I. LAS SEMILLAS
LA SEMILLA es el principal órgano reproductivo de la gran mayoría de las plantas superiores terrestres y acuáticas. Ésta desempeña una función fundamental en la renovación, persistencia y dispersión de las poblaciones de plantas, la regeneración de los bosques y la sucesión ecológica. En la naturaleza la semilla es una fuente de alimento básico para muchos animales. También, mediante la producción agrícola, la semilla es esencial para el ser humano, cuyo alimento principal está constituido por semillas, directa o indirectamente, que sirven también de alimento para varios animales domésticos.
La semilla es uno de los principales recursos para el manejo agrícola y silvícola de las poblaciones de plantas, para la reforestación, para la conservación del germoplasma vegetal y para la recuperación de especies valiosas sobreexplotadas. Las semillas pueden almacenarse vivas por largos periodos, asegurándose así la preservación de especies y variedades de plantas valiosas.
La ciencia de las semillas se ha desarrollado a lo largo de muchos años, acumulándose hasta la fecha un importante volumen de conocimientos acerca de muchos aspectos de su biología y manejo. Existen numerosas publicaciones científicas y técnicas en este campo y se conocen con detalle varias características de la biología de las semillas de las plantas cultivadas más importantes, y de algunos árboles de valor forestal; sin embargo, las semillas de las plantas tropicales y subtropicales no han corrido con igual suerte y su estudio se ha quedado muy rezagado.
Como parte del estudio de las plantas es necesario intensificar la investigación de las semillas, sus características fisiológicas, sus mecanismos de latencia y germinación, su longevidad (ecológica y potencial) y su posible uso para la propagación y conservación de las plantas.
La semilla es una unidad reproductiva compleja, característica de las plantas vasculares superiores, que se forma a partir del óvulo vegetal, generalmente después de la fertilización. Se encuentra en las plantas con flores (angiospermas) y en las gimnospermas. En las angiospermas los óvulos se desarrollan dentro de un ovario; en tanto que en las gimnospermas la estructura que los contiene es muy diferente, pues no constituye una verdadera flor; sin embargo, la estructura de las semillas de estas plantas es básicamente similar a la de las plantas con flores.
Para entender la ontogenia de la semilla, es decir, su formación y desarrollo, se requiere cierto conocimiento de la estructura de las flores y de los órganos reproductivos masculinos y femeninos, En términos generales, las flores consisten de varias capas de hojas modificadas que constituyen, de afuera hacia adentro, los sépalos, los pétalos, los estambres y el ovario. Hay grandes variaciones entre especies en la forma, tamaño y disposición de estas estructuras. Hay también plantas que producen flores con sexos separados, ya sea masculinas con estambres o femeninas con ovarios. Los estambres producen granos de polen que contienen los gametos masculinos, y el ovario contiene los óvulos que producirán a las semillas (figura 2). Los óvulos de las plantas superiores son estructuras pluricelulares, relativamente complejas. En su interior, el óvulo contiene al saco embrionario y éste a su vez contiene varias células haploides, no claramente delimitadas, cuyo número varía entre los diferentes grupos taxonómicos.
Al arribar el polen al pistilo, que es el órgano femenino, se produce la polinización,
con la consecuente formación del tubo polínico, que desciende por el estilo
hacia el óvulo. El tubo polínico lleva dos núcleos haploides: el gameto masculino
que se fusionará con el núcleo del saco embrionario que funciona como cigoto
o huevo, para originar un nuevo organismo diploide, y otro que se fusionará
con los dos núcleos polares del saco embrionario para formar un tejido triploide,
el endospermo. Este proceso se conoce como doble fecundación. Tanto la estructura
de estos órganos como la doble fecundación, que caracteriza a las plantas, son
complejas y escapa a los objetivos de este libro; sin embargo, están descritas
con mucho detalle en los textos de botánica. En la figura 3 se precisan las
principales estructuras participantes y resultantes de la fecundación.
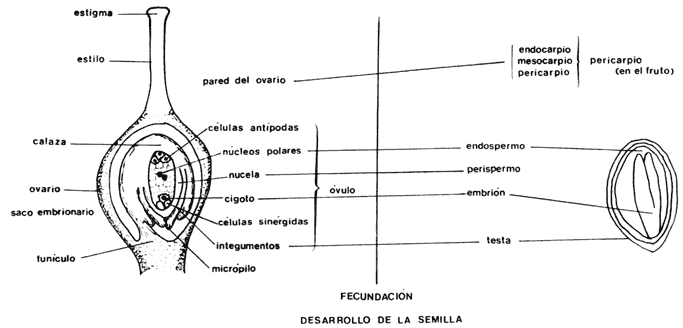
Figura 3. Estructura del ovario de una flor. Se muestran las diferentes
capas que lo componen y cómo se transforma después de la fecundación cada estructura
en los diversos componentes morfológicos de la semilla.
Al momento de la fertilización un óvulo típico de angiospermas está compuesto por una o dos capas protectoras y por el saco embrionario que contiene al núcleo que funciona como gameto. El óvulo está unido a la pared del ovario por el funículo. Después de la fertilización el huevo comienza a dividirse hasta formar el embrión de una nueva planta: las cubiertas exteriores o tegumentos originarán la cubierta de la semilla, como hemos visto, las otras células del tejido central en algunos casos originarán el endospermo, que contiene las reservas de la semilla que serán utilizadas en el desarrollo inicial de la nueva planta. En muchas especies el endospermo no se forma y es el embrión el que acumula las sustancias de reserva, generalmente en los cotiledones u hojas embrionarias, que pueden llenar todo el interior de una semilla. En la naturaleza se encuentran infinidad de variantes en la estructura de las semillas; por ejemplo, en las semillas pequeñas el embrión puede poseer un cúmulo de pocas células o tener ya la forma de una pequena planta en la que se distinguen claramente la radícula, los cotiledones u hojas embrionarias, la plúmula de la cual se desarrollará el tallo y el hipocótilo, conectando entre sí todas las estructuras (figura 4).
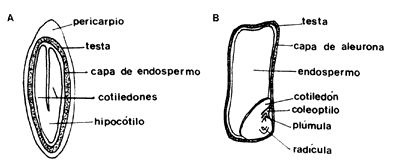
Figura 4. Anatomía de la semilla de una dicotiledónea y de una monocotiledónea.
Las reservas energéticas de la semilla son: grasas, carbohidratos y a veces proteínas que sostendrán a la futura planta durante sus primeras etapas de vida. Estas reservas, como hemos dicho, pueden encontrarse en diferentes tejidos o en el embrión mismo, y todo esto está relacionado con la germinación y el desarrollo de un nuevo individuo.
Existe cierto paralelismo en el desarrollo de las semillas en las plantas superiores y el desarrollo de los fetos en los mamíferos. En ambos casos existe una conexión de tipo placentario que transfiere recursos para el desarrollo del embrión, que aumenta sus probabilidades de sobrevivencia después de la separación de la madre, durante la germinación y las primeras etapas del establecimiento. La diferencia principal estriba en que esta separación, en el caso de los animales, se lleva a cabo por el propio movimiento de la descendencia, en tanto que en las plantas, el alejamiento de los descendientes de la planta madre ocurre en forma pasiva y con frecuencia auxiliada por agentes externos como el viento, el agua y los animales que se encargan de diseminar las semillas. En esta función el ovario tiene un papel importante, ya que con frecuencia se transforma en la estructura que va a permitir la dispersión de la semilla, bien sea formando un fruto carnoso, una estructura en forma de alas o pelos, o un fruto seco que al deshidratarse se abre bruscamente expulsando las semillas. En algunos casos, partes del fruto pueden llegar a formar parte de las cubiertas de la semilla, como en las gramíneas y algunas frutas como el durazno. En resumen, una semilla madura contiene tejidos originados directamente de la planta madre y tejidos formados por la nueva combinación genética producida por la fertilización.
La gran diversidad de formas de los frutos de las angiospermas está muy relacionada con los mecanismos de dispersión. El crecimiento y desarrollo de las plantas jóvenes bajo el árbol que las engendró es muchas veces difícil debido a la falta de luz, a la competencia de las raíces por nutrientes y a la presencia de una gran densidad de parásitos y depredadores específicos en esa área; la dispersión sobre una área más amplia asegura que algunas semillas encuentren condiciones adecuadas para germinar y crecer. Sin embargo, en las comunidades naturales la gran mayoría de ellas perece por los efectos de un ambiente inadecuado para establecerse, por la competencia con otras plantas, por depredación por animales o parásitos y por enfermedades.
En la naturaleza existen infinidad de variantes con respecto a la estructura de los frutos y los mecanismos de dispersión de las semillas, que han sido objeto de numerosos estudios. En el cuadro 1 se resumen los diferentes mecanismos de dispersión existentes, y su relación con los agentes que transportan las semillas.
UADRO 1. Mecanismos de dispersión y su relación con las características
de las diásporas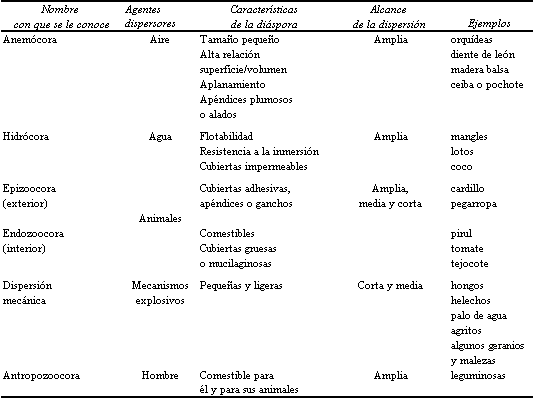
El tamaño de las semillas entre diferentes especies de plantas varía en una forma impresionante, a pesar de que se trata de un órgano vegetal cuyo origen ontogenético es constante y que tiene una función bien definida. Hay aproximadamente 11 órdenes de magnitud de diferencia en tamaño entre las semillas más pequeñas y las más grandes que existen en la naturaleza. Las semillas de una orquídea pueden pesar 0.1 mg, en tanto que la palma de coco doble del Pacífico produce semillas de 10 kg de peso. En una comunidad natural el rango de variación es menor pero es aún muy amplio; por ejemplo, en la selva tropical, éste es de aproximadamente seis órdenes de magnitud.
Los recursos de una planta para producir semillas son limitados, así que cierta cantidad de energía disponible para producirlas puede traducirse en un gran número de semillas pequeñas o en un número menor de semillas grandes. El número producido y su tamaño afectarán la capacidad de sobrevivencia y perpetuación de las especies. Las plantas que producen muchas semillas pequeñas se diseminan más ampliamente y tienen mayores oportunidades de encontrar un sitio favorable para germinar y crecer; sin embargo, su tamaño pequeño aporta poco al crecimiento de la nueva planta y ésta depende muy pronto de los recursos disponibles en su medio, por lo que su riesgo de morir es muy alto. También tienen menor resistencia a los efectos de la defoliación por herbívoros y pueden ser aplastadas fácilmente por la hojarasca que cae al suelo. Aunque esto se compensa de alguna manera por el gran número, sólo una pequeña fracción sobrevive a todos esos accidentes.
Las semillas grandes se producen en menor número y frecuentemente se diseminan a distancias más cortas, pero cuentan con mayor cantidad de recursos para iniciar su crecimiento y establecimiento en lugares con escasez de recursos, por ejemplo en la sombra de los bosques, ya que producen plántulas más grandes y resistentes, con mayor superficie de raíces y de hojas.
Las semillas de plantas que necesitan Sol (plantas heliófilas) suelen ser pequeñas, mientras que aquellas que se establecen a la sombra de otros árboles, como la mayoría de las semillas de los árboles de bosques maduros, suelen ser mayores. Sin embargo, existen muchas excepciones a esta afirmación. Se dice que las semillas grandes producen plántulas con mayor superficie radical y foliar antes de depender totalmente de los recursos externos, lo cual les permite sobrevivir en condiciones de baja disponibilidad de energía luminosa y nutrientes.
Un componente muy importante que determina el tamaño de las semillas es el origen filogenético de las plantas. La anatomía de los órganos reproductivos de las plantas varía entre diferentes linajes y estas características determinan un tamaño inicial de semilla en el grupo, sobre el cual la selección natural puede o no haber actuado.
Las plantas presentan frecuencias de floración y de subsecuente fructificación que van desde la producción continua de frutos a lo largo del año, como ocurre en algunos pioneros, a la producción sincrónica de frutos dentro de una generación de plantas a intervalos de más de un siglo, como ocurre en algunas especies de bambú. Entre estos extremos existen varios patrones de producción de semillas. Uno muy frecuente es la fructificación anual de duración relativamente fija; sin embargo, el tamaño de la cosecha de semillas de la población de una especie, o de ésta, entre plantas individuales, puede variar. El periodo de fructificación cambia entre diferentes localidades, incluso dentro de una misma región. Esta variabilidad se debe a variaciones en la disponibilidad de recursos para la reproducción o a los ciclos endógenos que diferencian distintos niveles de esfuerzo reproductivo entre años.
Algunas regiones caracterizadas por un clima muy homogéneo y poco estacional contienen poblaciones de plantas que presentan fructificación masiva cada pocos años seguidas por años de baja o nula producción de propágulos sexuales; mientras que en áreas climáticamente más estacionales son característicos los periodos bien definidos de fructificación. En el cuadro 2 se resumen los principales ciclos periódicos en la floración.
La regulación fisiológica de los diferentes patrones de reproducción y los sensores que los regulan son pobremente conocidos, no obstante, las presiones del ambiente han inducido la selección de los diferentes comportamientos fenológicos, los cuales han sido ampliamente analizados por muchos autores. La mayoría de ellos está de acuerdo en que los patrones fenológicos representan adaptaciones a presiones de tipo abiótico y biótico. Entre los factores abióticos, el fundamental parece ser la estacionalidad en la disponibilidad de recursos, por ejemplo la estación húmeda y la seca, además de condiciones ambientales apropiadas para la reproducción, como la disponibilidad de agua para la floración y fructificación y temperaturas favorables para la actividad de los polinizadores. Las presiones bióticas han sido objeto de mucha discusión y han generado diversas opiniones; sin embargo, la principal hipótesis acerca del origen de las diferentes periodicidades de floración y fructificación está basada en la idea de que éstas reducen la competencia de las plantas por polinizadores y dispersores.
UADRO 2. Periodicidad de la floración en zonas tropicales
de acuerdo con los criterios de Alvim en 1974
Las especies de plantas compiten entre ellas para atraer polinizadores de flores y dispersores de semillas, por lo cual han desarrollado comportamientos que minimizan la sobreposición fenológica con otras plantas que dependen del mismo vector animal en cada comunidad vegetal.
La fotosíntesis genera compuestos orgánicos que son invertidos en gasto respiratorio, crecimiento, reposición de partes, tejidos y órganos, y finalmente en la reproducción, por lo que la cantidad de semillas producidas por cada planta individual, en proporción a su biomasa total y al rendimiento fotosintético anual, es muy variable entre especies y poblaciones. Las plantas anuales colocan gran parte de su productividad fotosintética anual en biomasa reproductiva porque tienen una sola oportunidad en la vida de dejar descendientes, pero la mayoría de las plantas leñosas producen propágulos muchas veces durante su vida, por lo que esta producción se logra normalmente utilizando una menor proporción de la productividad fotosintética anual. Otro factor importante relacionado con el volumen de la cosecha es la frecuencia de la fructificación. Se ha visto que las plantas que fructifican frecuentemente producen menos propágulos que aquellas que lo hacen esporádicamente, y que las de fructificación estacional ocupan una posición intermedia. En el sureste de Asia existe el fenómeno conocido como fructificación "mast", el cual consiste en la producción masiva de frutos cada cierto número de años, seguida de una producción escasa en años intermedios.
Con gran frecuencia se observan años buenos y años malos en la producción de frutos en muchas plantas tropicales. Durante los años buenos las semillas son abundantes, sanas y con alta viabilidad, en tanto que en los años malos éstas son escasas, mal desarrolladas y de baja viabilidad. Esta variación está ligada a la calidad de la estación favorable para la productividad fotosintética y a factores bióticos, como la abundancia de polinizadores o parásitos de flores y frutos.
La germinación de las semillas comprende tres etapas sucesivas que se superponen
parcialmente: 1) la absorción de agua por imbibición, causando su hinchamiento
y la ruptura final de la testa (figura 5); 2) el inicio de la actividad
enzimática y del metabolismo respiratorio, translocación y asimilación de las
reservas alimentarias en las regiones en crecimiento del embrión, y 3)
el crecimiento y la división celular que provoca la emergencia de la radícula
y posteriormente de la plúmula. En la mayoría de las semillas el agua penetra
inicialmente por el micrópilo y la primera manifestación de la germinación exitosa
es la emergencia de la radícula (figura 6).
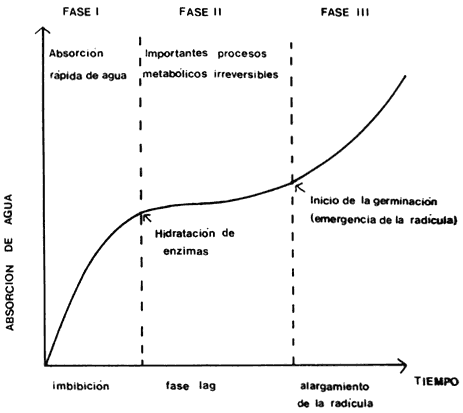
Figura 5. Etapas de la germinación que conducen a la emergencia de la radícula. Se inician con la absorción de agua y la activación metabólica del embrión.
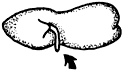
Figura 6. Semilla germinada que muestra la ubicación y apariencia de la radícula.
Existen varias etapas de desarrollo de la plántula cuyas características varían, dependiendo del tipo de germinación que presenta cada especie. Hay básicamente dos tipos de germinación (que a veces presentan algunas variantes), la germinación epigea y la hipogea. En la germinación epigea el hipocótilo se alarga y aleja a los cotiledones del suelo; en tanto que en la germinación hipogea el hipocótilo no se desarrolla y los cotiledones permanecen bajo el suelo o ligeramente sobre éste. En este caso las hojas cotiledonarias tienen sólo una función almacenadora de nutrientes, en tanto que en la germinación epigea estas hojas también tienen con frecuencia color verde y realizan funciones fotosintéticas durante el crecimiento temprano de la plántula. La testa de la semilla puede permanecer cubriendo los cotiledones en el caso de la germinación hipogea, en tanto que en la epigea se desprende, lo cual permite la expansión de las hojas cotiledonarias (figura 7).
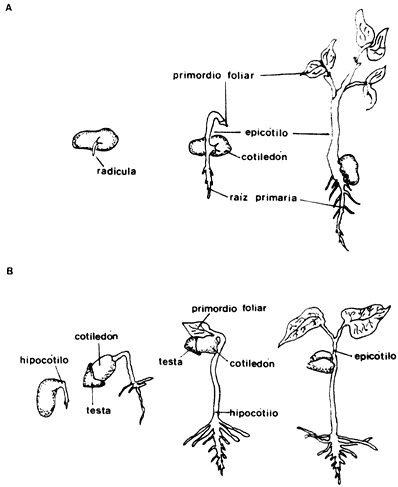
Figura 7. A) Germinación hipogea en haba. B) Germinación epigea en frijol.
En el trópico las semillas presentan tipos de germinación intermedios entre los dos descritos. Por ejemplo, en algunos casos los cotiledones son expuestos y fotosintéticos pero permanecen al nivel del suelo, y en otros casos se elevan sobre el suelo, sin embargo, permanecen plegados dentro de la cubierta de las semillas. Cada una de estas variantes ha recibido nombres que varían según los autores; la nomenclatura asignada a estas variantes hace hincapié en su carácter de presentación intermedia de los mecanismos de germinación.
Germinación de las semillas en el suelo
La gran diversidad de las plantas se refleja en la multitud de posibilidades del desarrollo y la temporalidad de la germinación de las semillas de diferentes especies. Existen plantas que diseminan sus semillas cuando ya han germinado en el fruto; en el otro extremo, algunas semillas que son dispersadas están provistas de una dura testa impermeable que sólo permite la germinación después de muchos meses de desgaste.
Cuando llegan las semillas al suelo, el recurso clave para iniciar los cambios fisiológicos que conducen a la germinación es el agua, que resulta indispensable para activar el metabolismo y el crecimiento de las células vivas de los tejidos de las semillas. La cantidad de agua que absorbe una semilla y la velocidad a la que lo hace no sólo dependen de las características de la semilla, como la permeabilidad de sus cubiertas, la composición química de sus reservas, su tamaño y su contenido de humedad, sino que también están determinadas por condiciones ambientales como la humedad del suelo, la humedad del aire y la temperatura.
Muchos árboles del bosque tropical producen semillas con alto contenido de humedad, cuya germinación se ve favorecida en ambientes húmedos y sombreados dentro del bosque. Es frecuente que estas semillas no puedan germinar bien en suelos desnudos que reciben insolación directa, pues la pérdida de agua de la superficie de la semilla a la atmósfera supera la cantidad absorbida en la interfase semilla-suelo, por lo que raramente alcanzan altas tasas de germinación en suelos en los que la evaporación es muy alta. Estas semillas, al igual que muchas otras de los bosques tropicales, tienden a germinar casi de inmediato que llegan al suelo, cuando las condiciones de humedad son adecuadas. Con frecuencia, en pocos días la radícula emerge de las cubiertas de la semilla y en pocas semanas ocurre la total germinación de las semillas viables. El periodo que transcurre entre la liberación de las semillas de los frutos y su llegada al suelo no se caracteriza por una latencia profunda, ya que el único factor que determina la germinación es la disponibilidad de agua.
Por otra parte, algunas semillas pueden tolerar cierto grado de sequía cuando se encuentran en el suelo, perdiendo humedad y entrando en un estado quiescente (de reposo) hasta que se incrementa la humedad del suelo, al inicio de la estación de lluvias. Las semillas del cedro y la caoba de América tropical son dos ejemplos de semillas que son dispersadas por el viento en la estación seca, que son liberadas con bajo contenido de humedad y que permanecen quiescentes en el suelo hasta que las primeras lluvias permiten su germinación. La aparición de multitud de plántulas de árboles en el suelo de los bosques tropicales al principio de la estación lluviosa se explica por la presencia de muchas semillas quiescentes que se diseminaron durante la estación seca.
La germinación de las semillas con frecuencia tiene lugar en la superficie del suelo, por lo que el equilibrio entre la ganancia de humedad del suelo y su pérdida por transpiración a la atmósfera determina el momento en que la semilla se satura de humedad y comienza a germinar. Algunas veces este equilibrio mejora cuando las semillas están parcialmente enterradas en la hojarasca.
Una vez que la semilla ha completado su desarrollo se inician los cambios que darán lugar al establecimiento del reposo en las semillas (cuadro 3). Este reposo o reducción del metabolismo se denomina quiescencia cuando la causa de que no ocurra la germinación es fundamentalmente la falta de agua, como es el caso de las semillas almacenadas en condiciones artificiales, por ejemplo un frasco con frijoles en la alacena o las semillas que permanecen en los frutos unidos a la planta madre por largo tiempo. En cambio, el reposo de las semillas se denomina latencía cuando la semilla no germina a pesar de encontrarse en un lugar óptimo en cuanto a la temperatura y la humedad. Las causas por las que no germinan pueden deberse a la existencia de un periodo cronológicamente regulado de interrupción del crecimiento y de disminución del metabolismo durante el ciclo vital. Ésta es una estrategia adaptativa de supervivencia frente a condiciones ambientales desfavorables que se presenta en algunos seres vivientes. En las plantas superiores puede existir latencia o interrupción del crecimiento en el tejido meristemático, por ejemplo en las yemas de crecimiento de las ramas, así como en las semillas. El establecimiento de la latencia está regulado por factores hereditarios que determinan los mecanismos fisiológicos endógenos de las plantas, los cuales interactúan con factores del ambiente en el que las plantas crecen; esto da lugar, a la larga, a cambios evolutivos en las plantas. Entre las condiciones más importantes del ambiente se encuentran las variaciones climáticas de temperatura y humedad, las variaciones microclimáticas derivadas de aspectos fisiográficos y bióticos, como la calidad espectral de la luz y el termoperiodo, así como las características específicas del lugar a las que las plantas se han adaptado para establecerse y crecer. Las variaciones micro y macroclimáticas, así como las condiciones hormonales y nutricionales de la planta progenitora tienen gran influencia en el establecimiento de la latencia de sus semillas durante su desarrollo, por lo cual pueden existir variaciones entre cosechas de semillas de una especie, según la época y el lugar de producción.
UADRO 3. Pasos en el desarrollo del reposo en las semillas
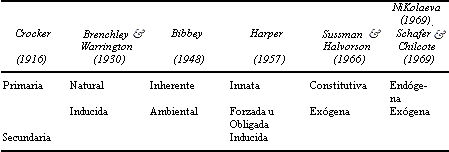
Se han definido varios tipos de latencia que en síntesis se presentan en el cuadro 4. Nos ajustaremos aquí a la clasificación que el ecólogo británico John Harper presentó en 1957, que toma en cuenta principalmente el comportamiento de este mecanismo fisiológico en la naturaleza. Existen clasificaciones más modernas, o con otro enfoque, que pueden consultarse en diversas obras.
CUADRO 4. Clasificación de los tipos de latencia de acuerdo
con diferentes autores citados por Koslowski en 1972
Se presenta en el momento en el que el embrión cesa de crecer (cuando la semilla aún está en la planta madre) y continúa hasta que el impedimento endógeno cesa, en ese momento las semillas están en condiciones de germinar en cuanto se presentan las condiciones ambientales adecuadas. La presencia de inhibidores químicos de la germinación en el embrión o la inmadurez de éste son probablemente las causas principales de esta latencia. La duración de la latencia innata es muy variable según la especie, en algunos casos, incluso puede variar entre las semillas de un mismo individuo. Algunos experimentos parecen indicar que en ciertas semillas tropicales existen procesos comparables a la posmaduración, característica de muchos árboles de climas templados, cuyas semillas sólo germinan después de haber transcurrido el invierno, de haber sido expuestas a bajas temperaturas en el laboratorio o mediante la aplicación de hormonas vegetales, como el ácido giberélico o almacenándolas en frío.
Latencia inducida o secundaria
Este tipo de latencia se produce cuando las semillas están en condiciones fisiológicas para germinar y se encuentran en un medio que presenta alguna característica muy desfavorable, como poco oxígeno, concentraciones de CO2 mayores a las de la atmósfera, temperatura alta, etc., lo que puede producir alteraciones fisiológicas reversibles en las semillas. En estos casos, las semillas pueden caer en un estado de latencia secundaria en el que ya no pueden germinar a pesar de continuar vivas. En algunos casos este tipo de latencia se rompe por medio de un estímulo hormonal. Algunas veces la latencia inducida también puede sumarse a otros tipos de latencia o sustituirlos.
En la naturaleza esta latencia se presenta en semillas aptas para germinar en condiciones adecuadas de humedad y temperatura media, es decir, adecuadas al hábitat que ocupan, pero que continúan latentes por falta de luz, requerimientos especiales de temperatura, oxígeno o de otro factor. Esta latencia está controlada por las condiciones físicas del ambiente que rodea a la semilla, y se presentan en aquellas que se encuentran en el suelo y que germinan sólo después de una perturbación que modifique el régimen lumínico o el contenido de oxígeno.
Germinación retardada por una testa impermeable
Muchas plantas producen semillas cuyo tegumento externo es duro impermeable al agua o a los gases, e incluso el micrópilo está provisto de una barrera que impide la penetración de agua al embrión. Esta característica es frecuente en varias familias de plantas, particularmente en las fabáceas o leguminosas, las malváceas y bombacáceas.
En el suelo del bosque la cubierta de la semilla gradualmente se vuelve permeable por intemperismo, degradación microbiana, factores del suelo como las saponinas o por el efecto de fluctuaciones de temperatura, y va germinando poco a poco. Este mecanismo de latencia pasiva es particularmente frecuente en los bosques tropicales secos, y puede haberse originado como un mecanismo de persistencia de las semillas en el suelo a lo largo de la estación desfavorable de crecimiento.
Frecuentemente se dice que el tránsito a través del tubo digestivo de animales es uno de los factores principales que rompen este tipo de latencia entre las semillas que son dispersadas por animales. Sin embargo, muchas de las especies que presentan testa dura no son ingeridas por animales; otras, aunque sí sean ingeridas, son destruidas o no muestran mucha diferencia en su germinación antes y después de haber sido ingeridas por animales.
Las altas temperaturas también pueden romper los tegumentos. Esto ocurre frecuentemente durante los incendios o las quemas en los terrenos de cultivo, sobre todo en los trópicos. Los tegumentos también pueden cambiar su estructura después de ser expuestos a la insolación directa por periodos prolongados.
Es probable que muchas de las semillas resistentes al calor presenten tegumentos impermeables al agua, ya que las semillas contienen enzimas, nucleoproteínas y otras sustancias que se desnaturalizan con facilidad con el calor; estos compuestos son menos lábiles cuando están deshidratados, por lo que una testa impermeable impide que la semilla se embeba y por lo tanto queda protegida durante las quemas.
GERMINACIÓN CONTROLADA POR EL AMBIENTE
Los cambios que ocurren durante la germinación comprenden procesos metabólicos que se producen en estrecha relación con la temperatura, y su efecto se expresa en la capacidad germinativa o en la velocidad de germinación. Las temperaturas cardinales de la germinación son óptima, máxima y mínima, y el intervalo térmico en el que las semillas germinan son características sujetas a la selección natural. Por esto, con frecuencia se presentan como adaptaciones muy claras a los hábitat en los que las plantas se desarrollan, y hay diferencia entre las especies, incluso entre distintas poblaciones de la misma especie de acuerdo con su distribución geográfica. En zonas templadas o frías, en muchos casos se ha visto que las semillas sólo germinan en los intervalos de temperaturas que caracterizan precisamente a las épocas del año más favorables o adecuadas para el establecimiento de las plantas.
Muchos fisiólogos han considerado que las temperaturas cardinales son parámetros fisiológicos muy útiles en el estudio de la germinación. Sin embargo, pueden presentarse dificultades al tratar de fijar en forma precisa las temperaturas cardinales de una especie, ya que con frecuencia éstas varían según el estado de maduración de las semillas o son dificiles de detectar debido a la lentitud de la germinación a ciertas temperaturas.
Además, en algunas plantas existen respuestas bimodales ante la temperatura, es decir, dos temperaturas óptimas separadas por temperaturas menos favorables. Normalmente la búsqueda de las temperaturas cardinales de las especies se ha realizado utilizando estufas o cámaras a temperatura constante. El uso de una placa de gradiente térmico también es muy útil para ese propósito, así como para conocer el intervalo térmico de germinación.
El estudio de las temperaturas cardinales o del intervalo térmico de germinación es insuficiente para conocer la respuesta germinativa de especies que producen semillas cuya germinación se ve favorecida por una alternancia de temperatura, como la que se produce por el calentamiento del suelo soleado durante el día. Las semillas que responden a este cambio ambiental pueden presentar diversos mecanismos para detectar este factor, por ejemplo, la presencia de una testa impermeable que se hace permeable al calentarse, o la existencia de un mecanismo químico endógeno que sólo puede activar el proceso de germinación cuando ocurren fluctuaciones de temperatura.
En el interior de los bosques la temperatura en la superficie del suelo se mantiene relativamente constante durante el día y la noche, en tanto que en los lugares abiertos ésta puede fluctuar hasta más de 10°C cada día. Esta forma de control de la germinación es frecuente en regiones tropicales de baja altitud, cubiertas por vegetación perennifolia densa, donde el suelo es prácticamente isotermal a lo largo del año; en tanto que las temperaturas de los suelos desnudos de vegetación presentan una fluctuación diaria muy marcada en la capa superficial durante los días despejados. Seguramente en esos ambientes las temperaturas cardinales y el intervalo térmico de germinación no tienen las mismas presiones de selección que se observan en regiones templadas o frías, donde la estacionalidad de la temperatura es determinante para que ocurra la germinación; en regiones isotermales son más bien las condiciones microclimáticas las que determinan el establecimiento de las plantas. Obviamente, este tipo de selección también es posible en cualquier otra región, incluyendo las templadas.
Muchos fisiólogos han experimentado con termoperiodos en pruebas de germinación y en muchas especies se ha observado que una o varias alternancias de temperatura pueden favorecer o disparar la germinación. El efecto de la alternancia de temperatura parece tener relación con la hidratación de las semillas, pues la escarificación de éstas es suficiente para permitir la germinación a una temperatura constante. Los cambios fisicoquímicos producidos por el termoperiodo en las semillas, que conducen a la desaparición de la latencia, son de muy diversa naturaleza según la especie. En general se piensa que la fluctuación de temperatura permite la activación de ciertas enzimas y hace permeables a algunas membranas, lo que finalmente trae consigo el desencadenamiento de la germinación.
A pesar de que en condiciones naturales el termoperiodo que se produce en la superficie del suelo tiene influencia en la germinación de ciertas semillas que regulan su germinación gracias a él, no siempre es necesario para la germinación de otras especies, aun cuando pertenezcan a comunidades en las que regularmente existe un marcado termoperiodo a nivel del suelo.
El fuego es un factor ambiental natural que el hombre ha favorecido, pues al parecer los fuegos naturales son menos frecuentes que los inducidos. Actualmente el fuego ha actuado y está actuando cada vez en mayor escala sobre el medio ambiente debido a las prácticas características de la agricultura nómada y a que los incendios forman parte del mantenimiento de pastizales y campos de cultivo colindantes con la vegetación natural.
Se ha observado que diversas especies vegetales poseen semillas capaces de tolerar altas temperaturas y aumentar su germinación bajo los efectos de éstas. Especies con estas características son frecuentes en las sabanas tropicales y los pastizales templados. En ciertos bosques de coníferas y otras zonas, en las que los fuegos son frecuentes, la existencia de incendios periódicos ha sido un importante factor de selección sobre ciertas características anatómicas y fisiológicas de las plantas, y contribuyen en gran medida a la estabilidad y mantenimiento de este tipo de comunidades.
Es muy probable que todas las semillas resistentes al calor presenten latencia de tipo tegumentario, debido que su tegumento es impermeable al agua, ya que las semillas hidratadas son más susceptibles a sufrir daños al calentarse. La estructura del tegumento cambia con el calentamiento húmedo o seco a temperaturas variables, según la especie, lo cual permite la entrada del agua o los gases al interior de la semilla. Este tipo de latencia es particularmente frecuente entre las fabáceas, malváceas, y otras plantas que germinan abundantemente en el suelo de bosques recientemente incendiados o de otras comunidades, como el chaparral de California, ciertos pinares y las sabanas tropicales. Los matorrales del Cabo, en Sudáfrica, llamados finbos, de los cuales proceden plantas de ornato tan conocidas como los geranios, están tan adaptados a los fuegos periódicos que incluso se vende comercialmente un compuesto preparado con el humo de la combustión de la materia orgánica de ese matorral para facilitar la germinación de esas semillas a aquellas personas interesadas en propagar plantas de esa comunidad. También en el chaparral de California y Baja California Norte se ha visto que el humo puede estimular la germinación de algunas especies.
El estudio del efecto de la luz sobre la germinación se ha mantenido en el primer plano del interés de los fisiólogos y de los ecofisiólogos durante mucho tiempo. Actualmente, la cantidad de información disponible en este campo es muy grande. La fotoinducción o fotoinhibición de la germinación es uno de los casos más claros del control de un proceso fisiológico por un factor ambiental.
No se sabe cuántas especies de plantas superiores presentan semillas fotoblásticas (germinación regulada por la luz), ya que la fisiología de las semillas de la gran mayoría de las plantas no ha sido investigada; sin embargo, existen evidencias que indican que el porcentaje de especies con semillas fotoblásticas es particularmente alto entre las plantas anuales.
Son tres las principales bandas del espectro lumínico que tienen acción sobre la germinación, y corresponden a la franja de 660 nanómetros (rojo), 730 nanómetros (rojo lejano) y la luz comprendida entre 400 y 500 nanómetros (azul), aunque con efectos mucho menos claros. Tanto el rojo como el rojo lejano son absorbidos por un compuesto denominado fitocromo, que es una cromoproteína que actúa como sensor. Este pigmento en su forma activa es inductor de la germinación e interviene en procesos de permeabilidad, activación de enzimas y expresión genética. La conversión del fitocromo inactivo (Pr) a fitocromo activo (Pfr) por lo general se lleva a cabo bajo el efecto de la luz roja, y la reacción opuesta ocurre bajo el efecto del rojo lejano. Estas dos formas del fitocromo corresponden a cada uno de sus picos de absorción de luz. Esta reacción de conversión en ambos sentidos está relacionada con la inducción y la inhibición de la germinación, y puede ser modificada o controlada por otros factores ambientales como la temperatura o el termoperiodo. La intensidad de la luz, el fotoperiodo y la cantidad de rojo en relación con el rojo lejano presente (denominada relación R:RL) modulan la respuesta de las semillas a la luz a través de este pigmento. La cantidad de fitocromo activo presente en una semilla en el momento de su liberación determina si ésta puede germinar en la oscuridad o si requerirá luz para iniciar el proceso.
En los lugares cubiertos de vegetación la luz que llega al suelo es pobre en rojo y rica en rojo lejano, mientras que en los lugares abiertos ambas longitudes de onda llegan en igual proporción, debido a que la luz no ha sido filtrada por un dosel vegetal. El mecanismo de captación de la luz del fitocromo es sensible principalmente a la calidad de luz, lo que permite a las semillas detectar, en particular, la proporción de R:RL que llega al suelo. De esta manera, algunas semillas de plantas de sol pueden permanecer latentes cuando son dispersadas hacia lugares que están densamente cubiertos de vegetación y que no serían propicios para su establecimiento. La hojarasca que cubre al suelo también modifica la relación R:RL, así como otras propiedades fisicas del suelo.
Este tipo de latencia regulada por la luz es muy común en semillas que permanecen latentes en la oscuridad, enterradas en el suelo, hasta que son exhumadas durante las prácticas agrícolas. Cuando estas semillas alcanzan la superficie del suelo quedan expuestas a la luz y germinan.
En las semillas pequeñas la conversión del fitocromo normalmente se realiza a muy bajas intensidades de luz; sin embargo, la temperatura y el fotoperiodo tienen influencia directa en el resultado final del proceso, expresado en porcentaje y velocidad de germinación.
En ocasiones el proceso fisiológico que induce la germinación necesita la luz, y la germinación total sólo se obtiene cuando la luz y la temperatura actúan simultáneamente. En algunos casos, la temperatura puede provocar que las semillas germinen en la oscuridad, aun cuando generalmente requieren luz. La luz y la fluctuación de temperatura interactúan de muchas maneras para determinar la germinación de las semillas presentes en el suelo en el momento en que las condiciones para el establecimiento de las plantas son las óptimas.
La regulación de la germinación producida por las fluctuaciones de temperatura, la calidad de la luz incidente o alguna combinación de ambos factores es común en hierbas y plantas colonizadoras. Los sensores ambientales de estas semillas detectan cambios en su ambiente que les indican la aparición de condiciones favorables para la germinación y establecimiento de las plantas. Estos cambios se dan cuando la cubierta de plantas se destruye, cuando desaparece la capa de hojarasca o cuando las semillas son desenterradas por algún disturbio, producto de un fenómeno natural o de la perturbación por el hombre.
En las zonas templadas las semillas de muchas especies responden al fotoperiodo, y se han encontrado casos en los que germinan mejor o solamente en días largos o en fotoperiodos especiales, previniendo así la germinación durante la época desfavorable.
En condiciones de humedad adecuadas, la temperatura y el fotoperiodo de los bosques tropicales es favorable para la germinación de la mayoría de las semillas; no obstante, algunas plantas producen semillas que son sensibles a las condiciones ambientales, y que retrasan la germinación hasta que se presentan ciertas condiciones específicas de luz o temperatura en su medio.
Temporalidad de la germinación
La falta de humedad limita muchas veces la germinación; sin embargo, independientemente de ésta, la existencia de un periodo de latencia es bastante común en muchas plantas. El flujo de la germinación en una cosecha puede ocurrir de varias formas: 1) toda la cosecha de semillas permanece sin germinar por semanas, meses o incluso años; 2) parte de la cosecha germina pronto y otra parte permanece latente por más tiempo; 3) la germinación es gradual y las semillas de una cosecha van germinando a lo largo de varias semanas o meses; 4) se presentan dos o más eventos de germinación separados en el tiempo dentro de la misma cosecha, y finalmente 5) las semillas germinan simultáneamente poco después de su llegada al suelo. Las causas de estos patrones de germinación retardada son variables, complejos y poco conocidos para muchas especies silvestres. Dos de las causas que los determinan son: 1) la permeabilidad limitada o diferencial de las semillas al agua o al oxígeno; 2) la existencia de una latencia endógena debida a la inmadurez del embrión, que toma tiempo en desarrollarse, o bien al equilibrio inicial de hormonas de crecimiento; por ejemplo, el ácido abscísico y las giberelinas, ya que la primera es inhibidora de la germinación y la segunda la induce.
Cada comunidad vegetal presenta mecanismos de germinación característicos que responden al efecto de la selección natural inducida por las condiciones ambientales predominantes sobre la naturaleza y fisiología de las semillas. Debido a la gran complejidad que presenta cada uno de los muchos ambientes poblados por plantas sobre la corteza terrestre, tomaremos sólo tres condiciones contrastantes para ejemplificar las adaptaciones del proceso de germinación predominantes en cada caso.
1) Los desiertos. Se caracterizan por presentar un periodo muy corto (que no todos los años se presenta) en el cual la humedad y el tiempo son adecuados para permitir la germinación y el establecimiento de plantas. Por este motivo en los desiertos predominan las plantas perennes de larga vida, que tienen muchas oportunidades de reproducirse a lo largo de su existencia. En la mayoría de los años todas las semillas perecen. Sin embargo, basta que ocasionalmente se presente algún año muy favorable para que cierto número de semillas logre sobrevivir y establecer nuevos individuos. Estas semillas, más que tener mecanismos de latencia complejos, presentan simplemente una quiescencia que les permite germinar en cuanto hay humedad. Con mucha frecuencia estas plantas presentan también reproducción vegetativa eficiente y a veces predominante como mecanismo de propagación. Por el contrario, las plantas anuales, o efímeras del desierto, producen semillas con latencias complejas, con frecuencia polimórficas, que impiden la germinación de toda la cosecha anual en un mismo año, de manera que algunas semillas pueden sobrevivir por varios años en el suelo y germinar a diferentes tiempos, mientras que otras germinan en cuanto hay humedad. Esto garantiza que haya algunas semillas que germinen en el año en que las condiciones son más favorables y la humedad suficientemente prolongada, para que las plantas se desarrollen hasta producir semillas que renueven su disponibilidad en el suelo.
2) Los bosques templados. En este caso hay dos posibilidades predominantes. Si la fructificación y diseminación de las semillas ocurre en otoño, éstas deben sobrevivir latentes en el suelo hasta la primavera siguiente, cuando las condiciones de temperatura son adecuadas para el establecimiento. Estas semillas suelen tener latencias endógenas que impiden su germinación inmediata y un mecanismo de maduración a baja temperatura que las prepara para germinar en la primavera. Si los frutos maduran en invierno o en primavera y las semillas son diseminadas en primavera, o al principio del verano, la germinación suele ser inmediata, por lo que la nueva planta aprovecha toda la estación de crecimiento, alcanzando un desarrollo que le permite sobrevivir el invierno siguiente.
3) La selva tropical húmeda. En este caso hay condiciones de alta humedad y temperatura continuas combinadas con una gran abundancia de organismos que utilizan las semillas como alimento, de manera que la mayoría de las especies de semillas tiende a germinar rápidamente, sin presentar mecanismos de latencia profundos. Esto favorece que las semillas escapen de sus depredadores y formen plántulas menos vulnerables a ellos. Las excepciones son las semillas de testa dura, que tienen que sufrir un proceso de escarificación para germinar, y las semillas de plantas colonizadoras de claros, que suelen presentar mecanismos de latencia impuesta que les impiden germinar a menos que haya un cambio en las condiciones ambientales que caracterizan a la selva densa, como las típicas aperturas en la vegetación.
Los estudios de ecología y fisiología de las semillas de plantas tropicales y subtropicales son escasos, y se refieren a una fracción insignificante de las especies que componen esas comunidades. La escasa información ha sido condensada en algunos trabajos de revisión que se han publicado durante la última década sobre temas como la dispersión de semillas, los bancos de semillas en el suelo, la germinación, la ecología de semillas, el análisis sobre el papel ecológico del tamaño de semillas y finalmente la relevancia de las semillas en el manejo de bosques.
Como consecuencia de la enorme diversidad de especies que caracteriza al trópico, combinada con la variedad de historias filogenéticas de sus componentes, las semillas poseen una rica diversidad que se manifiesta en aspectos tan variados como: la estacionalidad de la producción, el volumen de la cosecha de semillas, el número de semillas por fruto, el tamaño, la forma, la anatomía y morfología, el contenido de humedad, la naturaleza de sus reservas y de sus compuestos secundarios, su fisiología y su comportamiento ecológico. No hay una regla para describir las características de estas semillas y tampoco es posible establecer generalizaciones sobre su estructura y comportamiento; sin embargo, se ha dicho con frecuencia que los mecanismos de latencia y germinación más frecuentes en los bosques tropicales húmedos son menos complejos que aquellos que existen en regiones con clima muy estacional, así como entre las plantas anuales.
El banco de semillas del suelo está formado por las semillas viables no germinadas
presentes en éste, ya sea enterradas, depositadas sobre la superficie o mezcladas
en la capa de hojarasca y humus. Frecuentemente hay gran cantidad de semillas
latentes en la mayoría de los suelos, su número depende de factores como la
historia, la diversidad y la dinámica de la vegetación que cubre cada suelo.
Entre las muchas contribuciones que el naturalista británico Charles Darwin
hizo a la biología está el hecho de que quizá fue el primero que escribió acerca
del banco de semillas del suelo, en 1859:
El tiempo que las semillas de una especie de planta determinada permanecen
latentes en el suelo antes de germinar, morir o ser atacadas por parásitos o
depredadores está determinado por factores fisiológicos innatos y por las condiciones
ambientales del lugar. Los bancos de semillas de diferentes especies varían
en duración, de temporales a persistentes. En los primeros su duración corresponde
al periodo de diseminación de una cosecha de semillas, en los segundos las semillas
latentes de una especie siempre se encuentran en el suelo. Las semillas recién
diseminadas tienen tres destinos diferentes en el suelo, que son: 1)
la pronta germinación después de su llegada al suelo; 2) un periodo
de latencia que dura hasta que las condiciones ambientales son apropiadas para
la germinación, y 3) la muerte por envejecimiento natural o por la acción
de parásitos o depredadores. En el primer caso se forman solamente bancos de
semillas relativamente efimeros correspondientes a la época de diseminación
de las semillas de cada especie. En el segundo caso se forma un reservorio de
semillas de duración más larga que permanece en el suelo hasta que un cambio
ambiental interrumpe la acción del mecanismo que induce la latencia. Otra situación
en la que se forman bancos de semillas permanentes se debe a la llegada continua
al suelo de semillas de especies que presentan una ininterrumpida producción
(figura 8).
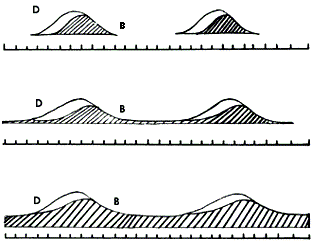
Figura 8. Persistencia de las semillas en el suelo. La parte blanca de la
curva corresponde a la diseminación de las semillas (D); la parte achurada corresponde
a la persistencia de semillas en el suelo (B): banco efímero (arriba), banco
de persistencia intermedia (en medio), banco persistente (abajo).
Longevidad de las semillas en el suelo
Las semillas desempeñan cuatro funciones importantes para la vida y la continuidad en el tiempo de las poblaciones vegetales. No todas son igualmente importantes para las diferentes especies de plantas, ya que han sido moduladas por la selección natural, por las condiciones que prevalecen en cada comunidad en interacción con las características de la historia de vida de cada población de plantas. Estas funciones son la reproducción, la diseminación dentro de la misma comunidad, la expansión hacia nuevos territorios o hacia otros hábitat, y finalmente la sobrevivencia del germoplasma en condiciones meteorológicas o ambientales desfavorables para el crecimiento. La capacidad de las semillas para permanecer vivas y viables, sin germinar por diferentes periodos en el suelo de la comunidad a la que pertenecen, se conoce como longevidad ecológica. Por otra parte, la capacidad para permanecer viables en una condición óptima de almacenamiento artificial por algún tiempo se conoce como longevidad potencial.
La longevidad ecológica varía mucho entre las diferentes especies y tiene estrecha relación con las características del hábitat de las especies. En bosques húmedos las semillas de los árboles de la vegetación madura tienden a germinar rápidamente y su longevidad ecológica es breve; las frecuentes temperaturas altas y el ambiente predominantemente húmedo de estos bosques han seleccionado características de las semillas que conducen principalmente a la realización de las funciones de reproducción y diseminación. Muchas de estas semillas tienen al momento de la dispersión un alto contenido de humedad y un metabolismo activo que se expresa en una tasa respiratoria ininterrumpida durante el periodo de quiescencia.
En bosques tropicales más estacionales, muchas de las semillas producidas durante la época húmeda deben sobrevivir en el suelo durante la estación seca para iniciar su crecimiento al principio de la siguiente estación húmeda, por lo que su longevidad ecológica es mayor y el periodo de quiescencia suele caracterizarse por una deshidratación profunda y la total interrupción del metabolismo. También es posible encontrar longevidades ecológicas más largas en aquellas plantas que se establecen en lugares de aparición aleatoria en la comunidad, como los claros naturales y los disturbios en la vegetación madura. Estas especies suelen presentar mecanismos de espera que incluyen una longevidad ecológica más larga, como la que existe en muchas hierbas y malezas de lugares perturbados por el hombre.
La longevidad potencial de las semillas está de alguna manera relacionada con la longevidad ecológica, ya que las semillas en forma natural sufren una deshidratación profunda, la interrupción del metabolismo y la posibilidad de sobrevivir latentes por algún tiempo; estas mismas cualidades pueden permitirles resistir un almacenamiento prolongado en condiciones ambientales controladas.
¿QUÉ TAN LONGEVAS SON LAS SEMILLAS?
Existe una extensa bibliografía sobre la longevidad de las semillas e informes de variable credibilidad acerca de casos extremos de longevidad. Una revisión extensa al respecto fue hecha por Lela Barton en 1961. En su libro se mencionan las famosas semillas de trigo, cebada y chícharo que fueron encontradas supuestamente viables en tumbas egipcias y otros casos de longevidad extrema de semillas de plantas cultivadas. La validez de estos informes ha sido puesta en duda pues se han realizado experimentos almacenando estas especies, a las temperaturas que privarían en tales condiciones, y nunca se han podido conservar viables por más de unos cuantos años. Sin embargo, otras investigaciones acerca de semillas encontradas en sitios arqueológicos o en edificios antiguos, formando parte de adobes o en recipientes de barro, son más convincentes, como el caso de algunos adobes de edificios coloniales de California y norte de México. Un análisis amplio de este tema se presentó en los libros de los estadunidenses Lela V. Barton en 1961 y David A. Priestley en 1986. En los siguientes párrafos se verá con mayor profundidad algunos de los aspectos más importantes de este interesante problema, que tiene implicaciones tanto en ciencia básica como en la aplicada a la agricultura y a la forestería, y en la actualidad a la conservación del germoplasma vegetal.
Los informes sobre la longevidad de semillas de diferentes especies silvestres y cultivadas pertenecen a tres categorías, dependiendo de la condición de almacenamiento en que estaban las semillas cuya viabilidad se determinó:
1) Almacenamiento en condiciones artificiales.
2) Almacenamiento en condiciones seminaturales (en el suelo).
3) Almacenamiento natural en el banco de semillas del suelo.
El almacenamiento en condiciones artificiales o controladas subóptimas ha sido fuente de numerosos informes sobre la longevidad de semillas. Las condiciones de almacenamiento más frecuentemente encontradas son: ejemplares de herbarios, botellas o cajas en gavetas de laboratorio, almacenes de semillas en condiciones ambientales no reguladas finamente y otros tipos de almacenamiento consciente o accidental en ambientes que no propician la expresión completa de la longevidad potencial de las semillas. Por ejemplo, los datos que se obtienen del herbario, aunque son de gran interés como fuente de información, no indican qué semillas tienen realmente una longevidad larga y cuáles no la tienen, sino cuales fueron capaces de sobrevivir a los tratamientos que sufren los ejemplares y a las condiciones de las gavetas de herbario.
El almacenamiento de semillas en el suelo, en recipientes parcialmente aislados, es una condición seminatural para determinar la longevidad que ha sido objeto de varios ensayos en diferentes regiones del mundo. Quizá el experimento más famoso de este tipo sea el del doctor Beal, quien enterró semillas de 19 arvenses (plantas silvestres de campos de cultivo) y de un trébol cultivado en un terreno de East Lansing, Michigan. Las semillas se pusieron en frascos llenos de arena en la superficie. El experimento fue diseñado para durar más de 100 años y aún quedan varios frascos. Después de 100 años persistían semillas viables de dos especies de hierbas. Existen otros experimentos de esta naturaleza, como los de Duvel en Virginia y los Dorph-Petersen en Dinamarca, entre otros.
La longevidad de las semillas en el lugar al que fueron naturalmente diseminadas es mucho más difícil de determinar, y sólo existen informes de condiciones muy especiales. Por ejemplo Porsild y colaboradores, en 1967, encontraron en el permafrost, formado en el pleistoceno, semillas de una leguminosa que fueron capaces de germinar normalmente. El danés Odum en 1965 encontró semillas aún viables de algunas plantas arvenses bajo piedras de construcciones de la era de los vikingos (año 1000 aproximadamente). Este mismo autor ha fechado la antigüedad de bancos de semillas de varios lugares de Dinamarca, y ha encontrado evidencias de una longevidad de varias décadas en bancos de semillas. Existen otras evidencias de considerable longevidad del banco de semillas del suelo para algunas regiones en zonas templadas. En los trópicos por lo general la longevidad ecológica es más corta. Existen casos en que las semillas caen al suelo ya verdes, desprovistas de su cubierta, cargadas de humedad y prácticamente germinando, como el jinicuil. Si estas semillas no encuentran un medio propicio para germinar pronto, mueren.
Estructura y composición de las semillas
La diversidad de formas, tamaños, estructura anatómica e histológica y manifestaciones de los procesos fisiológicos es enorme, por lo que también lo es la variabilidad en la longevidad potencial y ecológica que estas características determinan. Siendo imposible en este breve espacio dar aunque sea un pequeño bosquejo de la variabilidad morfológica de las semillas, remitimos a los lectores a la lectura del botánico mexicano Aníbal Niembro, publicada en 1984, como fuente de información inicial sobre la ontogenia y la estructura de las semillas. Por lo que respecta a la información acerca de la fisiología, la diversidad de mecanismos que determinan la latencia o letargo, su interrupción y la germinación propiamente dicha es también muy vasta. Una buena revisión relativamente reciente sobre este tema es la publicada por Bewley y Black en 1994. A pesar de esta gran diversidad y complejidad de estructuras y funcionamientos es posible hacer algunas generalizaciones y señalar cuáles son las características de las semillas que más directamente afectan su longevidad potencial, ya que la longevidad ecológica, o sea la que se da en la comunidad natural, no solamente depende de la estructura de las semillas, sino de factores externos que pueden variar de un lugar a otro y afectar de diferente manera a una misma especie de semillas. Estas características intrínsecas son: la dureza e impermeabilidad de la testa o cubiertas de la semilla; el contenido de agua inicial con el que se disemina la semilla y el que se puede lograr disminuir por deshidratación sin matarla; la tolerancia a la temperatura de congelación del agua, ya que los cristales de hielo de ésta pueden alterar las células de la semilla; la naturaleza de la latencia, la tasa metabólica o respiración mínima o interrumpida que se pueda alcanzar sin matarla; el tipo de reservas y su propensión al deterioro químico; el tipo de compuestos secundarios (sustancias que produce la semilla que pueden evitar la depredación o el parasitismo de éstas) y su ubicación dentro de las células de la semilla; la disposición del agua subcelular, o sea la que está unida a macromoléculas como proteínas, que forman las células; la composición de los lípidos (grasas) de las membranas celulares; la resistencia a la invasión de microorganismos y la resistencia al deterioro del material genético (cuadros, 5 y 6).
UADRO 5. Estructura y composición de las semillas y su relación
con la duración de la viabilidad
UADRO 6. Factores relacionados con la longevidad de las semillas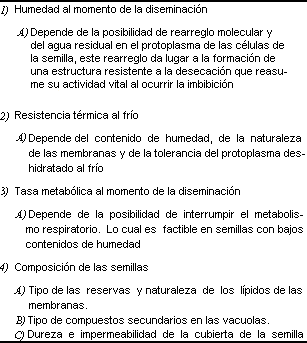
En el medio natural las semillas con testa impermeable y bajo contenido de humedad suelen ser muy longevas, como sería el caso de la especie de leguminosa encontrada en el permafrost, que se mencionó en párrafos anteriores, pero también pueden serlo algunas semillas de latencia fotorregulada que plenamente hidratadas pueden conservarse enterradas en suelos con una alta humedad estable, en condiciones de oscuridad total. Éste es el caso de las semillas de plantas arvenses de larga duración en el suelo. Su requerimiento de luz para germinar las mantiene latentes por mucho tiempo en condiciones de enterramiento, como las de muchas malezas que producen semillas fotoblásticas.
En los almacenamientos a temperatura ambiente las semillas con testa dura de las leguminosas, malváceas, ninfáceas, etc., son las más longevas. En los almacenamientos óptimos a bajas temperaturas muchas familias como las crucíferas, las compuestas, las cariofiláceas, las solanáceas, las gramíneas y otras han demostrado tener una longevidad potencial extremadamente larga.
En toda comunidad natural es posible encontrar especies con semillas de longevidad potencial muy larga y otras de longevidad corta, ya que la longevidad ecológica (y como consecuencia la potencial) está relacionada con varias características de las plantas que las producen (cuadro 7), como son: la historia de vida de la planta; la estacionalidad climática; la distancia temporal entre el periodo de fructificación y diseminación y el de germinación y establecimiento; el grado de discontinuidad en el hábitat, o sea que las plantas requieran para establecerse condiciones ambientales especiales que sólo aparecen esporádicamente en la comunidad en la que viven; el agente dispersor de las semillas, y los niveles de estrés ambiental y de presión de depredadores que sufre la semilla en el suelo, antes y durante la germinación. Debido a la complejidad de todos estos factores se mencionarán sólo algunos ejemplos.
UADRO 7. Algunos factores ecológicos relacionados con la longevidad
de las semillas
Con respecto a la historia de vida, las plantas anuales, que tienen una estación de crecimiento corta y que sobreviven sólo en forma de semilla el periodo desfavorable del año, suelen producir semillas con una longevidad ecológica y potencial más larga que muchas de las especies perennes que comparten su hábitat, para las que existen varias oportunidades de reproducirse durante su vida.
Las semillas cuyo periodo óptimo de germinación está separado varios meses de la época de diseminación, como el caso de las especies que pueblan zonas del planeta con un clima estacional muy marcado, suelen tener longevidades ecológica y potencial más largas que aquellas que en el medio natural tienden a germinar de inmediato porque encuentran condiciones permanentemente favorables al caer al suelo. Con respecto a la discontinuidad del hábitat, podemos mencionar a las plantas costeras que se diseminan por el agua de mar, que al tener estructuras de resistencia al agua marina y capacidad de flotación pueden tener longevidades ecológica y potencial muy prolongadas, en este segundo caso aun a la temperatura ambiente. Esto se debe a que estas plantas tienen una testa o cubierta protectora impermeable muy resistente que impide la entrada de agua salada a la semilla.
Las semillas de diseminación anemócora suelen tener bajos contenidos de humedad, ya que esta característica reduce el peso del propágulo, y como la baja humedad propicia la larga longevidad potencial frecuentemente se deshidratan y almacenan con facilidad. Tal es el caso de algunas especies tropicales de selvas húmedas como la balsa, las ceibas, el cedro rojo y la teca, que con frecuencia son las únicas semillas disponibles en los bancos de semillas del trópico húmedo.
En las selvas tropicales húmedas es frecuente que la germinación ocurra inmediatamente después de la diseminación. El estrés ambiental al que se ven sometidas las plántulas por la carencia de luz y a veces de nutrientes ha favorecido a las plántulas grandes, y por lo tanto a las semillas grandes. Éstas son fácilmente depredadas en este ambiente. Este conjunto de factores ha determinado que, por lo general, la longevidad ecológica y potencial de las semillas de estas comunidades sean muy breves, con excepción de las plantas que germinan y se establecen en los claros, o sea, que tienen un hábitat de establecimiento discontinuo en el espacio y en el tiempo.
La mayoría de las plantas presentan variabilidad genética y fenotípica en distintos lugares de su área de distribución. Esta variabilidad debe tomarse en consideración cuando se manejan semillas de diferentes localidades con cualquier propósito. En plantas procedentes de diferentes localidades pueden haber evolucionado características específicas para lidiar con los factores limitantes locales y con las interacciones bióticas específicas del sitio, lo cual las hace inapropiadas para crecer en un nuevo conjunto de condiciones ambientales.
Dentro de una población de plantas existe siempre gran variación genética que debe ser preservada cuando se toman muestras de semillas con diferentes propósitos; por ejemplo, hay heterogeneidad natural en las características fisiológicas de las semillas de diferentes individuos, por lo que cuando se usan semillas de uno o pocos individuos para estudiar la longevidad o los patrones de germinación, los resultados obtenidos a veces no corresponden al comportamiento más común o medio de la población.
Mantener la valiosa variabilidad genética es una parte importante del almacenamiento de semillas, por lo que estas semillas deben ser recolectadas preferentemente de poblaciones silvestres, ya que cuando las semillas se colectan de poblaciones pequeñas, ya sea manejadas o cultivadas, ocurre una indeseable selección de aquellas características que están directa o indirectamente relacionadas con la domesticación. De esta manera se pierde la diversidad genética, lo cual puede ser perjudicial si en un momento dado se quiere utilizar la diversidad genética como una fuente de variabilidad para transferir a las especies en cultivo cualidades como la tolerancia a factores desfavorables, como la sequía o suelos pobres o resistencia a enfermedades. Esta variabilidad natural de los seres vivos es también uno de los recursos naturales renovables que se deben preservar para las futuras generaciones.
Las poblaciones naturales de plantas comúnmente presentan años buenos y años malos en la producción de semillas. Durante un año bueno se pueden obtener muestras grandes de semillas sanas y bien desarrolladas con alta viabilidad, y lo opuesto puede ocurrir durante un año malo, cuando sólo se encuentran semillas escasas o de baja calidad. Esto se debe, entre otras razones, a ciclos internos reproductivos presentes en una población de plantas estrechamente emparentadas, o a factores externos, como condiciones climáticas desfavorables para la reproducción o abundancia anormal de parásitos o depredadores de las partes reproductivas de las plantas. Con el propósito de evitar riesgos, la recolecta de semillas debe continuar durante varios periodos reproductivos para obtener la mejor muestra de semillas para almacenamiento o propagación.
Número de individuos de los cuales deben recolectarse las semillas
Las semillas deben recolectarse a partir del mayor número posible de plantas individuales. El mínimo de individuos aceptable para conservar algo de la variabilidad que se encuentra en la población de una localidad es de 30 o más, siempre y cuando dichos individuos sean producto de reproducción sexual y no clonal, lo cual no siempre es fácil de percibir. Un número mayor de individuos y áreas de recolección más amplias permiten una mejor representación de tal variabilidad. Las semillas de cada genotipo materno deben mantenerse separadas si se desea mejorar la variación genética durante la multiplicación.
Cuando se recogen semillas de plantas anuales o perennes de corta vida no debe recolectarse más de 20% de la cosecha local total, ya que de otra manera podría resultar afectada su posibilidad futura de sobrevivencia. Esto es particularmente importante para las especies de plantas raras, con potencial reproductivo reducido o en plantas en vías de extinción.
La calidad de una muestra de semillas frecuentemente varía ampliamente dependiendo de su origen, nivel de maduración, grado de parasitismo y depredación, limitaciones de recursos para la reproducción dentro del año de colecta y las técnicas de recolección y manejo que se hayan empleado. La calidad de las semillas recolectadas tiene gran importancia, ya que las semillas de baja calidad no resisten el almacenamiento o no germinan tan bien como aquellas colectadas con más cuidado. Debe evitarse la recolección de semillas vanas, inmaduras, parasitadas, deformes o dañadas en alguna forma (cuadro 8).
UADRO 8. Problemas principales en la colecta, el almacenamiento
y el transporte de las semillas
El momento ideal para colectar semillas es cuando los frutos sobre la planta madre están maduros y se inicia la diseminación de las semillas. Antes de ese momento las semillas pueden estar aún inmaduras fisiológica o estructuralmente e imposibilitadas para terminar su maduración. Cuando las semillas se recogen del suelo pueden estar hidratadas o contaminadas con microbios de la descomposición u hongos patógenos o parásitos, por lo que se deben recoger directamente de la planta.
Una vez colectadas las semillas debe tenerse mucho cuidado de evitar o reducir su exposición a condiciones que afecten su longevidad. Por ejemplo, se deben mantener frescas, ya que la viabilidad de las semillas y su vigor se ven reducidos conforme la temperatura se incrementa o cuando se exponen al calor más tiempo del necesario; incluso media hora de exposición al calor del Sol o al calor de un vehículo puede dañarlas. Colocar semillas húmedas en recipientes cerrados puede interrumpir la respiración normal y terminar asfixiándolas; el agua condensada del vapor de la respiración también puede promover el desarrollo de moho. La forma ideal de mantener semillas recién colectadas hasta su arribo al laboratorio es dentro de bolsas de papel o sacos de tela de algodón, porque estos materiales permiten la circulación de aire entre el interior y el exterior.
En lo que respecta a los frutos indehiscentes, la mejor manera de transportar sus semillas es dentro del fruto mismo y extraerlas en el local donde se realizará el resto de las manipulaciones.
Pruebas para calcular la viabilidad
Los laboratorios de semillas bien manejados mantienen regularmente un calendario para probar la viabilidad de las semillas almacenadas y así evitar almacenar semillas muertas, reemplazar muestras envejecidas y obtener información valiosa sobre la longevidad de las especies almacenadas. La mejor manera de averiguar su viabilidad es con una prueba de germinación, ya que otros procedimientos (cuadro 9), como la prueba del tetrazolio o el uso de respirómetros, son complicados y frecuentemente no dan resultados satisfactorios. Otra prueba, la que se realiza con rayos X, sólo es útil para verificar la cantidad de semillas dañadas o parasitadas de una muestra.
UADRO 9. Pruebas de viabilidad
Por lo anterior, es conveniente realizar una prueba de germinación de las semillas al momento en que llegan al laboratorio; en caso de que la germinación sea dificil es recomendable aplicar una serie de tratamientos para inducirla. Después de almacenar las semillas debe probarse la germinación de la muestra cada cierto tiempo y compararla con los resultados iniciales.
Tratamientos para inducir la germinación
El número de investigaciones dirigidas al estudio de los factores que inducen la germinación de las semillas de diferentes especies es inmenso y se han propuesto numerosas alternativas para romper la latencia de las semillas, abreviar su duración, e inducir la germinación de semillas afectadas fisiológicamente, envejecidas o aun inmaduras. Los cuadros que se presentan en este capítulo intentan sintetizar lo que se ha escrito al respecto. Proponemos el orden siguiente para hacer uso de esta información (cuadro 10). Si se desconoce totalmente los requerimientos de la semilla debe efectuarse una simple prueba de germinación a una temperatura media (20-27°C), favorable para la mayoría de las semillas, en un substrato húmedo apropiado (cuadro 11) y a la luz. De no germinar en esta condición entonces puede haber varios factores que estén impidiendo el proceso. En cuadro 12 se resumen las causas más comunes por las que una semilla no germina.
UADRO 10. Cómo germinar semillas

UADRO 11. Medios de germinación
UADRO 12. Causas por las que las semillas no germinan
Esta información nos dará una idea de los factores que pueden estar regulando el proceso a un nivel fisiológico, a partir de lo cual se procedería a efectuar el tratamiento más adecuado de los que se indican en los cuadros 13 al 17. Varios de estos tratamientos simulan efectos del ambiente que se dan en la naturaleza, por lo que una combinación del conocimiento de los factores que regulan la germinación de la semilla, de bases informativas sobre el ambiente en que viven las especies que se están estudiando y de una buena dosis de sentido común seguramente conducirán al éxito para lograr la germinación, a no ser que la muestra de semillas tenga embriones inmaduros, o que no sea viable a causa de los procedimientos de recolecta y de manejo previo, o por causas endógenas; en este último caso se procede a aplicar alguna de las pruebas de viabilidad que están sintetizadas en el cuadro 9.
La influencia de la temperatura sobre la germinación se conoce realizando
pruebas a diferentes temperaturas constantes, en cámaras de germinación o incubadoras
(figura 9). Sin embargo, se puede llegar a un refinamiento mayor utilizando
gradientes de temperatura, que proporcionan gran número de condiciones térmicas
constantes entre dos temperaturas límites, las cuales se establecen según las
necesidades del experimento y las condiciones del hábitat de la especie. En
la figura 10 se muestra un modelo sencillo de gradiente térmico. Sobre esta
base existen otras posibilidades de modelos más complejos, tecnológicamente
más avanzados y con mayores posibilidades de modificaciones periódicas en las
condiciones de temperatura.

Figura 9. Cámara de germinación con ambiente de luz y temperatura controlados.
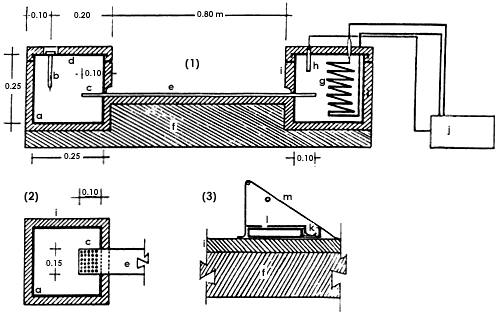
Figura 10. Gradiente térmico. En un extremo un tanque de agua fría provisto
de un serpentín enfriador (g) conectado a una unidad de refrigeración (j), controlada
por un termostato (h). En el otro extremo un tanque de agua (a) con un calefactor
con termostato (b); la barra de aluminio (c) penetra en ambos tanques y tiene
en esta zona perforaciones que mejoran su intercambio térmico con el agua. Ambos
recipientes están tapados por una cubierta del mismo material de la barra y
tanques (d). Esta barra tiene una superficie en la que tienen lugar los experimentos
(e); tanto los tanques como la barra están térmicamente aislados por una capa
de poliestireno (i) y se apoyan sobre una base de madera (f). Para los experimentos
de germinación se utiliza papel filtro (l), con un extremo sumergido en una
canaleta con agua (k). El experimento queda protegido por una cubierta plástica
sobre la barra (m), que lo cubre hasta la base de poliuretano (o)
Para conocer el efecto de las temperaturas fluctuantes en la germinación se debe fijar un termoperiodo conocido a cada una de las temperaturas que se incluyan en los tratamientos, procurando que la temperatura alta coincida con el periodo de iluminación (va a considerar la luz). Esto también puede llevarse a cabo en cámaras de germinación o en gradientes térmicos, haciendo los ajustes necesarios. Algunos de los tratamientos térmicos que se emplean para semillas de testa dura (cuadro 13) también pueden ser usados directamente o modificados, para determinar con precisión el termoperiodo y las temperaturas necesarias. La amplitud de la fluctuación de la temperatura y el termoperiodo deben establecerse tomando como base los que ocurren en el hábitat natural de la especie, particularmente en el micrositio del suelo, ya que la temperatura del suelo no sólo depende de las condiciones climáticas del lugar, sino también de la cobertura vegetal que hay sobre él, la hojarasca, la humedad, la profundidad y otras características.
UADRO 13. Pretatamientos para semillas de testa dura y/o impermeable
a los gases o al agua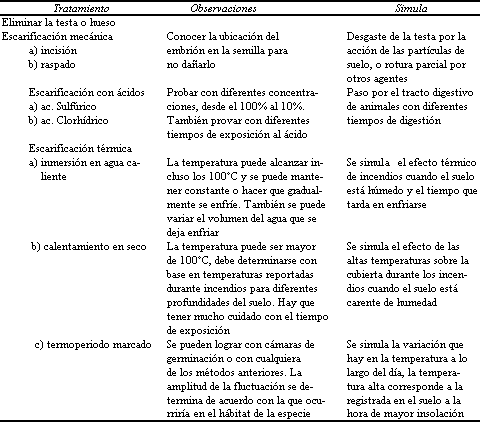
NOTA. En el caso de testa dura o impermeable asociada a la presencia
de inhibidores solubles: a) lavar las semillas con agua abundante o b)
aplicar un remojo prolongado, además de escarificar.
Con respecto al efecto de la luz también existen muchas posibilidades de experimentar. Es posible detectar si la luz afecta la germinación exclusivamente con pruebas de germinación en luz blanca y en total oscuridad. Hay que tener en cuenta que cantidades de luz mínimas, invisibles para el ojo humano, o exposiciones a la luz de fracciones de segundo, pueden afectar la germinación, por lo que no se recomienda el uso de plásticos negros u otros materiales poco confiables que dejan pasar algunas radiaciones luminosas. Un material adecuado para evitar la entrada de luz a los lotes experimentales que se desea mantener en la oscuridad es el papel aluminio comercial, del cual se recomienda poner una capa doble.
Incluso en el experimento más sencillo de luz debe determinarse un fotoperiodo, es decir, el tiempo que estarán las semillas a la luz y a la oscuridad durante el día, ya que fotoperiodos de 24 horas de luz pueden dar resultados diferentes de los que se obtienen con fotoperiodos de 12 horas o menores.
Experimentos más complejos para probar el efecto de diferentes longitudes de onda y su combinación en la germinación están resumidos en el cuadro 14. Los materiales con los que se puede crear, en forma económica pero segura, filtros de luz de longitud de onda conocida se presentan en el cuadro 15. Estos filtros son de banda ancha pues incluyen una banda de luz de aproximadamente 30 nanómetros, centrada en la longitud de onda de la radiación que interviene en los procesos fotoblásticos. En muchos países estos filtros son bien aceptados para el trabajo experimental. Los filtros de banda estrecha (de 20, 10 o menos nanómetros) son por lo general cristales de pocos centímetros de superficie y sumamente caros. Es muy importante señalar que la combinación de filtros y fuentes de luz debe ser la correcta para crear las condiciones lumínicas precisas para estudiar el fotoblastismo. En el cuadro 15 se señalan estas combinaciones. Si no se toman en cuenta es muy probable que se llegue a conclusiones erróneas.
CUADRO 14. Pruebas básicas para conocer la respuesta de las semillas a la luz. Con asterisco se marcan las pruebas que son opcionales. A = luz azul, LB = luz blanca, R = luz roja, RL= luz roja lejana, AG= adicionar al medio de germinación ácido giberélico y BFF= probar con flujos fotónicos más bajos de la luz indicada. Dependiendo del diseño experimental se pueden repetir las pruebas en semillas de más edad.
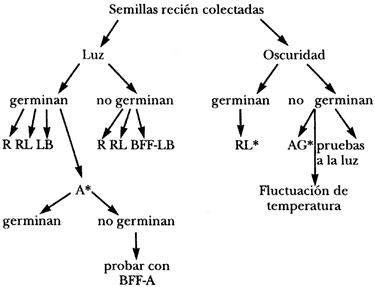
CUADRO 15. Materiales de bajo costo con los que se pueden generar
en forma sencilla ambientes de luz con calidad espectral conocida

Tal como vimos anteriormente, bajo la cobertura vegetal la calidad de luz no
es homogénea y también hay diferencias en la calidad de la luz filtrada por
distintas composiciones del follaje; esto afecta la germinación de las semillas
fotosensibles, debido a los diferentes valores de la relación rojo-rojo lejano
que se establecen en cada condición bajo el dosel. Diferentes valores de la
relación rojo:rojo lejano de la luz pueden simularse con el gradiente de luz
que se muestra en la figura 11.
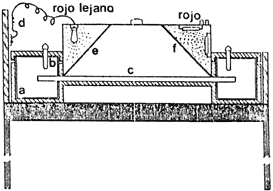
Figura 11. Gradiente de luz. El mismo dispositivo descrito en la figura
10, pero en este caso carece de una unidad de refrigeración para que la barra
se mantenga a temperatura constante. Esto se logra gracias a que: a) en ambos
tanques: existen calefactores, y b) que en caso necesario permitirían elevar
la temperatura de la barra por arriba de la del ambiente. Se trata de una caja
de aluminio rectangular. En un extremo tiene una luz incandescente con un control
de intensidad o dimer (d) con un filtro de color rojo y otro azul (e)
que permite el paso del rojo lejano. En el otro extremo dos lámparas fluorescentes
pequeñas y un filtro rojo f) que transmite luz roja. Sobre la superficie de
la barra c) se forma un gradiente de luz roja/rojo lejano de un extremo a otro.
Es posible determinar los termoperiodos y los fotoperiodos con base en lo que ocurre en condiciones naturales, aunque los que son cortos para una especie pueden resultar largos para otra. Los fisiólogos muchas veces emplean fotoperiodos y termoperiodos que, en términos generales, no corresponden con lo que ocurre en la naturaleza, pero que proporcionan información fisiológica muy precisa de los procesos que regulan la respuesta a la luz, y muchas veces tienen relación con el microambiente que experimenta la semilla en una determinada condición. Por ejemplo, una breve exposición a la luz cuando la semilla es desenterrada por el paso de un tractor, para quedar nuevamente enterrada al terminar de removerse el suelo. En la naturaleza es muy variable la cantidad de luz que llega a las semillas, dependiendo de si están enterradas o no, de la profundidad de enterramiento, de si existe o no una cobertura vegetal, de la presencia de hojarasca, incluso de las características de la cubierta de la semilla. Estas diferencias en el flujo fotónico (cantidad de fotones que llega a un lugar) también afectan la respuesta germinativa, por lo que, en la medida posible, es conveniente saber la cantidad de luz que estamos empleando en condiciones experimentales.
Algunas semillas que presentan latencias de cualquier naturaleza inician su germinación en presencia de sustancias reguladoras del crecimiento vegetal, algunas de ellas son hormonas vegetales. Estas sustancias desencadenan procesos fisiológicos relacionados con la germinación y sustituyen algún factor que actúa en el medio natural, como la luz o la temperatura (bajas o fluctuantes). Lo mismo puede ocurrir si se emplean sales minerales, compuestos nitrogenados y soluciones que modifiquen la permeabilidad de la membrana de las células de la semilla (cuadro 16).
UADRO 16. Estímulos químicos para la germinación
Existen especies que producen semillas cuya germinación es estimulada por los tratamientos físicos y químicos que sufren al transitar por el tubo digestivo de animales dispersores de semillas, que se alimentan de frutos o follaje, como las aves, los murciélagos frugívoros, los monos y muchos otros mamíferos. Existen tratamientos que intentan simular el efecto de ese tránsito. También hay plantas que requieren un medio externo muy particular o la asociación con otros organismos para germinar: en el cuadro 17 se resumen algunas de estas posibilidades.
UADRO 17. Semillas que reciben el estímulo germinativo a través
de interacciones bióticas.
La creatividad y el conocimiento previo de la ecología de las especies pueden dar lugar al diseño de innumerables variantes y combinaciones, con mezclas de experimentos de campo y de laboratorio. Esta riqueza de posibilidades hace de las semillas un material ideal para el diseño de experimentos de investigación científica, ya que su respuesta a un estímulo adecuado o inadecuado para inducir el proceso fisiológico de la germinación se detecta con relativa facilidad.
La respuesta germinativa, en cualquier condición experimental, se puede evaluar por el número de semillas germinadas (capacidad germinativa), el cual se expresa generalmente como porcentaje, o por la velocidad de germinación (número de semillas germinadas por unidad de tiempo). Estas formas de evaluación nos proporcionan información diferente, ya que muchas veces una especie tiene alta capacidad germinativa en una condición ambiental particular, pero el tiempo necesario para lograrla es muy largo, o bien puede ocurrir lo contrario. Estas dos posibilidades implican diferencias en la oportunidad de colonizar o competir por los recursos (espacio, luz, etc.) con otras especies.
El almacenamiento de semillas es una manera importante de mantener vivo, por largos periodos de tiempo, el germoplasma de plantas superiores. Sin embargo, los almacenes de semillas, particularmente en los trópicos, con frecuencia carecen de condiciones adecuadas para prolongar al máximo la duración de la viabilidad de las semillas, ya que se requieren técnicas de manejo y almacenamiento apropiadas y equipos relativamente costosos para lograr ese propósito.
No todas las semillas son almacenadas con éxito. Algunas permanecen vivas por tiempo indefinido, quizá varios siglos, cuando se almacenan en el ambiente más apropiado para interrumpir su deterioro natural, en tanto que otras sufren un deterioro rápido por causas endógenas, ajenas a las condiciones externas. Estas diferencias se deben al hecho de que las semillas son liberadas al medio con diferentes niveles de humedad, composición química y tasa metabólica, lo cual afecta su longevidad potencial.
Existen diferentes clasificaciones de las semillas, según la duración potencial de su viabilidad (cuadro 18). De acuerdo con el laboratorio de semillas de la Universidad de Reading, Reino Unido, las semillas se clasifican en tres categorías: ortodoxas, recalcitrantes e intermedias. Para llegar a esta definición se basaron en los siguientes criterios.
CUADRO 18. Clasificación de las semillas, con base en la duración
de su viabilidad, de acuerdo con diferentes autores

Estas semillas pueden ser desecadas hasta contenidos de humedad muy bajos sin sufrir daños, al menos hasta un nivel de humedad constante que se mantenga en equilibrio con una humedad ambiental relativa de 10% (a este valor las semillas con almidón tienen un contenido de humedad cercano a 5%, en tanto que las semillas grasas tienen valores de 2 a 3%).
Sus longevidades aumentan cuando disminuye el contenido de humedad y con la temperatura durante el almacenamiento, en una forma cuantificable y predecible.
En contraste con las semillas ortodoxas, las recalcitrantes no pueden ser desecadas por debajo de un punto relativamente alto en el contenido de humedad sin causarles daño. A pesar de que existe gran variación en el contenido de humedad crítico entre las especies, bajo el cual la viabilidad se reduce, algunas especies comienzan a morir rápidamente aun en equilibrio con una humedad relativa ambiental de 98-99%, y la mayoría de las semillas muere cuando su contenido de humedad está en equilibrio con una humedad ambiental de 60-70% (que corresponde a un contenido de humedad de 16-30% sobre el peso fresco). Todavía no existe un método satisfactorio para mantener la viabilidad de las semillas de estas especies, en particular las de origen tropical, por arriba de un periodo corto, menor a un año.
Una tercera categoría de comportamiento de las semillas en almacenamiento ha sido demostrada recientemente con semillas de café, palma aceitera, papaya y neem. La principal característica de este comportamiento es cierta sensibilidad a la desecación hasta un nivel de humedad relativamente bajo de 7 a 10% (en equilibrio con una humedad relativa ambiental de 30-50%). Sin embargo, la longevidad de las semillas secas de origen tropical se reduce en temperaturas bajas (por debajo de 5°C) y temperaturas bajo cero. Por esto, las condiciones ideales para el almacenamiento a largo plazo de semillas ortodoxas (5% de contenido de humedad, -18°C) son potencialmente dañinas para las semillas intermedias y no deben usarse ya que les provoca la muerte en pocos meses. A pesar de esto, es posible almacenar las semillas "intermedias" por periodos de alrededor de 10 años, desecándolas hasta un 7-10% de contenido de humedad, y manteniéndolas a la temperatura de un laboratorio.
Almacenamiento de semillas en el trópico
Muchas especies de árboles tropicales producen semillas con altos contenidos de humedad y rápidas tasas metabólicas, que se comportan como recalcitrantes cuando se pretende almacenarlas. Esto se debe a que carecen de la posibilidad de rearreglar la estructura de sus componentes celulares durante la salida del agua de las células, ocasionada por la deshidratación, por lo que se pierde la estructura funcional del protoplasma que ya no puede recobrarse al ocurrir la rehidratación. La presencia de agua libre en las células elimina el efecto protector de la congelación debido a la formación de cristales de hielo que pueden dañar a las células; sin embargo, estudios recientes muestran que algunas semillas que se supone eran recalcitrantes, son deshidratadas cuidadosamente a niveles que hacen posible su almacenamiento prolongado, como es el caso de la papaya. Es probable entonces que en el futuro, gracias a investigaciones más extensivas, se encuentre mayor número de semillas de comportamiento intermedio, indicando que lo ortodoxo y lo recalcitrante son los extremos de un gradiente de posibilidades.
Hasta ahora hay poca esperanza de que se encuentren tratamientos que prolonguen significativamente la longevidad en almacenamiento de las semillas más recalcitrantes, sobre todo si se toma en cuenta las escasas investigaciones científicas al respecto. Algunas alternativas para almacenar semillas recalcitrantes se presentan en el cuadro 19. Los laboratorios que continúan trabajando en esta dirección se encuentran en Inglaterra, Francia, Estados Unidos y Sudáfrica. Entre los más importantes están el Almacén Nacional de Semillas del Departamento de Agricultura de Estados Unidos en la Universidad Estatal de Colorado en Fort Collins, CO; el jardín Botánico de Kew en Sussex, Reino Unido; el Centro Nacional de Investigación Científica de Montpellier, Francia, y la Universidad de Natal en Sudáfrica.
UADRO 19. Alternativas para almacenar semillas de corta longevidad
Debido a la falta de técnicas apropiadas para el manejo de semillas recalcitrantes, producidas por muchas de las especies de árboles tropicales valiosos, y también a que en la mayoría de los países tropicales no existen sistemas eficientes de recolecta y distribución de semillas, la mayoría de los proveedores de semillas en el trópico ofrecen a sus clientes casi exclusivamente especies exóticas, como los eucaliptos, los pinos y sólo algunas especies tropicales con semillas típicamente ortodoxas. La mayoría de las especies forestales tropicales con madera, frutas o propiedades medicinales valiosas no están disponibles en ningún banco de semillas, ni comerciales ni institucionales.
Pruebas sencillas para definir si una semilla es ortodoxa o recalcitrante
A pesar de que es posible hacer presunciones acerca del comportamiento de una especie de semilla en almacenamiento basándose en su tamaño, apariencia, historia de vida y filogenia, es necesario hacer pruebas para saber con precisión el comportamiento de cada especie en particular. La prueba se inicia dividiendo una porción de semillas en dos partes iguales. Se prueba la viabilidad de una de las fracciones de semillas frescas y la otra mitad se somete a una desecación gradual y cuidadosa antes de probar su viabilidad. Para completar, una prueba adicional que se realiza antes y después de someter las semillas a congelación, indicará si las semillas que toleran la desecación son ortodoxas verdaderas o intermedias (cuadro 20).
UADRO 20. Pruebas para determinar semillas ortodoxas y recalcitrantes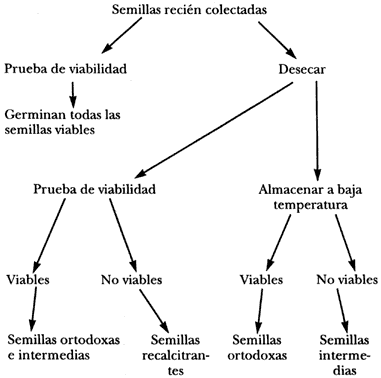
Almacenamiento de semillas ortodoxas
Las semillas ortodoxas se almacenan mejor cuando su contenido de humedad disminuye. El contenido de humedad óptimo puede ser ligeramente diferente entre especies, por lo que una manera sencilla de reducirlo, aproximadamente hasta el nivel óptimo para la mayoría de las especies, consiste en colocar éstas a una temperatura ambiente de alrededor de 15 a 19°C, expuestas a una atmósfera con humedad relativa de alrededor de 18 o 20% durante un mes. Esto se logra usando una cámara de deshidratación fresca o desecadores de campana de vidrio provistos de sílica-gel deshidratada o alúmina activada. Las campanas deben ser colocadas en un cuarto fresco.
Una instalación dedicada al almacenamiento de semillas debe estar provista de un cuarto de secado, con aire acondicionado y deshumidificadores, para destinarlo al proceso continuo de deshidratación de muestras de semillas. Debe recordarse que las semillas nunca deben secarse con calor, pues pocas sobreviven a estas condiciones.
Muchas semillas resisten una deshidratación muy profunda; en esos casos, el almacenamiento se realiza en tubos de cristal que contengan una sustancia deshidratante como el sílica-gel. También el hidróxido de sodio es un deshidratante muy utilizado para otros propósitos, pero nunca debe usarse con semillas.
Cuando las semillas están secas deben colocarse en contenedores o recipientes herméticamente cerrados y etiquetados, precisando el nombre de la planta, la fecha, el lugar de recolección y otros datos importantes. Los bancos de semillas emplean diferentes clases de recipientes, como frascos herméticos de cristal o de metal y tubos de cristal cerrados a fuego. Los bancos de semillas importantes utilizan preferentemente bolsas gruesas de polietileno selladas a fuego dentro de sobres de papel de aluminio.
Para instalaciones relativamente pequeñas de almacenamiento de semillas, son muy convenientes los viales de vidrio cerrados a fuego que contengan a las semillas, separadas de la sustancia deshidratante por medio de un algodón (figura 12), ya que con ello se evita la humidificación accidental de las muestras. La muestra inicial de semillas se distribuye en varios viales, de manera que se cuente con material para almacenamiento prolongado, para almacenamiento temporal y para intercambio, sin riesgo de que al abrir y cerrar los recipientes se humedezcan las semillas accidentalmente (cuadro 21).
Figura 12. Ámpula de vidrio sellada a fuego para almacenar semillas desecadas
a baja temperatura.
UADRO 21. Método para almacenar semillas ortodoxas en ámpulas de vidrio
Las cámaras refrigeradas para el almacenamiento de semillas pueden variar mucho en tamaño y complejidad, dependiendo de las necesidades de cada instalación de almacenamiento. Las cámaras subterráneas amplias, construidas de paredes gruesas de concreto, son ideales para una instalación de gran tamaño. Están aisladas de los cambios atmosféricos de temperatura, requieren menos energía para funcionar y, sobre todo, pueden resistir catástrofes ambientales o guerras. El almacén de semillas de pino del vivero del bosque Pine Ridge en Smoky Lake, Alberta, Canadá, es un edificio con paredes reforzadas de concreto de 43 cm de grosor, enterrado bajo una capa de 2 m de suelo, con excepción de la rampa de entrada y de un cuarto de servicios mecánicos. Entre las ventajas de esta instalación está: una buena protección del fuego; un ahorro considerable en consumo de energía, debido a la mínima exposición a las temperaturas de la superficie, y el hecho de que las fallas eléctricas y mecánicas tienen un efecto menor sobre las condiciones de almacenamiento, debido al excelente aislamiento del edificio. El cuarto de almacenamiento se mantiene a -18°C y menos de 50% de humedad relativa. El cuarto subterráneo de manejo de semillas se mantiene a 4°C para evitar la condensación de agua sobre las semillas frías, cuando los contenedores de almacenamiento tienen que ser abiertos. Instalaciones similares deberían construirse en varios lugares y alguna organización internacional debería encargarse de la construcción de un almacén de semillas, de gran capacidad, que estuviera situado en una instalación subterránea construida en algún lugar como la Antártida, el permafrost ártico o las nieves perpetuas de una zona montañosa. Esto permitiría que el valioso germoplasma de las semillas de plantas de cultivo o silvestres sobreviviera periodos caóticos de guerra, trastornos políticos, caos económicos, o catástrofes ecológicas.
Varias obras describen cómo construir una cámara de almacenamiento de semillas y se citan en la bibliografía. Cualquier laboratorio, herbario, jardín botánico o vivero puede mantener un banco de semillas ortodoxas operando con pocos recursos y personal tomando en cuenta las siguientes indicaciones:
Las semillas ortodoxas se pueden almacenar en un congelador doméstico de tipo horizontal o vertical, pero deben evitarse los cambios prolongados de temperatura durante fallas eléctricas mediante el empleo de un generador que se encienda automáticamente al ocurrir la falla eléctrica. El procedimiento de almacenar las semillas en viales cerrados a fuego es el más adecuado cuando no se dispone de un cuarto especial refrigerado para abrir los recipientes, ya que es posible dejar que los viales gradualmente tomen la temperatura ambiente antes de proceder a abrirlos, y sólo una pequeña muestra de semillas es expuesta cuando se requiere extraer algunas semillas del banco. En cualquier caso, ya se trate de un almacén de semillas pequeño o de un gran proyecto de importancia nacional, es indispensable una política estable de conservación del germoplasma, un administrador y personal bien preparado e interesado en el proyecto, así como un apoyo económico continuo para asegurar la perpetuación de la instalación y su buen funcionamiento.
Cómo extraer las semillas de la cámara fría
La forma ideal de transportar o intercambiar semillas almacenadas es mantenerlas dentro de los recipientes sellados que se utilizan en el almacén, pues de otra manera hay riesgo de una rápida rehidratación dañina. Así que lo mejor es distribuir una muestra de semillas grande en muchos recipientes pequeños para que no haya necesidad de abrir los recipientes para manejar las semillas.
Antes de plantar las semillas almacenadas éstas deben ser rehidratadas cuidadosamente. Primero se ponen, por algún tiempo, en contacto con una atmósfera fresca y ligeramente húmeda, antes de ponerlas en contacto con el agua para que comiencen a recuperar la humedad lentamente.
Para establer un centro de manejo y conservación de semillas se debe tener información preliminar acerca de las especies prioritarias en cuanto al manejo de semillas, la posibilidad de que exista una demanda de estas especies, la disponibilidad de las semillas y la capacidad de manejo en cuanto al número y variedad de especies que las instalaciones permiten.
Esta institución realizaría tres funciones básicas: conservación y distribución de semillas con fines de propagación, investigación básica y aplicada, y labores de extensión y entrenamiento. Las semillas se obtendrían en viajes de colecta al campo y por medio de compra o intercambio con otros laboratorios y jardines botánicos. Por esto sería importante que el organismo descrito tuviera un control adecuado de las fuentes de semillas para asegurar su estabilidad y disponibilidad, ya que de otra manera la existencia continua de semillas de alguna especie podría verse interrumpida al incrementarse la demanda. Además de la recolecta, las labores cotidianas del centro serían: 1) aplicar pruebas a las semillas, cada muestra de semillas que ingrese al centro deberá ser analizada para conocer su pureza, sanidad y poder germinativo, con el fin de conservar e intercambiar los mejores lotes de semillas; 2) documentación de las semillas, para lo cual se llevaría un registro completo de la edad, el origen y las características de cada muestra mediante el registro de datos esenciales como el lugar y la fecha de colecta, la viabilidad, los tratamientos aplicados y cualquier otro dato relevante. Estos dos procedimientos permiten obtener semillas certificadas, en las que su calidad y sus características están bien documentadas y pueden ser respaldadas por medio de un certificado.
Otra función del centro, de no menor importancia, sería la investigación básica de las semillas y de las especies que se manejan, ya que con frecuencia en el mundo tropical se carece del más mínimo conocimiento de las especies disponibles y esto ha sido un obstáculo serio para la utilización de muchas especies de árboles tropicales. Mediante la investigación es posible determinar los métodos más adecuados para colectar, procesar, almacenar y germinar las semillas de las diversas especies, además de que también se obtiene información básica sobre la fisiología de las semillas y sobre los mecanismos de germinación y establecimiento de plantas.
Un centro de tales características sería un modelo para futuros establecimientos similares en los trópicos; además, tendría una función importante en el entrenamiento de personal en las diferentes técnicas y en el desarrollo de nuevos procedimientos
PROBLEMAS EN EL USO DE SEMILLAS EN LA PROPAGACIÓN DE LAS PLANTAS
La mayoría de los árboles más valiosos de los bosques templados se propaga por medio de semillas, en tanto que los árboles frutales y de ornato con frecuencia son propagados utilizando segmentos vegetativos o injertos. Más recientemente se ha tenido éxito con técnicas de micropropagación en algunas especies.
Con respecto a los árboles tropicales, la mayoría de las especies que se cultivan extensivamente se propaga por medio de semillas; sin embargo, no todas las especies valiosas producen semillas que puedan utilizarse fácilmente para este propósito. Muchos árboles tropicales florecen y fructifican esporádicamente o producen cantidades pequeñas de semillas. Con frecuencia éstas son fuertemente dañadas por parásitos y depredadores. Muchas de ellas son de tipo recalcitrante, de vida corta, frágiles y difíciles de manejar o almacenar. Algunas requieren condiciones peculiares de germinación que son desconocidas o difíciles de obtener en un vivero o en una cámara de germinación; además, las plántulas que se obtienen de las semillas de los árboles tropicales a veces necesitan mucho tiempo para crecer y su maduración puede tomar toda una vida.
Un programa sobre conservación y manejo de semillas tiene objetivos a corto y a largo plazo. Los de corto plazo se concentran en el desarrollo de una estrategia para la recolecta representativa de semillas y para cubrir la demanda de éstas. Los objetivos a largo plazo son asegurar la existencia de una provisión sostenida de semillas de alta calidad y de larga viabilidad, y desarrollar plantas de mayor calidad mediante programas de mejoramiento y selección de las características genéticas más deseables de estas plantas. De esta manera, un programa bien equilibrado de manejo de semillas tiene esencialmente tres componentes: 1) la obtención de las semillas, que es el conjunto de prácticas organizadas para obtener y conservar semillas de alta calidad y mantener una disponibilidad permanente de éstas; 2) el mejoramiento de árboles, que es el conjunto de prácticas diseñadas para obtener genotipos más deseables de plantas para la reproducción y propagación, y 3) la conservación de recursos genéticos, que es el conjunto de prácticas diseñadas para conservar el germoplasma a largo plazo y con cierto grado de variabilidad genética para las demandas futuras.
En párrafos anteriores hemos tratado ampliamente los aspectos de manejo y conservación de semillas, por lo que sólo resta tratar aquí el tema del mejoramiento genético de una manera somera, ya que se trata de un asunto de suma complejidad.
Obtener muestras de semillas mejoradas es una necesidad que puede resolverse de diferentes maneras. En especies de vida larga, en que se requieren muchos años para que las plantas alcancen la edad reproductiva, las prácticas de entrecruza, hibridización y selección que se utilizan en los cultivos de plantas anuales son difíciles de aplicar y sólo se obtienen resultados a muy largo plazo. Por esta razón, en plantas leñosas se recurre frecuentemente a la selección clonal o vegetativa, de la que se habla más extensamente en el siguiente capítulo.
Una de las técnicas más empleadas para asegurar una fuente constante y de buena calidad de semillas, sobre todo en el caso de árboles de valor forestal, es la creación de rodales semilleros. Esto se lleva a cabo seleccionando árboles de alta calidad, buena forma y alta producción de semillas viables. Con estas semillas se forman cultivos en áreas de suelos y humedad adecuada y los individuos que aquí nacen constituyen una fuente valiosa de semillas, que son las que se utilizan para generar los viveros destinados a reforestación y reemplazo de los árboles cosechados en los bosques.
Sin embargo, no debe pensarse que las técnicas de selección sexual no se han aplicado a plantas longevas. Las culturas con larga tradición hortícola nos han legado árboles cultivados con propiedades alimentarias, o de otra índole, muy valiosas, que indudablemente fueron el resultado de técnicas de hibridización o entrecruza en las que se hizo gala de paciencia, una de las cualidades más escasas de la modernidad.
![[Inicio]](../img/begin.gif)
![[Anterior]](../img/prevsec.gif)

![[Siguiente]](../img/nextsec.gif)