colectan semillas y las llevan a sus nidos. Constituye un tipo de ectozoocoria, pero es tan especializado debido a la relación que se establece entre la hormiga y la planta, que los botánicos lo han tratado por separado, dándole un nombre específico. Hay muchas especies de plantas que aprovechan esta actividad de las hormigas para ser transportadas a un sitio adecuado para germinar. Este método de dispersión ha sido registrado en gran cantidad de especies tanto en zonas templadas como tropicales. Para tener una idea de cuán común es este fenómeno, basta saber que en los brezales de Australia más de 1 500 especies de plantas son dispersadas por hormigas.
Frecuentemente, las semillas tienen estructuras especiales para atraer a las hormigas. El elaiosoma es una protuberancia de tejido nutritivo, con un importante contenido de aceites (y a veces almidones), que atrae a las hormigas por ser una fuente de alimento. Cuando la hormiga encuentra la semilla la acarrea al nido, remueve el elaiosoma y abandona la semilla en una vieja galería o en un montón donde se acumulan los desperdicios. Por lo general estos ambientes son adecuados para el establecimiento de plántulas, ya que poseen más materia orgánica y nutrientes que el suelo de los alrededores. La plántula que inicia su crecimiento ahí tiene ventaja sobre otras plántulas, ya que tiene acceso a una mayor cantidad de elementos nutritivos que le permiten crecer rápidamente.
Para algunas especies vegetales, el único mecanismo de dispersión de semillas es el que llevan a cabo las hormigas. Para otras, las hormigas constituyen la segunda fase de la dispersión; la primera muchas veces se da por explosión del fruto, que esparce las semillas a cierta distancia de la planta progenitora. Éste es el caso del género Viola y Dendromecon, en el cual las semillas son arrojadas a una distancia de 1 a 5 m de la planta progenitora; muchas de estas semillas son localizadas por las hormigas, que las acarrean a sus nidos. Esta combinación de mecanismos permite a las semillas escapar en el espacio (explosión del fruto en un momento dado) y en el tiempo (dispersión por hormigas a lo largo de varios días).
Para que las hormigas puedan transportar las semillas, éstas no deben sobrepasar cierto tamaño; algunas formas ayudan más que otras. La elaboración del elaiosoma no implica un gran costo energético para la planta. Esta estructura, según la especie, tiene orígenes muy diversos: a veces es un engrosamiento del funículo o de la carúncula, otras veces es una parte específica del endospermo, del fruto, etc. Lo interesante es que, por medio de caminos muy variados, diversos grupos de plantas han elaborado estructuras atrayentes para las hormigas, usándolas como su principal mecanismo de dispersión de semillas.
La semilla es doblemente beneficiada cuando es transportada por las hormigas. En primer lugar, evita la depredación por otros animales y en segundo, las semillas quedan en sitios adecuados para germinar, aunque sólo sean acarreadas unos cuantos metros.
En la naturaleza existen relaciones asombrosas entre las plantas y las hormigas. Como ejemplo está el caso del arbusto parásito Pachycentria tuberosa que crece y se alimenta de otra planta, Hydnophytum formicarium. Las hormigas viven dentro de los tallos engrosados de esta última, y colectan semillas del arbusto parásito, transportándolas a sus nidos dentro de los tallos. Aquí las semillas germinan y las raíces se ramifican entre un depósito y otro, y las hojas emergen por los agujeros que presentan las ramas del hospedero.
Otro ejemplo es el de las hormigas que forman sus nidos en los árboles y que establecen los llamados "jardines de hormigas", plantando las semillas del árbol hospedero en sus nidos, los cuales parecen ser buenos sitios para la germinación y el establecimiento.
Entre las especies cuyas semillas son dispersadas por hormigas están: Turnera ulmifolia, Datura discolor, Dischidia gaudichaudii, Calathea spp., etcétera.
Hay hormigas que más que dispersoras son depredadoras, ya que recogen muy diversos tipos de materiales, almacenan aquellos que son comestibles y los ingieren después de que se han fermentado. Frecuentemente acarrean cualquier tipo de semilla, la cual es abandonada con los otros materiales, pero no es capaz de germinar en esas condiciones (capítulo X).
c) Dispersión interna por animales (endozoocoria). Numerosas semillas están adaptadas para ser transportadas por animales, pero en forma interna. Frecuentemente tienen una cubierta llamativa, nutritiva o tienen algún material comestible adherido que atrae a los animales y les sirve como recompensa. Son capaces de tolerar el paso por el sistema bucal (trituración) y por el tracto digestivo (tratamiento químico) de sus dispersores. Las semillas están diferentes lapsos de tiempo dentro del organismo. El menor tiempo se da cuando son regurgitadas, ya que entonces ni siquiera atraviesan el tracto digestivo de las aves. También hay frutos con sustancias químicas que aceleran su paso por los intestinos. O bien, simplemente es posible que las semillas tengan suficiente protección, por ejemplo una testa dura e impermeable, que las proteja durante toda su estancia en este ambiente tan ajeno y destructivo. Mientras más tiempo permanezcan dentro, la dispersión de las semillas será a más distancia, pero también será mayor la probabilidad de que sufran daños por estar en contacto con los ácidos del estómago. La dispersión será en pequeños grupos de unas cuantas semillas. En un experimento se alimentó un tapir con semillas de la leguminosa Enterolobium cyclocarpum (guanacaste o parota). Durante un periodo de 23 días el animal estuvo defecando semillas tanto intactas como dañadas; sin embargo, sólo 22% de las que ingirió no sufrieron daños.
En el caso de algunos pastos y otras semillas de tamaño pequeño, el follaje de la planta constituye el "atractivo" para los animales. Al comerse las hojas, ingieren frutos y semillas que luego son dispersados con las heces.
Todo este proceso mencionado en los párrafos anteriores significa una serie de costos para la planta: tejidos ricos en nutrientes, tamaño más bien grande, testas duras, sustancias químicas, producción de gran número de frutos para asegurarse de que algunos sobrevivan a la depredación, etc. A pesar de ello, gran cantidad de especies presentan frutos que son ingeridos por animales y algunos de ellos son dispersados, lo cual hace pensar que los beneficios de este tipo de dispersión son mayores que los costos.
En los ejemplos que se presentan a continuación se aprecia cómo los animales ingieren y dispersan los frutos de una manera más direccional. Esto significa que debido a los hábitos y comportamientos particulares de los animales, hay más seguridad en cuanto al tipo de ambiente en el cual va a ser depositada la semilla (por ejemplo, dentro de la misma comunidad, o a la sombra de un árbol, o enterrada) elevando así la probabilidad de sobrevivencia de la nueva plántula.
Las semillas dispersadas de esta manera generalmente son depositadas en concentraciones altas. Esto es característico de los grandes herbívoros: se encontró que en heces fecales de elefante, las cuales pesaban 8 kilos, había 12 000 semillas de la leguminosa Acacia tortilis. Esto explica por qué es tan frecuente en muchas especies que varias semillas germinen juntas.
Entre los principales dispersores animales están ciertos mamíferos como murciélagos, caballos, monos y emúcs, numerosas especies de aves, tortugas, algunos peces y también las lombrices.
No todos los animales que ingieren frutos o semillas son agentes dispersores. Muchas veces es difícil distinguir si un animal es un dispersor o un depredador porque puede desempeñar ambos papeles. Sólo se considera que un animal es un dispersor si las semillas salen del tracto digestivo del animal sin haber sufrido daños. Si mantienen su capacidad germinativa, el animal que las ingirió es realmente un agente dispersor. El análisis de heces fecales de mandriles en Ghana demostró que 59 especies pudieron atravesar el tracto digestivo y germinar. El análisis del guano de aves en Australia reveló un total de 70 especies.
Regresando al doble papel de dispersor-depredador, existen dos mecanismos para cumplir esta función. En ambos casos, sólo unas cuantas semillas logran escapar, pero recordemos que únicamente se necesita que una logre establecerse y llegue al estado adulto para que remplace a la planta progenitora:
1) Los animales que se comen y digieren la mayoría de las semillas pero algunas logran no ser dañadas. Los siguientes datos reflejan la importancia de las pocas semillas que logran sobrevivir. En África se alimentó a herbívoros (impalas) con frutos de Acacia. En el caso de esta especie la pared de las semillas frecuentemente es digerida junto con la vaina que forma el fruto. Se encontró que sólo 4.8% de las semillas sobrevivieron el paso por el tracto digestivo. Sin embargo, conociendo la densidad de ambas especies, se calculó que esta tasa baja de sobrevivencia representaba más de 1 000 semillas intactas dispersadas diariamente por kilómetro cuadrado, durante los dos o tres meses que la especie está fructificando.
2) Los animales que almacenan semillas pero dejan algunas abandonadas. Ejemplo de este caso es un cuervo (scatterhoarding jays) que entierra bellotas en la tierra para comérselas más adelante. En un censo realizado sobre las bellotas producidas en un año, se encontró que 54% de la producción había sido tomado por estas aves. Se calculó que cada ave entierra 4 600 bellotas al año y que una proporción de éstas no es comida debido al exceso de bellotas almacenadas, a la muerte del ave o simplemente por olvido. Lo mismo se ha visto que sucede con roedores o ardillas. Las semillas son enterradas superficialmente, con frecuencia en sitios adecuados para germinar, donde las plántulas de encinos se establecen. Se ha visto que por lo menos la mitad de las plántulas que se producen en un año nace de las semillas enterradas por estas aves.
He aquí algunos ejemplos de animales que ingieren semillas y las dispersan, y que difieren tanto en los hábitos y comunidades en las que viven, como en su taxonomía:
Lombrices. Estos pequeños invertebrados ingieren semillas pequeñas, las cuales defecan bajo tierra; de esta manera contribuyen a su distribución y enterramiento en el suelo. En su interior se han encontrado semillas de diversas especies, lo cual prueba que las comen. En Europa hay registros de plántulas de orquídeas que han germinado en túneles de las lombrices.
Peces. Existen muchas especies de peces cuyo alimento principal son partes vegetales, entre ellas frutos y semillas. Existen casos en que las semillas se usan como carnada, como por ejemplo la leguminosa Pithecellobium microcarpum en Borneo.
Los peces ingieren semillas de plantas que viven a la orilla de ríos y pantanos (el higo Ficus, la leguminosa Inga, las palmas Arecastrum y Geonoma, la anona Guatteria). Estudios realizados en peces del Amazonas demostraron que varios de ellos ingerían regularmente en su dieta frutos y semillas. Se registraron 33 especies vegetales en sus estómagos; de éstas, 16 germinaron.
Reptiles. Entre los reptiles actuales sólo las tortugas y algunas lagartijas tienen hábitos vegetarianos. Existen algunos ejemplos bien documentados sobre la dispersión de semillas por reptiles, como el de una variedad local de tomate (Lycopersicon esculentum var. minor) en las Galápagos, el cual sólo germina después de atravesar el tracto digestivo de las tortugas. Las semillas tardan entre doce y veinte días para ser defecadas y las que tardan menos tiempo presentan menor germinación. En estas mismas islas las tortugas ayudan a la dispersión de las semillas de cactos.
Varias especies del género Gopherus comen y dispersan semillas de Asimina y del cacto Opuntia a lo largo del sureste de EUA y noreste de México.
Aves. Las aves se encuentran entre los principales animales que dispersan frutos y semillas. Por lo general lo hacen de manera accidental, cuando realizan sus actividades alimenticias (depredación). Por ejemplo, cuando toman los frutos de la rama para comérselos, dejan caer algunas semillas; o también cuando transportan semillas a los nidos para almacenar una parte del alimento que comerán posteriormente y ya no vuelven por él. Los roedores igualmente ayudan a bajar al suelo estas semillas de los almacenes arbóreos, donde a su vez, cuando son olvidadas, pueden germinar. Estos casos parecen totalmente accidentales, pero cuando ocurren con suficiente regularidad para asegurar la dispersión y sobrevivencia de un pequeño número de semillas constituyen un mecanismo de dispersión. Uno de los ejemplos más conocidos es el de las aves del género Nucifraga, que se alimentan de nueces de Fagus, Quercus, Juglans, Castanea, entre otras. Se ha visto a diversas especies de charas (Garrulus) enterrar nueces y bellotas. Experimentos hechos en Europa demostraron que un ave transporta en promedio 4 600 bellotas en una estación, algunas de ellas a distancias hasta de 4 kilómetros.
Muchas aves ingieren semillas de manera casual junto con el follaje y para muchas especies vegetales esto se ha convertido en un mecanismo de dispersión. Esto es especialmente importante para plantas de la familia de las ciperáceas, como Nymphaea, Nuphar y Pontederia, lo cual les permite dispersarse a grandes distancias.
Las semillas ornitócoras son aquellas que están realmente adaptadas para ser ingeridas por aves sin sufrir daños.
En general, los frutos pequeños más carnosos están adaptados a la dispersión por aves. La relación entre las plantas y las aves brinda ventajas a ambas partes y se ha desarrollado a través de miles de años. Por un lado, la planta invierte recursos en rodear a la semilla de una cubierta carnosa y nutritiva, atractiva para las aves y a cambio de ello el ave ingiere gran cantidad de estos frutos, digiere la pulpa y arroja la semilla a cierta distancia de la planta progenitora, ayudando de esta manera a su dispersión. Se puede decir que es un caso de coevolución, pues tanto las plantas como las aves —por lo menos las frugívoras más especializadas— se han modificado mutuamente durante el curso de su evolución. En este sentido, el tamaño, forma y contenido nutritivo de los frutos de distintas plantas hace que estos sean conspicuos, atractivos y físicamente accesibles para ciertos tipos de aves. Las semillas deben estar protegidas de modo que sean capaces de resistir las situaciones que atraviesan antes de ser regurgitadas o defecadas. A su vez, los picos de las aves frugívoras, su sistema digestivo y otras características, se han adaptado para detectar, cortar, ingerir y digerir eficientemente los frutos.
Las aves dispersoras de semillas desempeñan un papel más importante en las comunidades tropicales que en las templadas. En estas últimas sólo hay frutos durante una parte del año, por lo que las aves tienen que tener dietas combinadas que les permitan obtener alimento y así subsistir todo el año. En cambio, en las zonas tropicales hay frutos durante todo el año, lo cual se debe a las especies que fructifican todo el tiempo, y a la distribución de las especies en fructificación, a lo largo de todos los meses. Esto permite que haya aves exclusivamente frugívoras.
Dentro de las aves frugívoras hay toda una gama que abarca desde las aves especialistas, las cuales dependen básicamente de los frutos para obtener proteínas, lípidos, carbohidratos y agua, hasta las aves oportunistas que sólo usan los frutos para obtener agua y carbohidratos y que tienen otras fuentes que les proporcionan lípidos y proteínas. Esto es un reflejo de lo que algunos autores llaman la calidad de la dispersión. Los especialistas son dispersores de alta calidad pues dependen de frutos particulares, lo que asegura que estarán presentes durante la fructificación y que ingerirán los frutos, además de que su sistema digestivo especializado les permite regurgitar gran cantidad de semillas sin que éstas sufran daños. El costo que tiene que pagar la planta por esta dispersión de alta calidad es la producción de una pulpa muy nutritiva. Ejemplos notables de ello son las familias de las palmas, las lauráceas y las burseráceas.
En cambio, las plantas cuyos frutos sólo son utilizados de manera oportunista por un alto rango de frugívoros casuales, brindan mucha menor recompensa a las aves. La planta equilibra la dispersión de baja calidad con la producción de un mayor número de semillas de menor tamaño, asegurando así que algunas de ellas sobrevivan a la depredación y sean dispersadas.
Los frutos también son alimentos importantes para muchas aves que migran entre zonas templadas y tropicales. Aves como el vireo ojirrojo norteño (Vireo olivaceus), el chipe peregrino (Vermivora peregrina), el zorzalito de Swainson (Catharus ustulatus) y el papamoscas copetón viajero (Myiarchus crinitus) habitan en el norte de América y durante el invierno migran al trópico. Se ha visto que entre estas cuatro especies se consume el 60% de las semillas del árbol tropical Guarea glabra. Curiosamente, muchas de las aves migratorias son insectívoras en sus zonas de origen y cuando migran al trópico se tornan frugívoras: el tirano dorsinegro (Tyrannus tyrannus) consume gran cantidad de frutos de Didymopanax morototoni cuando viaja de Sudamérica hacia el norte. Durante estos viajes las semillas son dispersadas a mayores distancias que con las aves residentes.
Existe una correlación entre las épocas de fructificación de las plantas de algunas zonas geográficas y la época del paso de las aves migratorias. Así, en el este de Norteamérica los frutos carnosos maduran entre agosto y principios de octubre. Las aves migratorias cruzan esta zona entre fines de agosto y mediados de octubre, coincidiendo con la época de mayor fructificación de especies ornitócoras. Algunos ejemplos son el chipe grupidorado gorjiblanco (Dendroica cioronata), que se alimenta y dispersa extensamente las bayas de la hiedra venenosa Rhus rhadicans y el zorzalito colirrufo (Catharus guttatus) que ingiere frutos de varias especies de Smilax.
Los frutos que requieren de aves para su dispersión han desarrollado una serie de características que los distinguen como semillas con síndrome ornitócoro. El desarrollo de estas características les ha permitido establecer una estrecha relación con las aves en la cual los frutos y semillas atraen al ave y le brindan alguna recompensa en forma de alimento, por lo cual el ave los busca con asiduidad y participa activamente en su dispersión. Las aves han desarrollado muy poco el sentido del olfato y utilizan principalmente la vista, por lo que las características que las semillas ornitócoras presentan son:
—colores atractivos cuando maduran;
—una parte comestible atractiva;
—una protección externa (verde o ácida) que evita que sea ingerida cuando
aún está inmadura;
—una protección interna que evita que la semilla sea digerida;
—las semillas no están encerradas dentro de un fruto duro y cuando lo
están, cuelgan o están expuestas.
Los colores más frecuentes son el rojo, el azul y el negro. Esta coloración conspicua aparece sola o en combinación. Los colores, además de atraer al ave, indican la madurez del fruto y por tanto son señal de que la semilla ha completado su desarrollo y que ya está lista para ser dispersada. Por lo general, la maduración no se da masivamente al mismo tiempo, y es por esto que la planta muestra una gradación de colores hasta el final de la estación, cuando todos los frutos ya están maduros. Se ha visto que las combinaciones de colores, por ejemplo rojo y negro, son más atractivas que la presencia de un solo color, por lo que las semillas son removidas más rápidamente. Muchas veces la presencia de dos colores se debe al contraste entre el fruto o la semilla y una parte accesoria de la planta, como podría ser un pedicelo, una bráctea, una cápsula, o simplemente las hojas. Se desconoce la importancia de ciertos tonos y colores y también se ignora qué ha favorecido la presencia de unos sobre otros.
Algunas semillas no comestibles presentan colores parecidos a los de otras especies que sí lo son. A veces las aves se confunden y las ingieren. No se sabe si esto promueve la germinación. Este mimetismo en semillas es frecuente sobre todo en la familia de las leguminosas: Abrus precatorius, Adenanthera pavonina, Cassia costata, Pithecellobium clypearia, P. lusorium, Erythrina spp., etcétera.
El tamaño de los frutos ingeridos por aves varía, pero no es muy grande. Se considera que el límite está en frutos de unos 4 X 7 cm, los cuales sólo pueden ser ingeridos por aves grandes, como los tucanes.
Las aves frugívoras emplean tres mecanismos para dispersar las semillas. El primero consiste en expulsar a la semilla del fruto cuando el ave se está comiendo la pulpa. En estos casos, las semillas caen muy cerca del árbol que produjo los frutos. En el segundo caso el ave se traga el fruto entero, incluyendo las semillas. Éstas pueden tener dos destinos: ser regurgitadas o atravesar el tracto digestivo y ser defecadas. En el primer caso las semillas son depositadas cerca de la planta progenitora pues el proceso es mucho más rápido, a veces de sólo unos cuantos minutos. Cuando las semillas son defecadas, ya ha transcurrido un lapso mayor (de una a varias horas) y por lo tanto el ave se encuentra más lejos de la fuente de frutos.
Como ejemplo de la actividad de las aves veamos lo que sucede con una mora silvestre (Rubus fruticosus), muy abundante en Inglaterra (Snow y Snow, 1988). Florece en junio y los frutos maduran entre agosto y octubre. Los frutos carnosos son pequeños, entre 12 y 15 mm. Se registraron ocho especies de aves dispersoras que varían mucho en importancia. El tordo, Turdus merula, realizó 138 visitas (121 de ellas en septiembre); el estornino, Sturnus vulgaris, ocupó el primer lugar pues realizó 335 visitas (227 en octubre y 108 en noviembre). Las otras seis especies de aves sólo hicieron 42 visitas entre todas. Estos datos muestran un conjunto diferente de aves que hace uso de un recurso y que además lo divide en el tiempo. Sólo se observaron tres aves depredadoras.
Mamíferos. (ungulados, primates, roedores, etc.). Al igual que en el caso de las aves, la dispersión por mamíferos es un fenómeno más común en los trópicos que en las zonas templadas. Entre los principales mamíferos frugívoros están los roedores, los murciélagos, los changos, los osos y los ungulados.
De nuevo, al igual que en el caso de las aves, muchos roedores son depredadores que de manera accidental dispersan las semillas. Ardillas, ratas y hámsters se alimentan de las semillas de encinos, pinos y cereales, pero también las transportan y almacenan, donde en ocasiones las olvidan y éstas germinan. Esto se ha documentado con detalle en lo que se refiere a las ratas canguro (Dipodomys), que se alimentan de las semillas de mezquite (Prosopis) y las almacenan, contribuyendo a su regeneración. En bosques templados las ardillas (Sciurus) las entierran en forma aislada o en pequeños grupos; estas semillas son de nogales (Juglans y Carya), encinos (Quercus), castaño (Castanea) y arce (Acer). Algunas de estas semillas logran germinar y establecerse.
Muchos animales, cuando encuentran un fruto comestible lo toman y transportan a otra zona, donde se lo comen. Durante este proceso pueden caerse algunas semillas que de esta manera son dispersadas sin ser depredadas. Esto ocurre accidentalmente, como parece suceder con los conos de cicadas, los cuales son acarreados por mandriles. Pero en otras ocasiones, el fruto tiene un diseño que desanima a los posibles predadores-dispersores. Los frutos de Sterculia apetala tienen pequeños pelos duros que se clavan en el hocico y patas del animal conforme éste va abriendo el fruto y tomando las semillas. Al cabo de un rato el animal deja el fruto y se dedica a quitarse los pelos incrustados. De esta manera, algunas semillas son acarreadas sin ser depredadas.
Otro ejemplo es el de los roedores que se alimentan principalmente de las partes carnosas, sin causar así daños a la semilla. Éste es el caso del fruto del chocolate, Theobroma cacao, del cual los roedores comen sólo la pulpa dulce que rodea a las semillas.
Roedores grandes como los agutíes también son dispersores (y depredadores) importantes en zonas tropicales. Por ejemplo, son capaces de romper las cápsulas leñosas de las nueces del Brasil (Bertholletia excelsa), comerse la pulpa interior y luego enterrar las semillas en el suelo como reserva. Lo mismo sucede con las semillas de algunas palmas como Attalea funifera.
Muchos mamíferos herbívoros ingieren semillas de manera accidental cuando se están alimentando del follaje de plantas. Esto sucede con especies de amarantáceas, quenopodiáceas y algunas leguminosas, entre otras, y es tan frecuente que se pensó que la especie Chenopodium album había sido usada como alimento por grupos humanos desde la prehistoria, debido a la gran cantidad de semillas encontradas cerca de habitaciones humanas. Posteriormente se demostró que la mayoría de las semillas estaban depositadas en capas de restos de excremento animal.
Entre las características que exhiben las semillas dispersadas por mamíferos están:
—mayor protección de la semilla contra daños mecánicos, ya que los mamíferos
tienen dientes y mastican con gran fuerza; esta protección a veces va acompañada
de sustancias tóxicas o de sabor desagradable;
—olor atrayente;
—tamaño más o menos grande.
Al igual que en el caso de las aves, mientras más tiempo pasen las semillas dentro del animal, mayor será la distancia a que serán dispersadas. Semillas de guanacaste (Enterolobium cyclocarpum) ofrecidas a un tapir fueron defecadas en pequeñas cantidades durante un lapso de 23 días, aunque ello aumenta el riesgo de que sufran daños. Del total ingerido, sólo 22% permaneció intacto. Para muchos animales, mientras más grande sea la semilla, mayor es el periodo de retención dentro del tracto digestivo.
En zonas tropicales los ungulados (rumiantes y elefantes) son dispersores importantes de semillas. En África existen numerosos ejemplos, debido a que en esta parte del mundo hay enormes extensiones de pastizales que alimentan a cuantiosas manadas de herbívoros grandes. Los elefantes, uno de estos herbívoros, se alimenta de hojas y frutos y se ha visto a varias especies de plantas germinar en sus excrementos: Dumoria heckeli, la palma Borassus flabellifer, Hyphaene, Adansonia, Durio, Mangifera, etcétera.
Existe una relación interesante entre las leguminosas y los rumiantes. Estos últimos no sólo se alimentan de pastos sino también de frutos que constituyen una parte esencial de su dieta alimenticia. Digieren las vainas leñosas de muchas de las leguminosas de la sabana, como es el caso de la relación entre los antílopes y los frutos de Adansonia. Entre los árboles más comunes en la sabana están los pertenecientes al género Acacia. Estas especies se han especializado en usar a los grandes herbívoros como mecanismo de dispersión; sus frutos son vainas nutritivas, duras, que en ocasiones permanecen en los árboles o bien caen al suelo. Las semillas son extremadamente duras y muy lisas, lo cual dificulta que sean trituradas por los molares del animal. Entre los géneros cuyas especies son dispersadas por rumiantes están: Cassia, Acacia, Prosopis, Ceratonia, etcétera.
Un investigador estadunidense, Daniel Janzen, que trabaja en las selvas y pastizales de Costa Rica ha planteado que muchas de las especies que actualmente se encuentran en el trópico de América están adaptadas a la dispersión por grandes herbívoros, al igual que en África. Estudios paleontológicos de depósitos de hace 10 millones de años muestran que esta fauna de grandes animales era mucho más numerosa que la actual, y que abarcaba organismos parecidos a los grandes proboscidios actuales, gliptodontes, équidos, osos y armadillos gigantes hoy extintos. Calcula que aproximadamente quince géneros (y por tanto gran cantidad de especies) de estos grandes herbívoros desaparecieron hace alrededor de 10 000 años. Ello produjo cambios en la distribución de muchas plantas, ya que dejaron de contar con dispersores eficientes, lo cual probablemente ocasionó una reducción de sus poblaciones.
La reintroducción de otros herbívoros de mediano tamaño como las vacas y los caballos ha remplazado en parte a los herbívoros desaparecidos. Cuando se les deja pastar libremente ingieren grandes cantidades de algunos frutos como los de Crescentia alata y los del guanacaste (Enterolobium cyclocarpum). El primero de ellos, que es el árbol productor de las jícaras, crece formando manchones aislados de 3 a 4 metros en los pastizales. Los frutos esféricos (6 a 15 cm de diámetro) contienen entre 200 y 800 semillas inmersas en una pulpa fibrosa. Los frutos caen al suelo sin abrirse, donde terminan de madurar. En este momento son relativamente dulces y la pulpa se vuelve negra y olorosa. Los caballos pueden romper la cubierta dura externa de los frutos con los incisivos y comerse la pulpa sin tener la necesidad de masticarla demasiado. Para tener una idea de la capacidad de dispersión que los caballos representan para esta especie, el mencionado autor da los siguientes datos: un grupo de 17 caballos se comió 666 frutos en un periodo de 24 horas.
Se desconoce el porcentaje de semillas que permanece viable durante el paso por el tracto digestivo de los animales, pero el excremento está lleno de semillas viables que germinan a partir del segundo día después de que el caballo come los frutos. Un 97% de las semillas extraídas del excremento son capaces de germinar. Observaciones hechas en el Parque Nacional de Santa Rosa, Costa Rica, demuestran la presencia de gran cantidad de plántulas e individuos jóvenes del árbol de Crescentia en aquellos lugares donde se permite pastar a los caballos; en cambio, están ausentes de las áreas en que estos herbívoros han sido excluidos.
Actualmente, diversas especies de changos y monos se alimentan de frutos, aunque en general son muy destructivos ya que se comen todo; sin embargo, algunos de ellos ayudan en la dispersión de semillas.
Otros mamíferos importantes son los tapires. En la zona del Amazonas entierran semillas de Araucaria angustifolia, muchas de las cuales después germinan. Gatos del género Paradoxurus suben a los árboles y consumen distintos tipos de frutos y semillas. Un fruto muy apetecido por distintos mamíferos es el de Durio zibethinus. El fruto es grande y espinoso. Las semillas no tienen ninguna protección, pero cuando el fruto está verde, son tóxicas. Se necesita cierta fuerza y habilidad para abrir el fruto y liberar la semilla junto con una especie de arilo con alto contenido de aceites. Entre los animales que gustan de este arilo están los orangutanes, rinocerontes, tapires, osos, elefantes y ocasionalmente carnívoros como las panteras.
Murciélagos. Dentro de este grupo de mamíferos hay numerosas especies cuyo alimento principal son frutos. Generalmente no ingieren las semillas y sólo consumen la pulpa jugosa de los frutos; cuando las semillas son muy pequeñas y están inmersas dentro de la carne del fruto, lo ingieren todo y luego regurgitan o defecan las semillas, como en el caso del Ficus (higos), Piper y Cecropia. Los murciélagos son de hábitos nocturnos e incapaces de percibir colores; tienen un agudo sentido del olfato. La dispersión no la realizan a grandes distancias. Frecuentemente, después de ingerir algunos frutos van a su lugar de reposo, donde regurgitan los restos de alimento, incluyendo las semillas. Estas distancias son de unos pocos cientos de metros.
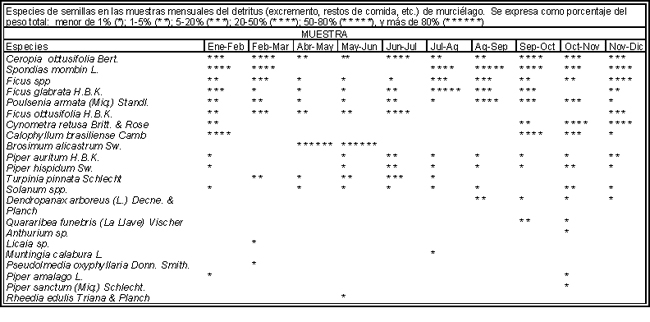
Figura IX.4. Especies dispersadas por murciélagos a lo largo del año en
los Tuxtlas, Veracruz. Hay especies que producen frutos todo el año, otras son
muy estacionales. La dieta de los murciélagos, y su actividad como dispersores
de semillas se produce en los doce meses. Los asteriscos indican el porcentaje
de cada especie del total de detritus recolectado: menos de 1% (*); 1- 5%(*
*); 5- 20% (* * *); 20- 50% (* * * *); 50- 80% (* * * * *), y más de 80% (*
* * * * *) (tomado de Vázquez-Yanez y colaboradores, 1975).
Trabajos realizados en Los Tuxtlas, Veracruz, por Carlos Vázquez-Yanes y sus colaboradores, demostraron que los murciélagos se alimentan de una gran cantidad de frutos de las plantas de la región (Figura IX.4), que incluyen tanto especies propias de selvas como de acahuales, o sea de vegetación que se establece cuando la selva se tira y se deja regenerar. Muchas de estas especies forman parte del banco de semillas (capítulo VII) y probablemente los murciélagos sean en parte responsables de su distribución a lo largo de la selva y de los acahuales. En la figura IX.4 es interesante notar cómo los murciélagos hacen uso de las especies que fructifican todo el año, pero también cómo son capaces de cambiar a especies estacionales como el ramón (Brosimum alicastrum), bajando en esa época su consumo de otras especies.
Los murciélagos varían de tamaño. Los más grandes, como los Pteropus, son capaces de transportar frutos de mango, aunque la mayoría son más pequeños, y por lo tanto el tamaño del fruto constituye una limitante. Se orientan con el sonar, por lo que se les dificulta cruzar follajes densos. Prefieren frutos que no estén inmersos dentro de la copa de los árboles.
Las características de los frutos preferidos por los murciélagos son:
—color poco llamativo;
—olor particular que recuerda el de sustancias fermentadas o rancias;
—expuestos fuera del follaje;
—partes jugosas poco protegidas;
—no demasiado pequeños.
La posición de los frutos en el árbol muchas veces les confiere una forma o estructura llamativa. Por ejemplo, Terminalia catappa tiene una ramificación en forma de estratos, en el extremo de los cuales cuelgan los frutos y se balancean en algunas especies de higueras (Ficus), Artocarpus, Lansium, etc. En otras especies los frutos salen directamente del tronco.
Entre las familias más visitadas por los murciélagos están: palmas, moráceas, crisobalanáceas, anonáceas, sapotáceas y anacardiáceas.
3) Dispersión por agua (hidrocoria)
El agua es un agente dispersor importante en plantas acuáticas y costeras, aunque algunas de estas especies también se dispersan mediante aves o peces. A continuación se describen algunos mecanismos que presentan las semillas y que les permiten utilizar el agua como agente de dispersión.
a) Dispersión por lluvia. Algunas semillas utilizan la lluvia para dispersarse. Las hay desde poco especializadas, que simplemente abren sus frutos y exponen las semillas de modo que cuando llueve, éstas caen del fruto y el agua de lluvia que escurre las aleja unos metros más. A veces los frutos están orientados horizontalmente, de modo que la superficie que recibe el impacto de las gotas de lluvia es mayor y las semillas son lanzadas a mayores distancias, como la Thlaspi alpestre (Crucífera), en la cual las semillas alcanzan una distancia de 80 centímetros.
b) Dispersión por corrientes. Las semillas que son dispersadas por
corrientes de agua tienen la necesidad de flotar, de modo que puedan recorrer
cierta distancia. Gunn y Dennis (1976) han encontrado diversas características
que les permiten a las semillas flotar tanto en agua dulce como marina y las
han conjuntado en cinco grupos (Figura IX.5):
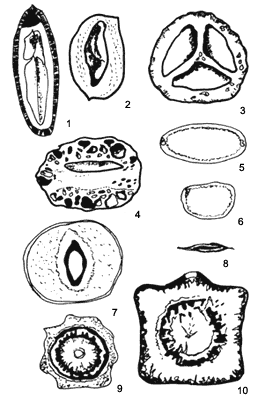
Figura IX.5. Semillas que se dispersan mediante las corrientes marinas, y que muestran diferentes mecanismos de flotación. Forman los grupos: Entada phaseoloides 1, Omphalea diandra 2, Canarium decumanum 3 y Sacaglottis amazonica 4. El segundo Dioclea reflexa (5 y 6); el tercero la Cerbera manghas 7, el cuarto Peltophorum inerme 8 y el quinto Grias cauliflora 9 y Barringtonia asiatica 10 (tomado de Gunn y Dennis, 1976).
Grupo 1. La capacidad de flotar se debe a una cavidad dentro de la semilla, que a veces existe entre los cotiledones (por ejemplo en el caso de varias especies de Entada e Ipomoea), o cuando el endospermo o el embrión no ocupan todo el espacio de la semilla (especies de Aleurites y de Caesalpinia). Cuando la unidad se dispersa en un fruto, puede haber cavidades o espacios dentro de la pared del fruto (especies de Juglans o nogal), o bien existir una cavidad central (en el caso de las palmas).
Grupo 2. Las semillas flotan debido a que el tejido de los cotiledones es muy ligero, como sucede en varias leguminosas de los géneros Erythrina (colorín), Dioclea (ojo de venado) y Canavalia (frijol de playa).
Grupo 3. Las semillas tienen una pared de tejido semejante al corcho, o bien de consistencia fibrosa; a veces aparecen ambas características juntas. Este patrón se presenta en las semillas de Terminalia (almendro) y de Hippomane mancinella (árbol de manzanilla).
Grupo 4. Las semillas son tan delgadas que son capaces de flotar, como en el caso del mangle blanco, Avicennia germinans.
Grupo 5. Una combinación de los factores arriba mencionados permite a las semillas flotar (Barringtonia asiatica, Cocos nucifera, coco).
b.1) Dispersión por corrientes de agua dulce. Muchas plantas acuáticas o que viven en las orillas, presentan semillas pesadas que requieren incrementar su superficie para poder flotar y ser transportadas pbr el agua. Algunas presentan pelos (Nymphoides orbiculata), otras una especie de arilo (Nymphaea alba). Frecuentemente, cuando se pudre la estructura que les permite flotar, las semillas caen al lodo, cesando así su viaje. A veces se ayudan de pequeños ganchos que les permiten mantenerse ancladas en el mismo sitio (Figura IX.3(b)).
Otras semillas flotan debido a que tienen un peso específico bajo, el cual logran por medio de espacios llenos de aire, cotiledones muy ligeros o tejidos especiales tipo corcho: Grias cauliflora, especies de Combretum y de Nelumbo.
b.2) Dispersión por corrientes marinas. La dispersión de plantas terrestres por corrientes marinas es un fénómeno muy complejo. Aunque muchas semillas llegan de nuevo a tierra firme, esto no garantiza que se vayan a establecer. Las condiciones en la orilla de la playa muchas veces no son adecuadas para la germinación y menos para el establecimiento de las plántulas.
Muchas semillas son arrastradas hasta la orilla del mar y después dispersadas por corrientes marinas. Algunas cuantas permanecen a flote, por lo que frecuentemente llegan a playas localizadas en lugares muy alejados de donde viven las plantas que las produjeron. A pesar de que se conservan en buen estado y que la testa permanece intacta, muchas de ellas no están viables. El embrión ya ha muerto y son incapaces de germinar. En estos casos, a pesar de que haya un flujo constante de semillas de una especie a otras playas, no germinarán y no habrá plántulas que sobrevivan para crear poblaciones permanentes en esas nuevas playas.
Existen autores, como Gunn y Dennis (1976), que han recorrido muchísimas playas buscando semillas arrojadas por el mar. Consideran que menos de 1% de las plantas tropicales producen semillas capaces de flotar, ya sea en agua dulce o marina por periodos de un mes.
A pesar de que en algunas playas es frecuente encontrar semillas, pocas veces éstas han sido observadas flotando en el mar, lejos de una costa. Muestras de plancton tomadas de la superficie del mar, no incluyen semillas. Estos mismos autores analizaron muestras de arena provenientes del fondo marino de zonas del Caribe, Golfo de Panamá y Golfo de Guinea, en África. Encontraron semillas de aproximadamente 20 especies de plantas tropicales. Entre las especies encontradas en estas muestras están la leguminosa (Pterocarpus officinalis), el coco (Cocos nucifera), el almendro (Terminalia catappa), el ciruelo silvestre (Spondias mombin), el mango (Mangifera indica), el mangle rojo (Rhizophora mangle), y el mangle blanco (Avicennia germinans). En general, el conjunto no es tan diferente del que se encuentra en las playas.
Las corrientes marinas son mecanismos de dispersión, pero también constituyen barreras infranqueables para la mayoría de las especies. Sin embargo, algunas han logrado adaptarse a este mecanismo de dispersión, lo cual les ha permitido llegar a regiones lejanas y tener una distribución muy amplia. Las semillas que se dispersan usando corrientes marinas dependen en primer lugar de la presencia de una corriente, en segundo lugar de la dirección del viento y finalmente de la topografía de la playa. Requieren también de una testa impermeable que proteja al embrión del agua marina. El tiempo que permanecen flotando puede ser bastante largo, por lo que necesitan estructuras y protecciones más complejas que las plantas que se dispersan por corrientes de agua dulce. Entre las plantas cuyas semillas son dispersadas por corrientes marinas están Caesalpinia bonduc, Cakile maritima, Canavalia rosea, Ipomoea pes-caprae, Hippomane mancinella, Cocos nucifera, Wedelia biflora, entre otras.
Dentro de este grupo se encuentran las especies en las que la propia planta es la encargada de la dispersión de las semillas y participa de manera más activa en el propio proceso. Los frutos de este grupo caen, explotan, y lanzan las semillas; se les denomina balócoras (ballo—aventar).
Existen frutos en los que el tejido cambia su tensión debido a cambios higroscópicos. Al disminuir la humedad en el entorno, los tejidos comienzan a perder agua, incrementándose la tensión en algunos puntos, hasta que se produce una ruptura violenta. Este mecanismo se presenta principalmente en hierbas y arbustos (es frecuente en leguminosas como Crotalaria incana y Chamaecrista chamaecristoides), pero también en algunos árboles como Hevea spp. y Hura crepitans.
A veces se requiere de un agente externo que procure la energía necesaria para que se lleve a cabo la ruptura y expulsión de las semillas. Frecuentemente es un animal que al pasar roza el fruto y hace que se abra. Otras veces es la lluvia y otras más es el viento.
En esta categoría se colocan las especies con semillas grandes y pesadas que caen por su propio peso. Sin embargo, es una categoría poco clara; se ha comprobado que muchas especies se consideraban barócoras debido a que no se conocía el mecanismo de dispersión. Ejemplos de especies que se consideran dentro de esta clase son Aesculus hippocastanum y Mora excelsa.
Una vez que las semillas han sido dispersadas y se han alejado de la planta progenitora, pueden ser dispersadas de nuevo por el mismo agente o por otro distinto. Esto les permite alejarse aún más, o bien incrementar la posibilidad de encontrar sitios adecuados para la germinación y establecimiento. Algunas semillas son totalmente pasivas, mientras que otras son más activas en este sentido. Así, hay semillas que presentan pelos o ganchos sensibles a cambios de humedad del ambiente, los cuales se mueven a través de un proceso de hidratación y deshidratación, desplazándose o enterrando parcialmente la semilla (Figura IX.3(c)). Por ejemplo, en las gramíneas de los géneros Stipa, Avena y Erodium se ha visto que este movimiento y anclaje o enterramiento favorece la germinación y el establecimiento exitoso de las plántulas.
En las dunas costeras existe una leguminosa endémica del Golfo de México (Chamaecrista chamaecristoides). Produce una vaina que explota y arroja las semillas alrededor del arbusto que las produjo. Durante la época de nortes, los vientos arrastran estas semillas y las distribuyen sobre la arena desnuda, sobre todo en las pendientes de barlovento. Cuando vienen las lluvias, germinan e inician la colonización y fijación en las arenas móviles.
ANCLAJE O FIJACIÓN DE LAS SEMILLAS AL SUELO
Algunas semillas tienen posibilidades de anclarse o fijarse al suelo y de esa manera evitar seguir siendo dispersadas. En ocasiones, las mismas estructuras que les ayudaron en la dispersión les permiten fijarse al suelo. Tal es el caso de algunas semillas que presentan pelos o ganchos, como el ejemplo descrito en el párrafo anterior. Es un fenómeno frecuente en epífitas plumosas así como en plantas acuáticas y de desiertos. Otras especies tienen semillas con una cubierta mucilaginosa, que al hacer contacto con el agua se vuelve gelatinosa, permitiendo así que las semillas se adhieran temporalmente al suelo.
Otro ejemplo más son las semillas enterradas por la propia planta, donde el fruto en desarrollo se encuentra en el extremo de un pedicelo que se alarga, dobla y entierra, como en el caso del cacahuate (Arachis) y de una planta que habita las playas del Golfo de México (Okenia hypogea). Este mecanismo se ha registrado principalmente en ambientes áridos, aunque no es exclusivo de ellos. Se podría considerar como un caso opuesto a la dispersión, pues la planta progenitora asegura que las nuevas plántulas crezcan cerca, en un ambiente adecuado debido a que ya hay un adulto reproductivo de esa especie.
Dentro de una comunidad vegetal como es el bosque, una selva o un desierto, generalmente hay un amplio rango de mecanismos de dispersión. Sin embargo, la proporción de especies que utilizan uno u otro mecanismo varía entre las distintas comunidades.
Entre la vegetación que coloniza un área abierta tiende a predominar la dispersión por viento (anemócora), ya que las semillas de una especie deben ser las primeras en llegar a un sitio abierto, libre de la competencia de otras plantas, si desean germinar y establecerse. En cambio, en bosques o selvas donde además existen numerosos animales, se incrementa notablemente la dispersión por aves y mamíferos y por tanto coexisten muy variados mecanismos de dispersión.
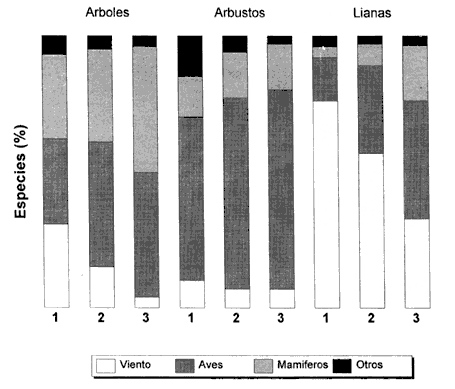
Figura IX.6. Porcentaje de semillas de árboles, arbustos y lianas dispersadas
por cada uno de los mecanismos de dispersión (viento, aves, mamíferos
y otros no identificados) en tres comunidades de selva y establecidas en ambientes
con diferente grado de humedad. La selva baja caducifolia (1) se localiza en
el parque Santa Rosa en Costa Rica, la selva mediana en Barro Colorado, Panamá
y la selva alta perennnifolia en Río Palenque, Ecuador (tomado de Gentry, 1982).
En la figura IX.6 se muestra el porcentaje de especies de árboles, arbustos y lianas dispersadas por el viento, por aves y por mamíferos en varios tipos de selva de Costa Rica y Panamá. En las lianas, es alta la dispersión de semillas que usan como agente el viento, seguida por la dispersión ornitócora (por aves) y la dispersión por mamíferos sólo se presenta en un porcentaje pequeño de especies. En los arbustos de los tres tipos de selva el porcentaje de especies dispersadas por viento es muy bajo, siendo el de aves el predominante. En el caso de los árboles predomina la dispersión por aves y mamíferos y es variable la de semillas dispersadas por el viento, aunque menor que en el caso de las lianas. Si se observan ahora los datos según el tipo de selva, se aprecia que la dispersión por viento es más importante en las selvas más secas. Estas selvas se denominan selvas bajas caducifolias y se establecen en ambientes donde la precipitación es estacional, con una clara época de secas. Son selvas cuyos árboles alcanzan alturas de 10 a 15 m y que debido a la estacionalidad pierden sus hojas durante la época de secas; así mismo, las plantas herbáceas mueren para germinar con las siguientes lluvias. En estos casos, muchas de las especies fructifican durante la época de secas, la cual coincide con los vientos. Los árboles no tienen follaje, lo cual permite que las semillas sean fácilmente transportadas y depositadas por las corrientes de aire. En el otro extremo está la selva siempre verde, también llamada selva alta perennifolia, la cual se establece en condiciones de mucha mayor humedad. En estos casos la estacionalidad es mucho menos marcada. Abundan las aves y mamíferos durante todo el año y las plantas los utilizan para dispersarse.
DISPERSIÓN A GRANDES DISTANCIAS
Un aspecto muy atrayente de la dispersión es la colonización de islas. La flora y fauna actual en gran cantidad de islas procede de la que existe en los continentes, y sólo algunos ejemplares logran llegar y colonizar exitosamente los nuevos territorios. En las islas surgidas recientemente (por erupciones volcánicas o emergencia de atolones arrecifales) existen laboratorios donde se estudia la colonización y por tanto la eficiencia de los distintos mecanismos de dispersión de los diversos grupos de organismos, tanto animales como vegetales.
A las islas constantemente arriban organismos, pero igual que llegan, desaparecen. Sin embargo, unos cuantos logran establecerse y aunque esas llegadas sean poco frecuentes, el resultado es exitoso. Se ha calculado que la actual flora de Galápagos pudiera ser el resultado de una llegada y colonización exitosa cada 7 900 años; para las islas de Hawai se ha calculado en 20 000 a 30 000 años.
Las islas son pequeños puntos de tierra (ambientes favorables para plantas y animales terrestres) rodeados de inmensas extensiones de agua que constituyen barreras físicas para los que quieren llegar a ellas. Algunas especies se diseminan por el agua y otras por el aire, pero en ambos casos son pocas las posibilidades de que lleguen a tierra. Por tanto, la probabilidad de que las semillas lleguen a una isla depende de varios aspectos:
—la distancia a la fuente más cercana de semillas;
—su posición en relación con las rutas de migración de aves;
—dirección de los vientos dominantes; frecuencia de huracanes;
—tamaño de la isla.
La colonización exitosa dependerá del medio físico que presente la isla y de lo adecuado que sea para que la semilla germine y la plántula se establezca y desarrolle.
Carlquist, en una interesante serie de trabajos analizó la dispersión a largas distancias. En uno de ellos comparó el probable mecanismo de dispersión de la flora presente en una serie de islas localizadas en distintos puntos del Pacífico (Figura IX.7). Para ello asignó a cada especie uno de cuatro mecanismos: corrientes de agua, viento, aves (transporte externo), aves (transporte interno). Encontró que en casi todos los casos, las aves, sobre todo a través de endozoocoria, eran responsables de más de la mitad de las llegadas exitosas. Le sigue la dispersión por corrientes marinas, ejemplos de la cual están representados en todas las islas. Finalmente, el mecanismo menos exitoso de dispersión es por viento; su eficiencia disminuye conforme aumenta la distancia con respecto a tierra firme. En algunos casos, como en las islas de Pascua, no se encontró ninguna especie que hubiera sido dispersada por viento.
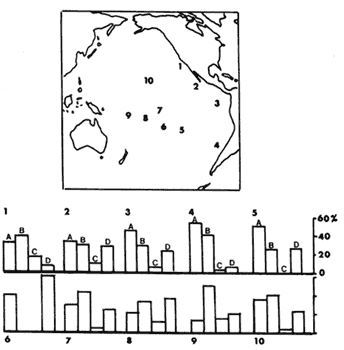
Figura IX.7. Principales mecanismos de dispersión (viento, aves —interno
y externo— y corrientes marinas) responsables de la colonización de islas
del Pacífico (modificado de Carlquist, 1967). 1. San Clemente, 2. Islas de Revillagigedo,
3. Galápagos, 4. Juan Fernández, 5. Isla de Pascua, 6. Oeno, 7. Marquesas,
8. Rarotonga , 9. Samoa, 10. Hawai. La columna A indica el porcentaje de semillas
dispersadas por aves (transporte externo), la B por aves (transporte interno),
la C por viento y la D por corrientes de agua.
Otro posible mecanismo de introducción de especies a las islas son las tortugas. Se considera que introducen varias especies en los atolones de Aldabra, en el Océano Índico. Una tortuga necesita cinco días para viajar de Madagascar a Aldabra, lo cual es mucho menos del tiempo que requiere para defecar la última comida (hasta 50 días).
En general, las aves es el grupo animal que más se encuentra en las islas, en contraste con el bajo número de mamíferos y reptiles (en Cuba, por ejemplo, sólo existen dos especies de víboras y en cambio las aves son muy numerosas; estos valores son semejantes en otras islas).
La colonización a largas distancias también se presenta en tierra firme, ya que hay distancias y barreras entre picos de montañas, lagos o entre manchones de hábitats similares. Trabajos realizados en las altas montañas en el Sudán africano demuestran que la flora tiene un alto componente de especies transportadas externamente por aves. Casi ninguna especie presenta señales de haber sido transportada de manera interna por aves u otros organismos. La gran altura del sistema (3 000 m/nm) puede constituir una barrera física para el paso de aves migratorias frugívoras. Otro grupo de especies es dispersado por el hombre y su ganado. Ello es lógico, debido a que esta región tiene una antigua tradición de pastoreo.
Datos de otro autor (Sugden, 1982) indican la existencia de 126 especies de plantas vasculares en Colombia, en la Serranía Macuira, con una altura de 865 m/nm. La flora de esta región tiene un origen reciente y no ha estado unida a otras zonas de alta montaña. Este investigador encontró que 41% de las especies presentaba dispersión endozoocora, 39% anemócora (viento), 5% ectozoocora y el resto era desconocida. Aunque no coincide totalmente con los resultados del Sudán, en ambos casos las aves desempeñaron un papel fundamental.
Actualmente, la tala y destrucción tan extensa que ha hecho el hombre de las selvas y bosques ha creado paisajes en donde la vegetación natural forma "islas", grandes espacios rodeados de campos agrícolas, ganaderos o bajo algún sistema de uso. Por tanto, para conservar estas "islas" con todas sus especies se necesitan mecanismos de dispersión que lleven las semillas a mayores distancias que hasta hace algunos años. Asimismo, se requieren organismos animales capaces de viajar entre islas, de cruzar otros medios ambientes distintos de la vegetación original. En la zona de Los Tuxtlas, en el sur de Veracruz, Sergio Guevara y sus colaboradores han demostrado que existe un intenso tránsito de aves ornitócoras entre los distintos manchones de selva (corredores a lo largo de ríos, cercas vivas, remanentes de selvas, etc.). Estas aves son muy importantes en el transporte de semillas y por lo tanto en la regeneración de numerosas especies arbóreas de la selva.
A este mismo problema se han venido enfrentando las reservas naturales. Y aunque éstas muchas veces tienen un tamaño suficiente para que las poblaciones vegetales puedan dispersarse y establecerse con éxito dentro del área delimitada artificialmente como reserva, no siempre es éste el caso. Frecuentemente, los parques y monumentos sólo cubren pequeñas extensiones.