III. ESTRUCTURA Y FORMACIÓN DE LAS SEMILLAS
LAS semillas constituyen una de las innovaciones más importantes
de las plantas vasculares que surgieron durante el curso de la evolución. Son
uno de los factores responsables del dominio actual de las plantas superiores
en la flora de nuestro planeta hoy en día. Cuando se examina una semilla inmadura,
o sea un óvulo, se ve que tiene una serie de tegumentos o capas que la protegen,
así como una gran cantidad de alimento almacenado. De este modo, el ovario no
sólo protege del medio ambiente al gametofito femenino, sino que también suministra
alimento a la nueva plántula que surge cuando la semilla germina. Este cuidado
parental de la semilla permite la mejor adaptación de las plantas vasculares
al medio ambiente y hace que predominen sobre otros grupos vegetales. La semilla,
por lo tanto, es fundamental en la sobrevivencia de las especies, como se vio
en el capítulo anterior. La evolución de la semilla constituye un mecanismo
invaluable de adaptación a la vida en la Tierra. La cubierta de la semilla o
testa la protege de muchas de las inclemencias del medio ambiente. El embrión
puede permanecer latente durante mucho tiempo, hasta que haya las condiciones
adecuadas para germinar y la existencia de sustancias de reserva le brinda alimento
hasta que la planta pueda sobrevivir por sí misma (capítulos IV, V y VI).
Las semillas más antiguas que se conocen proceden del periodo Devónico (hace
350 000 000 de años). Éstos fósiles fueron encontrados en rocas de Pensilvania,
EUA, pero no se tienen plantas fósiles que nos permitan conocer cómo eran las
plantas que las produjeron. Entre las semillas más antiguas que se conocen está
la Genemosperma, la cual todavía no ha desarrollado una cubierta protectora
completa, sino que está rodeada como por ocho lóbulos. En las semillas más avanzadas
estos lóbulos se han fusionado formando un tegumento protector (Figura III.1).
Durante los siguientes 50 000 000 de años se desarrollaron diversas formas de
semillas, algunas de las cuales estaban adheridas a plantas parecidas a los
helechos actuales. Pertenecían al grupo de las gimnospermas y hoy en día están
extintas. Se conocen con el nombre de Pteridospermae que significa "helechos
con semillas". En la figura aparece un dibujo de este tipo de plantas. Una de
ellas es muy parecida a un helecho y alcanzó los cinco metros de altura.
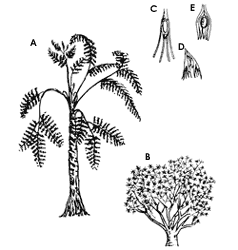
Figura III.1. Dibujo de dos plantas fósiles, las llamadas "helechos con
semillas", por su aspecto. En lugar de esporas, ya tienen semillas. A) La especie
Medullosa vivió en el Carbonífero Superior y el Pérmico; la Caytonia
(B), en el Triásico. C) Semilla de Genemosperma, del registro fósil,
constituye una de las primeras evidencias de la existencia de semillas. Tiene
un anillo con 8 lóbulos separados, la primera etapa en la evolución de los tegumentos.
D) En la semilla de la Archeosperma los lóbulos se han fusionado parcialmente;
es una etapa más evolucionada del proceso de formación de las semillas. E) Semilla
actual en la cual hay una fusión completa de los lóbulos que forman una cubierta
protectora a su alrededor (tomado de Thomas, 1981 y Gómez-Pompa y col. 1986).
En las plantas ancestrales productoras de semillas, éstas se separaban de la planta progenitora y se dispersaban antes de que el embrión madurara. Como el embrión constituye la parte fundamental de la semilla que dará origen a la raíz, tallo y hojas, la semilla tenía que madurar el embrión antes de germinar. En cambio, en la mayoría de las plantas productoras de semillas de hoy en día, el embrión madura antes de separarse de la planta progenitora. Por lo tanto, está protegido hasta que termina su desarrollo. Este es un mecanismo más por el que se incrementa el tiempo de cuidado materno de la nueva progenie.
A principios de la era Mesozoica, hace 230 000 000 de años, tuvo lugar una radiación de los reptiles que puede correlacionarse con el incremento de las masas continentales y por lo tanto, con una mayor gama de posibilidades y condiciones para los organismos terrestres. Había animales pequeños, grandes como los dinosaurios, acuáticos, terrestres y otros más que volaban. Entre las plantas predominaban las cicadales, gingkos y coníferas.
Hacia fines del Mesozoico y principios del Cenozoico hubo muchos cambios. Aparecieron las primeras angiospermas que probablemente vivían en regiones montañosas, mientras que las coníferas y cicadales dominaban las planicies. La aparición de nuevas clases de animales quizá fue el principal factor que dio la primera ventaja a las angióspermas o plantas con flores. Estas son superiores a las gimnospermas pues poseen mecanismos más eficientes de entrecruzamiento, dispersión y establecimiento de las semillas. Los insectos y las aves son agentes mucho más efectivos para transportar polen y semillas que el viento y el agua. Como ejemplo podríamos señalar que durante esta época, la familia de las abejas estaba en plena evolución. Los escarabajos, abejas, mariposas, aves y mamíferos primitivos debieron haber tenido una considerable influencia selectiva en la evolución de las familias y géneros de estas plantas.
Actualmente las plantas con semillas están divididas en dos grandes grupos: las gimnospermas y las angiospermas.
El término gimnosperma significa semilla desnuda y las plantas de este grupo reciben este nombre porque los óvulos y semillas de todos sus miembros se forman expuestos sobre la superficie del esporófilo. A las gimnospermas se les puede considerar como diversas líneas de plantas con semillas, con características comunes entre ellas. A continuación se mencionan las características más sobresalientes:
Es el grupo de gimnospermas más importante de la actualidad, siendo además el más conocido. Se distribuyen principalmente en el hemisferio norte y zonas aisladas del sur. Incluyen sobre todo árboles, entre ellos los más grandes y longevos del mundo, como las sequoyas. Tienen hojas perennes en forma de aguja (pinos, abetos, etc.) o escamosas (cipreses). Las flores son unisexuales, es decir o masculinas o femeninas. Por lo general cada árbol tiene flores de los dos sexos (monoicos), aunque algunos sólo tienen flores de un sexo en el mismo árbol. Los frutos se denominan conos y presentan una serie de escamas, llamadas brácteas, generalmente leñosas que protegen a las semillas.
Es un conjunto de plantas parecidas a las palmas que habitan principalmente
las zonas tropicales y subtropicales del mundo. Actualmente existen sólo
nueve géneros y alrededor de 100 especies. Sin embargo, llegaron a ser tan
comunes y numerosas durante el Mesozoico, que con frecuencia se habla de
la "era de las cycadas y de los dinosaurios". Estas plantas presentan las
hojas en la parte superior; al igual que las palmas, y producen una especie
de conos, donde se forman las semillas que están localizados en la parte
superior del tronco, en el centro del macizo de hojas. Existen individuos
masculinos que producen polen y otros femeninos que producen óvulos, donde
se forman las semillas (Figura III.2). Algunas especies llegan a alcanzar
hasta 18 metros de altura.

Figura III.2. Esquema de dos cycadas que habitan en México: Dioon
edule y Ceratozamia mexicana (tomado de Vovides, 1983).
En México existen representantes de los géneros Zamia, Dioon, y Ceratozamia. Habitan principalmente en selvas caducifolias y perennifolias, bosques mesófilos y encinares. Son plantas que actualmente están en peligro de extinción debido al comercio que se efectúa con ellas como elementos decorativos.
Este grupo casi ha desaparecido de la superficie de la Tierra. Actualmente sólo queda un representante: el Ginkgo biloba. Fue abundante, junto con otras pocas especies de la clase Ginkgoinae, durante el Mesozoico. Este árbol se reconoce fácilmente por sus hojas en forma de abanico. Se cree que ya no existen individuos en estado silvestre y se sabe que fue conservado en templos y jardines de China y Japón. En los últimos 150 años ha sido muy usado como elemento decorativo en los parques y jardines de Europa y América. Durante el otoño las hojas se tornan amarillas, dándole un aspecto muy hermoso a este árbol.
También existen individuos masculinos (portadores del polen) y femeninos (formadores del óvulo). En Ginkgo es posible que la fertilización de los óvulos se dé hasta que éstos se hayan separado de la planta progenitora. Los embriones se forman después de la fertilización, cuando las semillas ya están en el suelo.
Esta clase de gimnospermas está constituida por unas 70 especies, formadas únicamente por tres géneros. Tiene formas de vida muy caprichosas y poco comunes. Comprende plantas arbustivas que habitan ambientes desérticos o esteparios (Ephedra y Welwitschia) y lianas que viven en las selvas (Gnetum).
Las angiospermas o plantas con flores incluyen aproximadamente 300 000 especies, siendo el grupo de plantas más numeroso que existe actualmente. Son muy diversas en forma y estructura y dominan la superficie de nuestro planeta. Sus flores están compuestas por cáliz y corona que rodean los órganos propiamente reproductores. Los óvulos o células femeninas se encuentran cubiertos o protegidos por los llamados carpelos u hojas modificadas que se cierran sobre sí mismas, protegiendo así las células encargadas de la reproducción.
Se subdividen en dos grandes grupos: las monocotiledóneas que incluyen alrededor de 70 000 especies y las dicotiledóneas, con aproximadamente unas 230 000. Dentro de las primeras se encuentran familias de plantas tan importantes como las gramíneas (pastos), orquídeas, bromelias, palmas, etc. Entre las segundas están muchas de las plantas que adornan jardines, habitan bosques, desiertos, tundras, y que abarcan desde enormes árboles hasta pequeñísimas hierbas. Entre las familias más conocidas están: leguminosas, cactáceas, compuestas, solanáceas, crucíferas, magnoliáceas, urticáceas, moráceas, etcétera.
En las plantas con semillas las esporas femeninas, en vez de ser liberadas del esporangio, quedan retenidas y protegidas en interior del mismo. En este sitio germina la espora y produce un pequeño gametofito femenino, protegido por el tegumento que lo envuelve completamente, excepto por una pequeña abertura en la parte superior, el micrópilo. Posteriormente, el tegumento se desarrolla para formar la testa de las semillas.
Los gametos o células sexuales femeninas y masculinas son haploides (1n)
Los gametos de sexo opuesto se fusionan para formar un cigoto (2n). En
la figura III.3(a) se presenta el ciclo de vida de una gimnosperma, donde se
ve la alternancia de generaciones y la producción y desarrollo de las semillas
después de la fertilización. En la figura III.3(b) se presenta la misma situación
para una angiosperma.
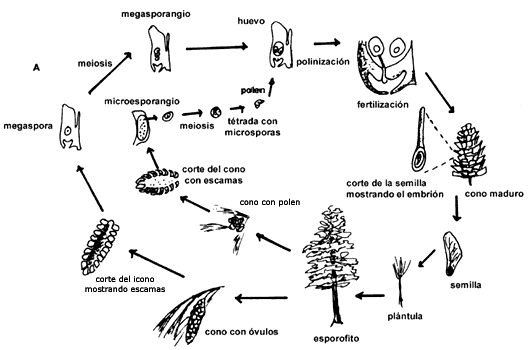
Figura III.3. (a) Ciclo de vida de una gimnosperma (tomado de Raven y Johnson,
1989).
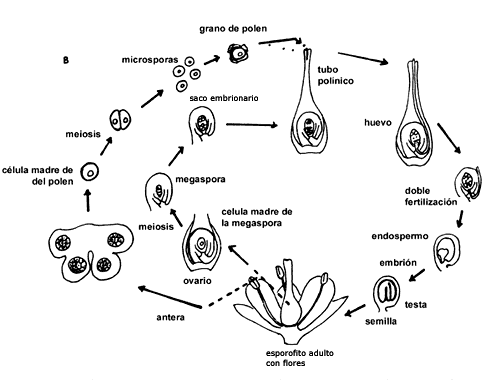
Figura III.3. (b) Ciclo de vida de una angiosperma (tomado de Raven y Johnson,
1989).
Las angiospermas y gimnospermas comparten, ambas, las primeras etapas del desarrollo del embrión, que se inicia con la división del huevo fertilizado o cigoto. Una de las dos células formadas dará origen a la parte superior del embrión y la otra a la parte inferior. Por medio de una progresión ordenada de divisiones el embrión se va diferenciando, iniciándose así la formación de los meristemos primarios que son los precursores de los futuros tejidos de la planta. Al mismo tiempo se van formando los cotiledones.
Las semillas, como se vio en el apartado anterior, son óvulos maduros. Se forman en el ovario, el cual se desarrolla para formar el fruto; sin embargo, hay ocasiones en que participan otras estructuras además del ovario en la formación del fruto.
La semilla, consta de una cubierta o testa, material alimenticio almacenado y un embrión.
Todas las semillas están rodeadas por una cubierta llamada testa (Figura III.4,
a y b), la cual puede tener muy distintas texturas y apariencias. Generalmente
es dura y está formada por una capa interna y una externa de cutícula y, una
o más capas de tejido grueso que sirve de protección. Estas características
le confieren a la testa cierto grado de impermeabilidad al agua y a los gases.
Ello le permite ejercer una influencia reguladora sobre el metabolismo y crecimiento
de la semilla. Frecuentemente en la testa se puede observar el micrópilo. En
muchas ocasiones está asociado con una cicatriz llamada hilio, que marca el
punto donde la semilla se separó del tallo (funículo) por medio del cual estaba
adherido al fruto. En algunas semillas estas estructuras de la testa están ausentes
pero lo que en realidad sucede es que se está observando el pericarpio de un
fruto y no la testa, como por ejemplo en el caso de Helianthus annuus
(el girasol, que pertenece a la familia de las compuestas) y de la lechuga.
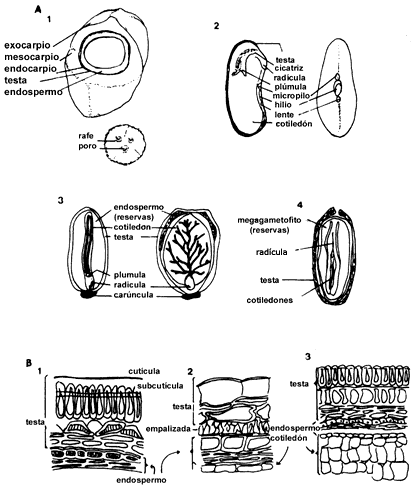
Figura III.4. A) Morfología de semillas de coco: Cocos nucifera (1),
de frijol, Phaseolus vulgaris (2), ricino, Ricinus communis (3)
y pino, Pinus pinea (4). Muestran las distintas estructuras externas
de la testa y algunos aspectos de la morfología interna. B) Anatomía de la testa
de tres especies (1) Melilotus alba, (2) Sinapsis alba, mostaza
y (3) Glycine max, soya. Se puede observar las diferentes capas que la
componen. Esto permite cierta complejidad y variación en las cualidades de la
testa entre las distintas especies (tomado de Hamly, 1932 y Vaughan, 1970).
El endospermo tiene como función almacenar las reservas alimenticias de las semillas, aunque no siempre está presente. Entre las semillas que tienen un endospermo bien desarrollado están las gramíneas como el trigo, el maíz, la cebada y algunas dicotiledóneas como Ricinus communis. En estos casos los cotiledones son relativamente pequeños. El endospermo de las gramíneas y de otras especies se caracteriza por presentar una capa externa o aleurona. Tienen paredes gruesas y en su interior se desarrollan los llamados granos de aleurona. Estas células permanecen vivas, a diferencia de las células del endospermo de otros cereales, las cuales se convierten en células muertas empacadas con almidón y algo de proteínas.
El embrión es el origen de la raíz, hojas y tallo de la nueva planta, por lo que resulta de interés entender con más detalle su funcionamiento. El embrión maduro de las plantas que tienen flores consiste en un eje parecido a un tallo (eje embrionario) en cuyo extremo están uno o dos cotiledones (Figura III.4(a)). Estos cotiledones frecuentemente se conocen como las hojas de las semillas o las hojas cotiledonarias, debido a que son las primeras hojas en aparecer, aunque tienen forma y función diferentes de las hojas que aparecerán subsecuentemente durante la vida de la planta. En ambos extremos del eje embrionario hay meristemos formados por células con gran capacidad de reproducción, responsables del crecimiento. En el embrión, el meristemo apical del tallo se localiza en la parte superior del eje embrionario, justo arriba de los cotiledones, y por eso se le conoce como epicótilo —arriba de los cotiledones—. En algunos embriones el epicótilo consta solamente del meristemo apical, mientras que en otros, presenta una o más hojas jóvenes. En este último caso, el epicótilo, junto con las hojas jóvenes, se denomina plúmula. La parte del eje embrionario entre el epicótilo y el ápice de la raíz se llama hipocótilo, por encontrarse inmediatamente abajo de los cotiledones. Finalmente, en el extremo se encuentra el ápice de la raíz o radícula.
Estas partes son mucho más fáciles de identificar en las dicotiledóneas que en las monocotiledóneas. En las últimas, el cotiledón único se llama escutelo. La envoltura basal del cotiledón se ha elongado para formar el coleoptilo y en algunas especies el hipocótilo se ha modificado parcialmente. La coleorriza puede considerarse como la base del hipocótilo que envuelve la radícula.
Sintetizando, diríamos que el embrión está formado básicamente por un eje hipocótilo-raíz con uno o dos cotiledones (dependiendo si son mono o dicotiledóneas) y un meristemo apical en los ápices de raíz y tallo.
Como ya se había mencionado anteriormente, las plantas con flores se dividen en monocotiledóneas, o sea aquellas que tienen un solo cotiledón, como sucede en las gramíneas; y en dicotiledóneas, o sea aquellas que tienen dos cotiledones, como sucede en la mayoría de las angiospermas: leguminosas, compuestas, lauráceas, etc. Los cotiledones de la mayoría de las plantas dicotiledóneas son carnosos y contienen las sustancias de reserva de las semillas. En algunas dicotiledóneas, y en la mayoría de las monocotiledóneas, las sustancias de reserva están almacenadas en el endospermo y los cotiledones, que son delgados y muy delicados, funcionan como estructuras de absorción. Su papel fundamental estriba en absorber el alimento ya digerido en el endospermo y transportarlo a las partes del embrión que están creciendo.
Durante el proceso de germinación, generalmente la primera estructura en emerger de la semilla es la raíz del embrión, llamada radícula. Esta raíz rápidamente penetra en el suelo y permite que la planta se ancle y comience a absorber agua y nutrientes.
Con el paso del tiempo los cotiledones disminuyen de tamaño, se van secando y finalmente se desprenden. Todas las sustancias almacenadas en ellos ya han sido utilizadas por la nueva plántula y por lo tanto sólo quedan restos de lo que eran. Para este momento ya han transcurrido varios días y a veces hasta semanas, y la plántula, que antes dependía de los cotiledones para obtener su alimento, ya es una planta capaz de obtener del suelo y del Sol lo que necesita para sobrevivir. Absorbe elementos del suelo y lleva a cabo la fotosíntesis activamente. En este momento ya se le considera una planta independiente y establecida. El periodo de tiempo que transcurre entre el momento en que la semilla germina y en el que la plántula se establece como un organismo independiente constituye una de las fases decisivas y más delicadas en el ciclo de vida de la planta. Es el momento en que el individuo es más susceptible a una gran cantidad de daños, como enfermedades por hongos, depredación por insectos, sequía, desenterramiento, etc. La mortandad en la etapa de plántula es enorme y sólo unos cuantos individuos llegan a establecerse.
Como vemos, las plantas tienen varias estrategias para afrontar esta etapa de la vida. Encontramos una gran diversidad de tamaños, formas, velocidades de respuesta, etc., que iremos considerando a lo largo de este libro.
COMPOSICIÓN QUÍMICA DE LAS SEMILLAS
La composición química de las semillas es variable. Se puede hablar por un lado de un conjunto de compuestos que está presente en todos los tejidos y por el otro de un conjunto de compuestos de almacenamiento de reservas que está presente en las semillas en grandes cantidades. Además, muchas semillas presentan distintos tipos de compuestos secundarios, cuya presencia y tipo son muy variables. En general se puede decir que las proteínas que se encuentran en las semillas difieren en composición química y propiedades con respecto a las proteínas presentes en otras partes de la planta; asimismo, hay gran cantidad de lípidos, lo cual no es frecuente en otros tejidos vegetales.
Las semillas se pueden dividir en función del tipo de material que almacena reservas. En algunas son principalmente los carbohidratos y en otras son los lípidos o grasas, predominando estas últimas. Entre las especies en que los carbohidratos alcanzan valores altos están el maíz (Zea mays con 50-70% de carbohidratos en la semilla secada al aire contra 5% de lípidos), el chícharo (Pisum sativum, con 30-40 contra 2), el castaño (Castanea vesca, con 42 contra 3) y el encino (Quercus pendunculata, con 47 contra 3). Por otro lado, entre las especies con un mayor porcentaje de lípidos en materia secada al aire está el cacahuate (Arachis hypogaea, con cantidades entre 8-21% de carbohidratos y 40-50% de lípidos), el girasol (Helianthus annuus, que no presenta carbohidratos y en cambio tiene entre 4 y 50% de lípidos) y el ricino (Ricinus communis, con 64% de lípidos y 0 carbohidratos).
Son pocos los casos en que las proteínas funcionan como reservas. Esto sucede en el frijol de soya, por ejemplo, el cual tiene 40% de proteínas, 18% de lípidos y 7% de carbohidratos.
Como ya se ha mencionado, los materiales de reserva se pueden almacenar en el embrión o en tejidos extraembrionarios, como el endospermo. En muchas especies, como algunas leguminosas, las proteínas y carbohidratos se encuentran en los cotiledones, los cuales pueden permanecer bajo tierra durante el crecimiento de la plántula (Pisum sativum, chícharo) o conforme crece el hipocótilo van siendo levantados sobre la superficie del suelo (Phaseolus vulgaris, frijol). Los cotiledones también funcionan como estructuras de almacenamiento para lípidos y proteínas de especies como la calabaza (Cucurbita pepo) y la mostaza (Sinapis alba). Sólo ocasionalmente el propio eje embrionario funciona como región de almacenamiento, como sucede con la nuez del Brasil (Bertholletia excelsa). En la tabla III.1 aparece la composición de las reservas de algunas semillas y el principal órgano de almacenamiento. Puede verse que la mayor parte de los carbohidratos (almidón) y proteínas de los cereales y otras gramíneas se almacenan en el endospermo. Gracias a estos ejemplos vemos que las reservas no son exclusivas de un tejido. Algunas de ellas se localizan en varios tejidos; también se observa que un solo tejido puede contener todas o casi todas las reservas de las semillas y por lo tanto que diferentes reservas coexisten en un tejido.
|
|
||||||
|
|
||||||
|
Composición promedio
|
Principal
|
|||||
|
Especies
|
(% de peso seco)
|
órgano de
|
||||
|
P.
|
L.
|
Ch.
|
almacenamiento
|
|||
|
|
||||||
| Zea mays - maíz |
11
|
5
|
75
|
almidón
|
endospermo
|
|
| Zea mays - maíz dulce |
12
|
9
|
70
|
almidón
|
endospermo
|
|
| Avena sativa - avena |
13
|
8
|
66
|
almidón
|
endospermo
|
|
| Triticum aestivum - trigo |
12
|
2
|
75
|
almidón
|
endospermo
|
|
| Secale cereale - centeno |
12
|
2
|
76
|
almidón
|
endospermo
|
|
| Hordeum vulgare - cebada |
12
|
3
|
76
|
almidón
|
endospermo
|
|
| Vicia faba - haba |
23
|
1
|
56
|
almidón
|
cotiledón
|
|
| Linum usitatissimum - lenteja |
24
|
36
|
24
|
almidón
|
cotiledón
|
|
| Pisum arvense - chícharo |
24
|
6
|
56
|
almidón
|
cotiledón
|
|
| Arachis hypogaea - cacahuate |
31
|
48
|
12
|
almidón
|
cotiledón
|
|
| Glycine max - soya |
37
|
17
|
26
|
almidón
|
cotiledón
|
|
| Gossypium spp. - algodón |
39
|
13
|
15
|
cotiledón
|
||
| Citrullus vulgaris - sandía |
38
|
48
|
5
|
cotiledón
|
||
| Bertholletia excelsa - nuez |
18
|
68
|
6
|
radícula
|
||
| del Brasil |
hipocótilo
|
|||||
| Elaeis guineensis - coquito |
9
|
49
|
28
|
endospermo
|
||
| Phoenix dactilifera - dátil |
6
|
9
|
58
|
endospermo
|
||
| Ricinus communis - ricino |
18
|
64
|
|
trazas
|
endospermo
|
|
| Pinus pinea - pino |
35
|
48
|
6
|
megagametofito
|
||
|
|
||||||
La composición química de las semillas está determinada genéticamente pero las cantidades relativas pueden variar en función de factores ambientales como la presencia de nutrientes minerales o el clima.
El almidón y la hemicelulosa son los dos tipos de carbohidratos que almacenan reservas. Es común encontrar el primero de ellos en la mayor parte de los cereales y las leguminosas. Además, las semillas contienen otros tipos de carbohidratos que no cumplen una función de almacenamiento, como por ejemplo los mucílagos que recubren la testa de algunas semillas y que posiblemente ayudan a la dispersión y fijación de la semilla al sustrato. Los mucílagos se encuentran sobre la superficie de la testa o en celulas especiales en la misma testa, como sucede en Brassica alba. Las hemicelulosas, tanto el pentosano como el hexosano, están presentes en el endospermo de las palmas y en los cotiledones de algunas leguminosas.
El almidón se localiza en cuerpos subcelulares denominados granos de almidón. Tienen una apariencia característica para cada especie y son esféricos, angulares o elípticos. La forma del grano depende de la cantidad de amilosa, uno de los polímeros que conforman este polisacárido; así, mientras más redondeados sean, mayor cantidad de amilosa poseen.
Los lípidos están presentes en forma de glicéridos de ácidos grasos. En realidad son un grupo de sustancias químicas heterogéneas. La mayoría son del tipo no saturado y entre los principales están el ácido oleico, el linoleico y el linolénico. El contenido de lípidos de algunas semillas aparece en la tabla III.1. Los triglicéridos se localizan en forma de organelos subcelulares que han recibido varios nombres (oleosomas, cuerpos de aceites, etc.). Cuando las semillas tienen grandes depósitos de lípidos, estos cuerpos ocupan todo el espacio, excepto el de los otros organelos.
Las semillas contienen muchas proteínas que metabólicamente están inactivas y que funcionan como reservas, las cuales varían según la especie. También presentan proteínas que metabólicamente son activas, como las enzimas proteicas. En el trigo, por ejemplo, se han encontrado por lo menos cuatro tipos de proteínas: prolaminas, glutelinas, globulinas y albúminas, predominando las dos primeras. El total de proteínas activas sólo alcanza el 15% del total, siendo las principales las albúminas. En la avena (Avena sativa), el 80% del porcentaje de proteína total corresponde a la globulina, 15% a la prolamina y 5% a la glutelina; en el pepino (Cucurbita pepo) también predomina la globulina. En cambio en el arroz (Oryza sativa), 80% corresponde a la glutelina, 10% a la globulina y 5% a la albúmina y a la prolamina. En el maíz (Zea mays) los valores son 31, 0, 14 y 48% respectivamente.
Cabe mencionar que la subdivisión de las proteínas de almacenamiento es algo arbitraria, ya que se basa principalmente en su capacidad para solubilizarse en ácidos y bases débiles. Estas proteínas generalmente se presentan en organelos bien definidos llamados cuerpos de proteínas o granos de aleurona. Estos cuerpos se han encontrado en numerosas semillas como oleaginosas y los cereales. En estos últimos se localizan en la mayoría de las células del endospermo, pero en especial se le llama capa de aleurona a estos cuerpos de proteínas que predominan en la capa que rodea al endospermo. En general tienen un alto contenido de nitrógeno y de prolina, y una baja cantidad de lisina, triptofano y metionina.
Además de los compuestos mencionados, las semillas contienen muchas otras sustancias. Contienen algunos minerales, y la composición de éstos es similar a la del resto de la planta. Además del nitrógeno que está presente en las proteínas, las semillas contienen cierta cantidad de aminoácidos y de amidas (por ejemplo, glutamina y aspargina en las nueces). Otros compuestos con nitrógeno son los alcaloides, como la piperina en semillas de Piper nigrum, la ricinina en Ricinus communis, la cafeína en Coffea spp., la teobromina y cafeína en Theobroma cacao, etc. Támbién se han encontrado vitaminas en todas las semillas analizadas, aunque su función se desconoce.
Éstas son sólo algunos ejemplos de la gran cantidad y variedad de sustancias presentes en las semillas; sin embargo, es importante mencionar que la mayoría de los análisis se ha hecho en semillas de interés comercial o farmacéutico, y que aún se sabe muy poco sobre la composición de las semillas silvestres.