II. LA PROPAGACIÓN VEGETATIVA
A DIFERENCIA de lo que sucede en los vertebrados, la semilla es liberada cuando aún sus tejidos no se han diferenciado totalmente (véase Cómo viven las plantas, La Ciencia desde México, núm. 49). La semilla contiene células embrionarias que darán origen a todos los tejidos de la nueva planta después del proceso de germinación; además, muchas de las células de los tejidos vegetales ya maduros conservan la potencialidad de diferenciarse y dar origen a diversas estructuras; estas células forman parte de meristemos primarios y secundarios que pueden encontrarse en todos los órganos de las plantas. Gracias a esto es posible obtener plantas enteras a partir de tejidos de yemas, tallos, raíces y hasta hojas de casi cualquier planta.
La propagación clonal o vegetativa de plantas es una producción a partir de partes vegetativas. Se utilizan tejidos vegetales que conserven la potencialidad de multiplicación y diferenciación celular para generar nuevos tallos y raíces a partir de cúmulos celulares presentes en diversos órganos. Este tipo de propagación tiene esencialmente tres variantes, que son: 1) la micropropagación a partir de tejidos vegetales en cultivo in vitro; 2) la propagación a partir de bulbos, rizomas, estolones, tubérculos o segmentos (esquejes) de las plantas que conserven la potencialidad de enraizar, y 3) la propagación por injertos de segmentos de la planta sobre tallos de plantas receptivas más resistentes.
La propagación vegetativa comprende desde procedimientos sencillos, conocidos de tiempos inmemoriales por los campesinos de todo el mundo, hasta procedimientos tecnológicamente muy avanzados, basados en la tecnología del cultivo de tejidos vegetales, mediante los cuales se puede lograr la propagación masiva de plantas genéticamente homogéneas, mejoradas y libres de parásitos. Los procedimientos modernos permiten la obtención de cultivares totalmente libres de agentes patógenos, incluyendo virus, e incluso la fabricación de semillas artificiales por medio de la técnica de embriogénesis somática y encapsulado. Además de la propagación, las técnicas de cultivo de tejidos in vitro también permiten seguir procedimientos modernos de conservación de germoplasma gracias al mantenimiento prolongado de cultivos de crecimiento lento y la criopreservación de tejidos.
ESTRUCTURAS DE PROPAGACIÓN VEGETATIVA
Varias especies de plantas vasculares, en su mayoría especies cultivadas, no producen semillas aunque tengan flores, su multiplicación o propagación vegetativa no implica la fusión de células germinativas. Esta forma de propagación también se presenta en plantas que normalmente producen semillas, y sólo se le considera como reproducción asexual cuando sustituye en gran parte a la reproducción sexual.
Se trata de un proceso que implica el enraizamiento y la separación de una parte de la. planta original cuando mueren los tejidos vegetales que las semillas unían. De esta manera, las células, tejidos u órganos desprendidos se desarrollan directamente en nuevos individuos. Las zonas de abscisión pueden ser precisas, como sucede en la separación de los bulbilos, o puede darse la fragmentación de una planta debida al deterioro y muerte del individuo parental o bien de los tejidos de interconexión, como en el caso de los brotes de las raíces.
Las estructuras de propagación vegetativa funcionan también como órganos de resistencia y de almacenamiento en las temporadas adversas, los cuales algunas veces son almacenados por tiempos prolongados.
Estructuras de propagación vegetativa en plantas no vasculares
La propagación vegetativa se presenta en todo el reino vegetal; por ejemplo, en algunas algas pluricelulares la propagación vegetativa se realiza mediante su fragmentación en dos o más individuos. Las cianobacterias presentan a lo largo de sus filamentos unas células muertas, agrandadas y de pared gruesa, que se encuentran a intervalos a lo largo de sus filamentos, las cuales ayudan a la fragmentación.
Varios tipos de plantas no vasculares tienen estructuras especializadas relacionadas con la propagación vegetativa. Las hepáticas producen estructuras semejantes a las yemas llamadas propágulos, que al desprenderse de su pedicelo son arrastrados por la lluvia hasta sitios en los que se desarrollan como nuevas plantas, mientras que los líquenes producen cuerpos reproductores conocidos como soredios, integrados por masas de hifas fúngicas y de células algales.
Estructuras de propagación vegetativa en plantas vasculares
En virtud de la totipotencialidad del tejido vegetal, es decir, de su capacidad para formar yemas y raíces adventicias, casi cualquiera de los órganos de una planta vascular tiene relación con su propagación vegetativa al sufrir modificaciones anatómicas y funcionales que le permiten desarrollarse en un organismo vegetal completo e independiente, con las mismas características genéticas de la planta progenitora. Las yemas, por lo general, se encuentran en las axilas de las hojas, en la porción terminal del tallo, o bien se desarrollan en cualquier porción del tallo y dan origen a raíces adventicias.
Entre las estructuras de propagación vegetativa algunas comparten semejanzas en su desarrollo, por lo que no siempre es posible hacer una diferenciación muy clara entre ellas, sino que más bien se ubican en un continuo de características. Sin embargo, algunos autores las clasifican tomando en cuenta los órganos vegetales de los cuales se originan. Con base en este criterio describiremos algunas de las más comunes.
Propagación vegetativa por tallos y yemas
Los tallos horizontales aéreos y subterráneos de varias especies silvestres y cultivadas se alargan y forman raíces adventicias en sus nudos. Mientras los tejidos se mantienen intactos se trata del crecimiento de una sola planta, como sucede en muchas especies de gramíneas. A este individuo completo de extenso crecimiento se le conoce como genet o clon. Pero cuando el tejido de interconexión muere o es cortado, cada uno de los segmentos da lugar a un nuevo individuo al que se le conoce como ramet (figura 13A).
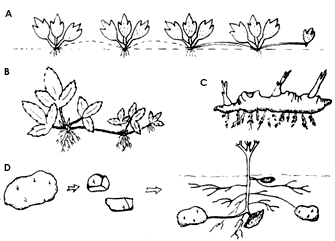
Figura 13. A) Separación de los ramets individuales por muerte del tejido
de interconexión; B) estolones; C) rizoma; D) el tubérculo (papa) se corta en
piezas y cada una contiene una yema para que a partir de los tubérculos se propagen
más plantas.
Una modificación de este tipo de propagación ocurre cuando el extremo libre de un tallo largo alcanza el suelo y además de desarrollar raíces adventicias, la yema de crecimiento da lugar a un tallo erecto, lo que se conoce como acodadura (figura 14); este proceso ocurre frecuentemente en la propagación de las frambuesas y las zarzamoras.

Figura 14. Acodadura. Las ramas alcanzan el suelo y enraizan.
Por otro lado, los tallos aéreos de algunas hierbas y arbustos caen por su propio peso al suelo. La producción de raíces adventicias y la muerte de las conexiones con el individuo parental permiten la generación de plantas independientes.
En otros casos, la sola fragmentación de los tallos o de las ramas y su contacto continuo con el suelo es suficiente para que los segmentos formen raíces y se desarrolle un individuo completo. Este tipo de propagación es común en los cactos, los sauces y en la planta acuática conocida como elodea.
Entre las principales estructuras de propagación vegetativa originadas a partir de los tallos y de las yemas se encuentran las siguientes:
Propagación vegetativa por tallos
1) Estolones. Constan de secciones relativamente largas y delgadas de tallos aéreos horizontales con entrenudos largos y cortos alternados que generan raíces adventicias. La separación de estos segmentos enraizados permite el desarrollo de plantas hijas. La fresa es un ejemplo de las especies que comúnmente presentan este tipo de propagación (figura 13B).
2) Rizomas. Se generan a partir del crecimiento horizontal de un tallo subterráneo, por lo general más robusto que el que da origen a un estolón. Las viejas porciones se degradan y se separan en fragmentos que deberán enraizar de manera independiente. Este tallo subterráneo presenta hojas escamosas en las axilas, donde se pueden generar yemas axilares, además de presentar raíces adventicias (figura 13C). Una vez formado el vástago principal se da un crecimiento continuo. Cada estación de crecimiento presenta un crecimiento simpodial por medio de la yema axilar o monopodial por medio de la yema terminal. El rizoma funciona como órgano de almacenamiento de reservas. De esta manera se propagan especies de importancia económica, tales como el bambú, la caña de azúcar, el plátano, así como algunos pastos.
3) Tubérculos. Son estructuras gruesas, suculentas, que actúan también como estructuras de reserva. Se forman en el extremo de tallos subterráneos delgados. Un ejemplo muy conocido lo constituye la papa. Los tubérculos presentan en su superficie nudos con hojas escamosas, arreglados de manera espiral, y cada uno de ellos consta de una o más yemas pequeñas. Cuando se inicia el crecimiento del vástago principal las raíces adventicias se desarrollan en la base del tubérculo y las yemas horizontales se alargan y producen tallos etiolados en forma de estolones. A partir de los tubérculos que han formado ramas horizontales se forman tubérculos nuevos (figura 13D).
Los tubérculos y los rizomas son muy semejantes y en algunos casos es casi imposible distinguirlos. Sin embargo, una característica distintiva de un rizoma verdadero es que presenta un grosor uniforme en toda su longitud, sobre la cual crecen raíces adventicias, las cuales no existen en los nudos de los tubérculos. Otra diferencia entre estas estructuras consiste en que el rizoma formará el vástago principal de la nueva planta, mientras que el tubérculo forma ramas laterales (figura 13C y 13D).
4) Brotes. Se definen como ramas o tallos que desarrollan raíces adventicias sin que sean independientes de la planta progenitora. Se desarrollan en las axilas de las hojas escamosas o de las yemas adventicias sobre las raíces. En la piña comestible los brotes se desarrollan en las axilas de las hojas inferiores que son cubiertas por el suelo.
Propagación vegetativa por yemas.
A partir de la producción de las yemas axilares con orientación vertical en
los tallos de algunas plantas (figura 15A) y de su posterior desprendimiento
y caída al suelo, se producen estructuras de propagación vegetativa tales como
los bulbos que se presentan en la cebolla, el tulipán y el lirio o los cormos
del gladiolo y el azafrán. Ambas estructuras, una vez liberadas, se establecen
de manera subterránea pero forman ramas que dan lugar a nuevas plantas.
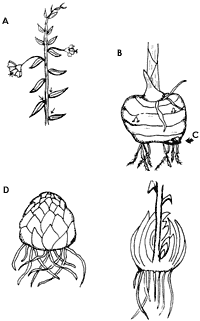
Figura 15. A) Yemas axilares; B) cormo; C) cormelo; D) bulbo.
1) Cormos. Se forman en las yemas de las axilas de las hojas de un tallo robusto y suculento que proporciona los nutrientes necesarios para la nueva estructura, la cual se desprenderá del progenitor y se desarrollará subterráneamente como un tallo corto, erecto y sólido con nudos y entrenudos. Los cormos tienen forma de esferas aplanadas dorsoventralmente, como los del gladiolo y el azafrán. Están envueltos en delgadas hojas escamosas que los protegen del daño físico y de la pérdida de agua, pero que no funcionan como estructuras de almacenamiento, a diferencia de las escamas de los bulbos. Cuando se desprenden las escamas marcan círculos alrededor del cormo. Éste desarrolla raíces adventicias ventrales o basales. El ápice del cormo es un vástago terminal que se desarrollará en las hojas y en un vástago floral terminado por una inflorescencia, y en cada uno de los nudos se producen las yemas axilares (figura 15B). El cormo se multiplica ramificándose simpódicamente, y si se corta un cormo, manteniendo una yema en cada sección, cada uno de estos segmentos desarrollará un cormo nuevo.
2) Cormelos. Sobre el extremo inferior del cormo se producen pequeñas estructuras semejantes a los estolones conocidos como cormelos (figura 15C). La muerte del cormo parental permitirá la separación de los cormos hijos, los cuales pueden ser almacenados durante el invierno y plantados durante la temporada favorable para el crecimiento.
3) Bulbos. Se desarrollan sobre tallos cortos y engrosados, a partir de yemas axilares de hojas carnosas. De éstas obtienen elementos de reserva, a diferencia de los cormos que las obtienen a partir del tallo, lo cual les permite producir rápidamente raíces adventicias. Se desarrollan subterráneamente en forma de tallos carnosos, cubiertos con hojas engrosadas a manera de escamas que funcionan como órganos de reserva (figura 15D).
Es posible que se produzca más de un bulbo a partir de cada yema. En algunos casos se desarrollan masas de bulbos en el extremo del tallo, cada uno de ellos llamados bulbilos, los cuales pueden ser dispersados lejos del bulbo parental. En el centro de los bulbos existe un meristemo vegetativo o un vástago floral.
Por su consistencia existen dos tipos de bulbos: 1) los tunicados, que están cubiertos por escamas secas y membranosas que protegen al bulbo y le dan una estructura más o menos sólida. A esta clase pertenecen la cebolla y el tulipán; 2) los no tunicados, que no presentan la cubierta seca y sus escamas están separadas y unidas a la placa basal. Este tipo de bulbos daña fácilmente por lo que deben ser manejados con cuidado.
4) Pseudobulbos. Esta estructura vegetativa se da en la familia de las orquídeas. Son crecimientos tuberosos del tallo completo o de parte de éste o de las ramas. En las axilas de las escamas de la base de los pseudobulbos se forman vástagos nuevos o pseudobulbos en serie que conectan segmentos del tallo (figura 16A).
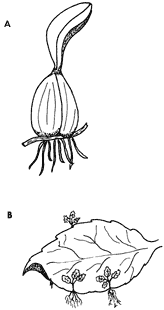
Figura 16. A) Pseudobuldo, y B) producción de nuevas plantas en el margen
de la hoja, como en la planta millonaria.
5) Turiones. Se presentan generalmente en especies acuáticas como estructuras de resistencia a condiciones ambientales adversas; se conocen también como yemas de invierno y se forman a partir de yemas que se desprenden del tallo o que persisten cuando el resto de la planta muere. Las hojas del turión contienen elementos de reserva y cuando se restablecen las condiciones favorables se inicia la producción de las raíces adventicias.
6) Chupones. Son estructuras que se forman en las axilas de las hojas escamosas de los tallos subterráneos y de los rizomas, o de las yemas adventicias de las raíces. El chupón forma varios entrenudos cortos; después de formar uno o más nudos desarrolla raíces adventicias y puede formar una nueva planta. El plátano y el bambú forman chupones.
Propagación vegetativa por raíces
Una forma extensa de propagación de las plantas se da mediante numerosos brotes que crecen de sus raíces horizontales. Tales brotes se forman sólo si la raíz es dañada, entonces los brotes se diferencian en un tejido calloso. Las raíces carnosas y aglomeradas de los camotes, las dalias y las peonias son también un medio de propagación vegetativa.
Propagación vegetativa por hojas
Este tipo de propagación no es tan frecuente en la naturaleza como los dos anteriores. Sin embargo, es posible encontrarlo en las hojas de algunos helechos, que forman una especie de acodadura al entrar en contacto con el suelo; en otras especies, entre las que se encuentran las violetas africanas, se forman nuevos individuos a partir de las hojas que se desprenden y caen al suelo y que posteriormente desarrollan raíces adventicias (figura 16B).
Propagación vegetativa por estructuras florales
En algunas plantas los meristemos apicales que normalmente se desarrollarían como flores se convierten en yemas vegetativas asociadas con raíces adventicias. Estas estructuras crecerán independientemente al ser liberadas de la planta progenitora. La producción de yemas vegetativas en lugar de flores se conoce como prolificación o falsa viviparidad.
Los bulbilos son yemas axilares de consistencia carnosa que almacenan reservas. En algunas especies de cebolla los bulbilos se forman en lugar de las flores, mientras que en algunas especies de agave las inflorescencias son reemplazadas por cientos de bulbilos. El nombre de estas estructuras de propagación vegetativa se debe a que visualmente se parecen a los bulbos, pero su color es verde; cuando están sobre la planta progenitora carecen de raíces adventicias, las cuales se desarrollan cuando los bulbilos son liberados.
PROPAGACIÓN VEGETATIVA INDUCIDA
Como hemos visto, la potencialidad de las plantas para generar nuevos individuos
a partir de segmentos de su organismo está distribuida ampliamente en las plantas
de muchos ambientes. Para muchas especies la reproducción asexual predomina
sobre la sexual, y es que las condiciones de su ambiente hacen muy improbable
que la semilla llegue a generar una planta capaz de establecerse debido a las
limitaciones de recursos fundamentales como el agua, la luz o la competencia
con las plantas establecidas. Un caso bien conocido en nuestro país es el de
las cactáceas y otras plantas de las zonas áridas que presentan muchas de las
estructuras reproductivas antes citadas. Por ejemplo, los nopales se reproducen
fácilmente en forma natural a partir de segmentos del tallo, que tienen una
forma muy peculiar y se les conoce como pencas, y en términos botánicos como
cladodios. Éstos se desprenden espontáneamente o a consecuencia de algún hecho
traumático y enraizan en forma natural, lo que constituye en muchos casos el
principal mecanismo de reproducción de estas plantas (figura 17).
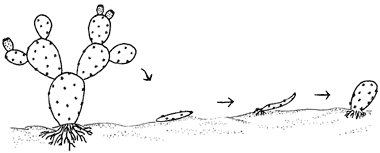
Figura 17. Propagación vegetativa por medio de los cladodios de plantas
de nopal (segmentos del tallo).
Con base en la potencialidad presente en la naturaleza en lo que respecta a la propagación vegetativa de las plantas, se han desarrollado métodos de propagación inducida, cuya complejidad va desde las tecnologías más rústicas hasta los métodos más tecnificados.
Esta técnica de propagación tiene muchas ventajas y se emplea exitosamente sin necesidad de gran inversión económica. La técnica más común es la inducción de la formación de raíces en una sección del tallo o de la rama, de manera que se origine una planta independiente. En los casos en que se ha experimentado propagar árboles mediante la enraización a partir de segmentos se ha tenido éxito en más de 80 por ciento.
Según la parte de la planta de donde se obtienen los segmentos (cortes o fragmentos) se ha dividido en cortes de: hojas, de brotes o renuevos, de raíz y de ramas. La selección de cualquiera de ellos depende básicamente de las características inherentes a cada especie, de las facilidades para obtener y manipular los cortes (en función del estado fenológico de la planta), del propósito de la propagación y de la disponibilidad de recursos económicos.
A continuación se mencionan algunas de las características de los diferentes tipos de cortes:
1) Cortes de hojas. Algunas especies herbáceas, como las violetas africanas y las peperomias, producen raíces a partir de sus hojas y posteriormente tallos; sin embargo, esto no ocurre con facilidad en la mayoría de los árboles. Los cortes que incluyen además de la hoja una yema axilar y un fragmento de rama son adecuados para propagar algunas plantas —como las camelias y los rododendros, que son especies leñosas— y también se utilizan para propagar árboles cuando la cantidad disponible de otro tipo de segmentos es escasa.
2) Cortes de raíz. La capacidad de muchos árboles de producir ramas a partir de sus raíces (en condiciones adecuadas de crecimiento) se utiliza para propagar algunas plantas, como los plátanos y los guayabos.
3) Cortes de ramas. La propagación vegetativa mediante segmentos de ramas o brotes es uno de los métodos más usados para propagar plantas leñosas en vivero. Según las características de madurez de la madera de donde se obtienen las ramas o brotes, los cortes se han dividido en cortes son: de maderas duras, semiduras y suaves. Aunque las diferentes fases de maduración se presentan de manera continua, generalmente se distinguen por la forma y el color de las hojas y por los cambios de coloración del tallo o ramas. Las técnicas de propagación de árboles por medio de cortes de ramas se dividen en dos tipos básicos: de segmentos foliados y de segmentos defoliados. Cada uno de éstos utiliza cortes de madera con un grado de maduración diferente, y como proceden de árboles de contrastante ciclo fenológico, esta diferencia se relaciona con la acumulación de reservas en los tejidos del tallo. En los árboles caducifolios, de los cuales se obtienen los segmentos defoliados, antes de la caída de las hojas hay acumulación de reservas, las cuales están destinadas a formar posteriormente hojas nuevas. A partir de estas reservas se generan las raíces y las hojas en el segmento; en cambio, los segmentos foliados por lo general proceden de árboles de hoja perenne, que no acumulan reservas en el tallo y que deben continuar fotosintetizando para producir los recursos necesarios para generar nuevo crecimiento.
Enraizamiento de segmentos defoliados
Esta técnica de reproducción vegetal se da espontáneamente en la naturaleza cuando una rama o fragmento de una planta cae al suelo y logra enraizar otra vez y producir así un nuevo individuo. También se le ha empleado desde tiempos inmemoriales por los horticultores para la propagación de árboles de ornato y frutales; un ejemplo de este método son las cercas vivas de palo mulato o colorín, que vemos alrededor de potreros y cultivos en el trópico mexicano. Para la construcción de estas cercas los campesinos cortan los segmentos o ramas al efectuar los desmontes, los almacenan en un lugar fresco y sombreado y los plantan al principio de la época de lluvias.
Los árboles que más fácilmente se propagan de esta manera son los que presentan una fase fenológica de defoliación y latencia meristemática al final de la época favorable para el crecimiento, como muchos árboles de las selvas bajas caducifolias y subcaducifolias, los cuales tienen la posibilidad de enraizar a partir de segmentos defoliados; por ejemplo, el cacahuananche, el palo mulato, el ciruelo amarillo de tierra caliente y el colorín, entre otras. En ocasiones, esta forma de propagación también es adecuada para algunas leñosas no deciduas de hojas angostas.
El método consiste básicamente en cortar ramas o pencas y plantarlas en el suelo húmedo para provocar su enraizamiento. Este ocurre fácilmente sin necesidad de emplear sustancias enraizadoras, ya que al encontrarse en un estado de latencia meristemática, al volver al estado de crecimiento los propios cambios hormonales que ocurren en el segmento desencadenan la producción de raíces en la superficie que está en contacto con el suelo. Los cortes se obtienen de ramas de crecimiento de la estación anterior y se realizan cuando la etapa de crecimiento cesa y la abscisión de hojas se ha presentado (finales de otoño o en el invierno). Debido al tamaño de los segmentos y a las condiciones de lignificación de la madera (madera dura), los cortes no se deshidratan y conservan la humedad el tiempo suficiente para generar un nuevo crecimiento de raíces y ramas.
A continuación se enumeran los pasos y criterios que se deben considerar para realizar esta actividad:
1) Seleccionar donantes vigorosos y sanos con alta cantidad de reservas alimenticias, preferentemente de un banco de plantas donantes que han crecido en condiciones de completa iluminación y que por lo tanto contienen alta cantidad de reservas alimenticias.
2) Elegir los segmentos basales o centrales de la rama, que son los que tienen más reservas alimenticias necesarias para el desarrollo de las nuevas raíces, pues de ellos se derivan las ramificaciones secundarias. Por ello no se deben elegir ramas con entrenudos muy largos o de ramas pequeñas y débiles.
3) El tamaño de los segmentos varía entre 15 y 75 cm de largo, el criterio adecuado para elegirlo depende de la especie, ya que se requiere que se incluyan por lo menos dos nudos, aunque lo recomendable es de cuatro a seis, sobre todo cuando los entrenudos son muy cortos. El diámetro de las ramas en que se realizan los cortes puede ser de 0.6 a 5 centímetros.
4) El corte basal se hace justo abajo de un nudo (sitio donde preferentemente se forman raíces adventicias) y el corte superior se realiza de 1.3 a 2.5 cm arriba del otro nudo. El corte puede ser de mazo (incluye una sección del tallo de madera más vieja), de talón o tacón (la porción de madera vieja es más pequeña) y el recto (no incluye madera vieja) (figura 18).
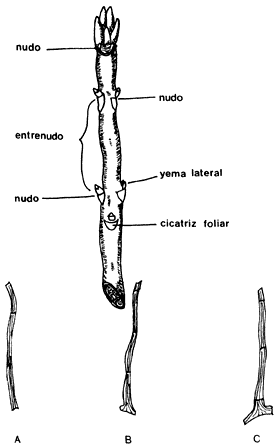
Figura 18. Se señalan en la parte superior las partes de una estaca y en
la porción inferior los tipos de corte utilizados para la obtención de estacas:
a) recto, b) talón o tacón y c) mazo.
5) Empaquetar las estacas cuidando su orientación, para mantener su polaridad y permitir que el flujo de savia siga su dirección normal. Por eso se marca la base con un corte sesgado o se baña la base con cera, lo cual ayuda también a evitar la pérdida de humedad, que podría propiciar enraizamientos pobres.
6) El enraizamiento de segmentos defoliados ocurre fácilmente, ya que el propio ciclo fenológico hace coincidir la producción de hormonas de crecimiento con el periodo de enraizamiento y crecimiento de yemas del segmento. Aun así, se favorece notablemente el enraizamiento si se emplean hormonas y algunos procedimientos para asegurar el desarrollo rápido de los segmentos. Las sustancias más usadas para acelerar el enraizamiento son el ácido naftalenacético (ANA) y el ácido indolbutírico (AIB), de los cuales se hablará posteriormente. El enraizamiento también se favorece colocando los segmentos a temperatura baja (5-8°C) por algunas semanas, ya que esto estimula la síntesis de hormonas en plantas que proceden de climas en los que hay una estación fría.
Finalmente, para lograr un buen enraizamiento hay que escoger los segmentos con las características óptimas de madurez de la madera y que carezcan de hojas.
7) Para la siembra se eligen terrenos que reúnan las características sugeridas en el capítulo III.
Algunas opciones para preparar y manipular las estacas antes de la plantación se presentan en el cuadro 22.
CUADRO 22. Métodos de preparación y manipulación de los segmentos
o cortes defoliados previos a su plantación definitiva.

Enraizamiento de segmentos foliados
Según las condiciones de lignificación de la madera los segmentos foliados se dividen en: cortes de maderas blandas (meristemos), cortes de maderas semiduros (parcialmente maduros) y cortes de maderas duras siempre verdes (lignificados). Cualquiera de éstos llevan hojas o brotes meristemáticos (material fisiológicamente juvenil) y su tamaño es mucho mas pequeño que los defoliados. Las especies que se propagan con esta técnica son la caoba, el zapote negro, el chicozapote y el barí, entre muchos otros más.
Debido a la presencia de hojas que continúan transpirando activamente y a las condiciones diferenciales de madurez en las ramas jóvenes, los segmentos pueden deshidratarse muy fácilmente. Por esto, es necesario mantenerlos en compartimientos sombreados y húmedos hasta que enraizan y toman del suelo suficiente agua. Siempre es necesario emplear sustancias enraizadoras como el ácido indolbutiírico (AIB) o el naftalenacético (ANA) y con frecuencia es indispensable mantener los segmentos recién plantados bajo agua nebulizada para evitar la deshidratación.
Segmentos foliados de maderas blandas
Los cortes se seleccionan en la estación de crecimiento, antes de que se presente la lignificación, y son tomados de los renuevos tempranos o de ramas jóvenes generadas a finales de la primavera y principios del verano. Esto significa que las ramas se encuentran en crecimiento activo, por lo que en su acopio y manipulación se deben tener cuidados especiales. La clave para el éxito es mantener los cortes turgentes, desde el periodo de colecta hasta que son puestos a enraizar en dispositivos diseñados especialmente para que las estacas mantengan la turgencia.
Se ha encontrado en varias especies que los segmentos provistos de meristemos cercanos al tronco principal, o al eje de crecimiento apical tienen mayores probabilidades de enraizar que las puntas de las ramas distantes. Otras veces, los ejes de crecimiento que surgen de un ápice podado son los que presentan mayor potencial de enraizamiento. Esto varía en función de la especie y de la técnica utilizada, por lo que se debe probar cuál es la fuente de material que origina las estacas juveniles (suculentas), no lignificadas, más adecuadas. Pueden ser: rebrotes de tocones, rebrotes basales de árboles en pie, rebrotes de plantas jóvenes y ápices de árboles podados. En cualquiera de estos casos se produce cierto número de brotes laterales con crecimiento vertical (los que no presentan crecimiento vertical deben desecharse), que se utiliza como fuente de estacas. Se deben considerar tres aspectos para realizar este tipo de propagación: a) la elección y manejo de la planta donante, b) la obtención de las estacas y c) el enraizamiento y establecimiento del segmento. A continuación se detallan estos puntos:
Elección y manejo de la planta donante
Las plantas donantes deben ser vigorosas, sanas y estar sujetas a un buen manejo para asegurar la producción continua y prolongada de gran número de estacas de fácil enraizamiento.
Se pueden cosechar brotes de una misma planta donante cada dos o tres meses, pero no se recomienda hacer cosechas muy frecuentes, pues se afectarían las reservas alimenticias de la planta, su sistema radicular y la fertilidad del suelo.
La planta donante debe ser fertilizada con regularidad y mantener por lo menos una rama con hojas que pueda continuar fotosintetizando y que de esta manera sirva como brote alimentador para la planta donante. También debe mantenerse en la sombra, al menos por unas semanas, lo cual favorecerá el futuro enraizamiento de las estacas, ya que en esta situación la planta no padece estrés hídrico.
Para obtener y manipular adecuadamente las estacas deben tomarse en cuenta varios factores: la alta humedad del aire, la intensidad moderada de luz, con temperaturas estables, un medio favorable de enraizamiento, y una protección adecuada contra el viento, las pestes y las enfermedades. Sobre todo debe evitarse la deshidratación, pues los cortes con hojas pierden rápidamente agua por medio de la transpiración, aun cuando exista una alta humedad relativa. Y es que, como no tienen raíces, la absorción de agua es mucho más lenta, y esto afecta el estado de hidratación de la estaca.
A continuación presentamos unas recomendaciones para obtener los cortes de la planta donante:
1) La obtención de ramas de la planta donante debe realizarse por la mañana o por la tarde (antes de las 10 am o después de las 4 pm), con la finalidad de evitar la pérdida de agua durante las horas de mayor insolación.
2) Es conveniente que la poda de las ramas elegidas (con crecimiento vertical) se realice a la altura de los 10 nudos o menos, como en el caso de los brotes obtenidos de tocones. Cuando se dificulte distinguir el número de nudos es recomendable tomar como criterio una altura del brote o rama, desde 10 cm hasta 1 m, para asegurar una mayor capacidad de enraizamiento (figura 18).
3) Las hojas de las ramas de donde se obtendrán los cortes deben tener entre 8 y 10 cm de largo, de lo contrario hay que reducir el área foliar, debido a que hojas muy grandes favorecen la pérdida de agua y las muy pequeñas no producen suficientes carbohidratos u otras sustancias necesarias para que el corte sobreviva. Se puede reducir el área foliar cortando las hojas con unas tijeras y cuidando que el tejido no se dañe por machacamiento o estrujamiento (figura 19).
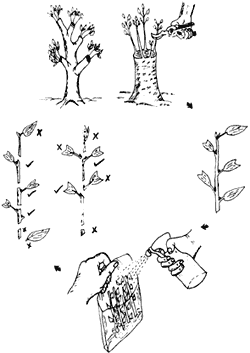
Figura 19. Proceso de obtención de estacas de la planta donante y manipulaciones
de los cortes. Las estacas más fácilmente enraizables son las que se obtienen
de rebrotes en troncos cortados. Hay formas correctas e incorrectas de resegmentar
los rebrotes que se muestran a la izquierda. Es importante mantener húmedos
los segmentos durante su transporte.
4) Ya cortados los brotes se marcan con el número de la planta donante (número de clon), se introducen lo más rápidamente posible en bolsas de plástico con algún material que retenga bastante agua y se cierran para evitar la pérdida de humedad. Deben mantenerse en un sitio fresco y sombreado y en cuanto sea posible se trasladan al área de enraizamiento del vivero.
5) Al extraer los brotes para hacer los cortes deben mantenerse húmedos y frescos, exponiéndolos lo menos posible al viento, ya que éste incrementa la pérdida de humedad. Los cortes deben hacerse con instrumentos filosos, en forma oblicua por arriba del nudo, o bien rectos para evitar que el sistema radicular se forme de un sólo lado. La longitud óptima de las estacas es usualmente entre 3 y 10 cm. Independientemente del tipo de corte o tamaño, éstos siempre deberán contar al menos con una hoja en la punta de la estaca, para que ésta proporcione nutrientes y otras sustancias necesarias para el enraizamiento.
Enraizamiento y establecimiento
El área donde se colocarán las estacas para el enraizamiento debe ser fresca y sombreada. La temperatura óptima para que ocurra se encuentra entre los 20 y 25°C. Cuando las temperaturas suben arriba de 30°C la humedad relativa de la atmósfera o contenido de vapor de agua presente en el aire tendrá que ser muy alto (más de 90%) para impedir que las plantas pierdan demasiada agua al incrementarse su transpiración y terminen marchitándose. La sombra se puede producir con materiales de origen vegetal como hojas de palma, paja, ramas secas, o con mallas plásticas especiales diseñadas para ese propósito. Es importante que el material utilizado transmita una luz que sea apropiada para activar la fotosíntesis de las plantas.
1) Inducción del enraizamiento
Como se mencionó, no todas las plantas tienen la capacidad de enraizar espontáneamente, por lo que a veces es necesario aplicar sustancias hormonales que provoquen la formación de raíces. Las auxinas son hormonas reguladoras del crecimiento vegetal y, en dosis muy pequeñas, regulan los procesos fisiológicos de las plantas. Las hay de origen natural, como el ácido indolacético (AIA), y sintéticas, como el ácido indolbutírico (AIB) y el ácido naftalenacético (ANA). Todas estimulan la formación y el desarrollo de las raíces cuando se aplican la base de las estacas.
La función de las auxinas en la promoción del enraizamiento tiene que ver con la división y crecimiento celular, la atracción de nutrientes y de otras sustancias al sitio de aplicación, además de las relaciones hídricas y fotosintéticas de las estacas, entre otros aspectos. La mayoría de las especies forestales enraizan adecuadamente con AIB, aunque se ha observado que para algunos clones la adición de ANA resulta más benéfica.
Un método sencillo es la aplicación de la hormona por medio del remojo de la base de las estacas (de 2 a 3 cm) en soluciones acuosas y con bajas concentraciones de auxina (de 4 a 12 horas), según las instrucciones de los preparados comerciales. Sin embargo, este método es lento y poco exacto, difícil de realizar cuando los cortes son numerosos y algunas veces las hojas se marchitan durante el proceso; entonces se puede recurrir a las auxinas disponibles en aerosol.
Para las especies forestales tropicales se recomienda la inmersión de la base de las estacas en soluciones de AIB al 4% en alcohol etílico como solvente, por periodos muy cortos (5 segundos). Posteriormente se acomoda la base de la estaca en aire frío para evaporar el alcohol, antes de colocarlas en el propagador (figura 20).
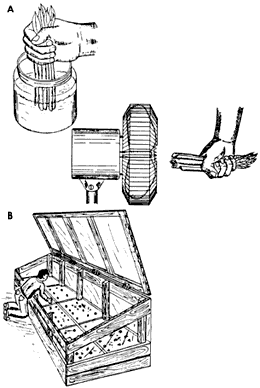
Figura 20. Aplicación de auxinas solubilizadas en alcohol a los segmentos,
evaporación del alcohol de los segmentos y siembra en el propagador rústico.
2) Propagadores y medios de enraizamiento. El ambiente en el cual las estacas son puestas a enraizar es de vital importancia. Los propagadores deben reunir características que eviten cualquier desecación en las estacas.
Un propagador es una construcción que evita la pérdida de agua del medio que rodea a las estacas. Su función es similar a la de un almácigo, pues ambos propician las condiciones ambientales adecuadas para la germinación y establecimiento de las plántulas o para el enraizamiento de las estacas, según sea el caso de que se trate.
Hay propagadores con sistemas de aspersión de alto costo que regulan automáticamente la frecuencia y la intensidad de la aspersión. Se instalan en invernaderos con control de luz y humedad. Sin embargo, la humedad también se puede controlar de manera sencilla en un compartimiento que tenga una tapa transparente para permitir el paso de la luz y evitar la pérdida de humedad; el fondo del compartimiento se cubre con una mezcla de arena y grava saturadas de agua, sobre la cual se pone el medio de enraizamiento. Adicionalmente se debe reducir la insolación del dispositivo y dar aspersiones manuales periódicas (figura 21).
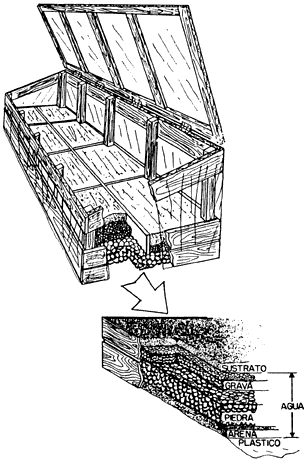
Figura 21. Aspecto del propagador rústico y composición del medio de enraizamiento
para las estacas.
3) Sustrato de enraizamiento. Un buen medio de enraizamiento se obtiene con arena gruesa o grava fina, que debe estar limpia (aunque no necesariamente estéril) húmeda y bien aereada. Si su capacidad de retención de agua es baja se puede mejorar adicionando aserrín (no demasiado fresco), turba, vermiculita u otros materiales (véase el capítulo III). En el caso de haber inicios de pudrimiento en las estacas será necesario aplicar algún fungicida al medio de enraizamiento.
4) Siembra de las estacas en el propagador. Las estacas ya preparadas se siembran rápidamente pero tomando en cuenta las siguientes indicaciones: los cortes deben colocarse a una profundidad de 2 a 3 cm; para asegurar que queden firmes es necesario compactar un poco el sustrato de enraizamiento; cuando se utilizan estacas multinodales con varias hojas se debe evitar que las hojas inferiores queden en contacto con el medio de enraizamiento para evitar la putrefacción.
5) Trasplante y acondicionamiento de las estacas. En varias especies propagadas vegetativamente se ha observado que el enraizamiento de las estacas se inicia después de dos semanas, y está lo suficientemente desarrollado después de 4 a 6 semanas (cuando las raíces miden de 1 a 2 cm) (figura 22). Las estacas que enraizan en tiempos más largos son débiles y no deben conservarse. El trasplante de las estacas tiene que hacerse inmediatamente después de ser removidas del medio de enraizamiento. Al sacar las estacas de su medio hay que tener cuidado de no dañar las raíces, despúes se verifica que el sistema radical tenga tres raíces como mínimo y que su distribución sea radial. Cuando las estacas presenten una o dos raíces, o bien cuando el sistema radical se forme sólo de un lado se deben desechar, para no poner en riesgo el vigor o una adecuada forma de crecimiento.

Figura 22. Segmento foliado enraizado.
Posteriormente se pasan a recipientes que contengan sustrato aereado y con buena fertilidad. Es recomendable agregar tierra del sitio donde naturalmente crece la especie para así favorecer la inoculación de la microflora apropiada. Es necesario estabilizar los trasplantes adecuadamente, para lo cual los envases deben llenarse con el medio de crecimiento aproximadamente a la mitad de su capacidad. La estaca se coloca en el envase en posición correcta (con la yema al ras del suelo y en su mayor parte dentro del medio del envase) y se termina de llenar. Esto ayuda a que no queden espacios de aire en su base y a que las raíces no se dañen, lo que asegura que éstas queden bien distribuidas en el envase (sin curvaturas o enrollamientos). Cuando hay más de una yema se recomienda eliminar algunas con el fin de asegurar la formación de plantas con un sólo eje y favorecer que el eje se desarolle en forma recta (figura 23).
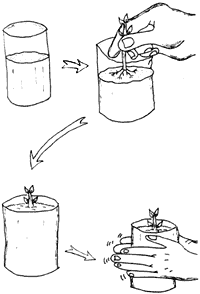
Figura 23. Proceso de trasplante de estacas del propagador a los envases
de crecimiento.
Algunas estacas recién enraizadas se deshidratan al pasarlas directamente al medio externo, por lo que se recomienda dejar los envases unos días más en el propagador, protegiendo a éste con plástico para evitar su contaminación con el material de los envases. En el periodo en que las estacas se aclimatan a las condiciones ambientales que existen fuera del propagador es conveniente colocarlas primero en un ambiente sombreado y húmedo por dos o tres semanas, y después exponerlas paulatinamente a condiciones decrecientes de humedad y crecientes de luz y temperatura.
Segmentos foliados de maderas semiduras y segmentos foliados de maderas duras siempreverdes
La diferencia entre este tipo de segmentos y los de maderas blandas estriba en las condiciones de lignificación de las ramas y en la época del año en que éstas se obtienen de la planta donante. El uso de esta técnica es muy importante en la propagación vegetativa de varias especies de pino, las especies siempre verdes de hoja angosta con maderas duras, y en la propagación de árboles siempreverdes de hoja ancha. Para el enraizamiento de este tipo de segmentos hay que seguir las mismas indicaciones que se mencionaron para el manejo y selección de brotes en las plantas donantes de maderas blandas, así como también para la manipulación y cuidado de los cortes y evitar así el estrés hídrico en las estacas. Sin embargo existen algunas diferencias en el manejo de los cortes de especies siempreverdes de hoja ancha y de hoja angosta.
Especies siempreverdes de hojas anchas
En el caso de los segmentos de maderas semimaduras, las ramas de especies siempreverdes de hojas anchas se obtienen en el verano, cuando el crecimiento empieza a declinar y la madera se encuentra parcialmente madura. Los cortes pueden tener longitudes de 7.5 a 15 cm, y las hojas de la punta de la estaca se conservan, si éstas son muy grandes se debe disminuir su tamaño (figura 24).

Figura 24. Segmentos foliados siempreverdes de hojas angostas y de hojas anchas.
Segmentos de maderas duras siempreverdes de hojas angostas
Para los segmentos de maderas duras siempreverdes de hojas angostas el enraizamiento toma mucho tiempo (desde varios meses hasta un año) y es muy importante evitar que las estacas sufran estrés hídrico durante este periodo. Los cortes se obtienen de plantas donantes jóvenes, ya que este factor es determinante para tener éxito. Generalmente los cortes se realizan en ramas terminales, de la época de crecimiento pasada, a finales de otoño o bien en invierno. La longitud de las estacas varía de 10 a 20 cm, y hay que tener cuidado de remover las hojas de la mitad de la estaca hacia la base (figura 24).
A diferencia de los segmentos foliados de maderas blandas estos dos últimos
tipos de cortes requieren un medio de enraizamiento que incluya mezclas de vermiculita,
y turba en proporción 1:1, o bien de perlita y vermiculita que proporcionan
nutrientes y mayor humedad. Además, el enraizamiento se puede favorecer aplicando
calor a la base de las estacas (24 a 26.5°C). Esto es particularmente favorable
para las especies de hoja angosta. Asimismo, para estos últimos cortes la aplicación
de AIB a altas concentraciones es usualmente benéfico para incrementar
la velocidad y porcentaje de enraizamiento, además de favorecer el desarrollo
de sistemas radiculares robustos.
Ambos tipos de cortes, los de hojas anchas y angostas, enraizan mejor en invernaderos con sistemas de aspersión automáticos o bajo propagadores rústicos, en condiciones de alta iluminación y humedad relativa, o bien, bajo ligeras nebulizaciones.
La técnica de injerto consiste en tomar un segmento de una planta, por lo general leñosa, e introducirlo en el tallo o rama de otra planta de la misma especie o de una especie muy cercana, con el fin de que se establezca continuidad en los flujos de savia bruta y savia elaborada, entre el tallo receptor y el injertado (figura 25). De esta manera, el tallo injertado forma un tejido de cicatrización junto con el tallo receptor y queda perfectamente integrado a éste, pudiendo reiniciar su crecimiento y producir hojas, ramas y hasta órganos reproductivos. Esta técnica es muy antigua y ya era practicada por los horticultores chinos desde tiempos remotos. Tiene grandes ventajas, sobre todo para el cultivo de árboles frutales y de ornato, pues permite utilizar como base de injerto plantas ya establecidas que sean resistentes a condiciones desfavorables y enfermedades, utilizándolas como receptoras de injertos de plantas más productivas y con frutos de mejor calidad y mayor producción.
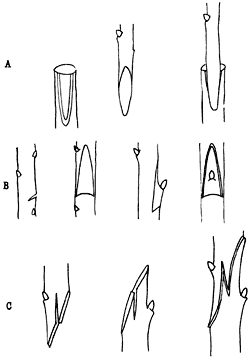
Figura 25. Existen diversas técnicas de injerto que permiten la unión de
segmentos de dos individuos diferentes: a) injerto de una rama sobre una incisión
lateral de un tallo, b) injerto de una yema en la corteza de un tallo, c) injerto
de un tallo sobre otro.
Al contrario de lo que generalmente se cree, el injerto no produce una combinación de características entre la planta receptora y la injertada. Los frutos de la planta injertada no cambian sus propiedades ni su sabor, la única ventaja es que el injerto permite utilizar bases ya desarrolladas, lo que acelera la producción de frutos. También se aprovecha la resistencia (a enfermedades o condiciones desfavorables) de variedades de árboles frutales de menor calidad, para injertar en ellos variedades de mayor calidad pero menos resistentes.
El injerto es una forma particular de reproducción asexual por segmentos que se utiliza en gran escala en la fruticultura. Una de las industrias que recurren con mayor frecuencia a esta técnica es la vitivinicultura o cultivo de la vid. Con gran frecuencia las plantas productoras de uvas de baja calidad, pero muy resistentes a la sequía y a las enfermedades, son injertadas con segmentos de vides de alta producción y calidad. Esta técnica es muy empleada para mejorar la producción de viñedos antiguos ya establecidos desde hace mucho tiempo.
Las técnicas de injerto son muy variadas y existe un método óptimo para cada propósito y tipo de planta. En esencia todos los procedimientos consisten en tener a la disposición buenas plantas receptáculo y buenos segmentos que injertarle. La técnica se inicia haciendo un corte en el tallo receptor y otro en el segmento a injertar, para que hagan contacto los tejidos vasculares del injerto con sus equivalentes en la planta receptora. Una vez realizado el injerto se protege la herida con una cera especial y se cubre con tela o con una cuerda para evitar que se desprenda. El tejido injertado por lo general está defoliado y es conveniente que el injerto coincida con la época de primavera para que, al reiniciar el crecimiento de los tejidos, los estímulos hormonales que caracterizan ese momento de la vida de la planta induzcan el establecimiento de una conexión apropiada entre los tejidos de ambas partes.
Para que un segmento pueda injertarse sobre otra planta, como ocurre en los animales que reciben injertos de órganos, tiene que haber una afinidad entre los tejidos que van a ponerse en contacto para que el injerto no sea rechazado. Por esto el injerto sólo es posible entre plantas de una misma especie, aunque sean de diferentes variedades o razas. A veces es posible el injerto entre especies diferentes, siempre y cuando éstas sean muy cercanas entre sí, o sea, que por lo menos pertenezcan al mismo género.
El desarrollo de las técnicas de cultivo de tejidos vegetales, aunadas al descubrimiento de las hormonas de crecimiento y diferenciación de las plantas —auxinas y citoquininas—, permitió el desarrollo de varias técnicas de cultivo orientadas hacia la propagación de plantas que son ahora de uso corriente en muchos laboratorios de investigación del mundo y también en empresas de propagación comercial.
Estas técnicas están basadas en el hecho de que los tejidos vivos de las plantas conservan la potencialidad de dar origen a un organismo completo. Las células que conservan mejor esta potencialidad son las que están menos diferenciadas hacia una función específica, como las que están presentes en las yemas y en otros tejidos primarios de las plantas, por ejemplo los extremos de las raíces, los segmentos nodales, las semillas, el parénquima del tejido vascular foliar, el cambium y algunas partes florales.
La técnica de cultivo de tejidos se inicia con la toma de segmentos de plantas en crecimiento que se esterilizan y se cultivan en soluciones nutritivas especiales, con frecuencia gelificadas. A estos medios se incorporan combinaciones adecuadas de hormonas de crecimiento para obtener una proliferación celular en el segmento. A partir de esta proliferación puede ocurrir la formación directa de raíces y tallos que originen una o varias plantas nuevas completas (figura 26). También es posible inducir la multiplicación de las células germinales del explante para formar un cúmulo de tejido poco diferenciado llamado callo o una estructura peculiar carente de paredes celulares llamada protoplasto, cada uno de los cuales se emplea en diversos procedimientos de propagación.
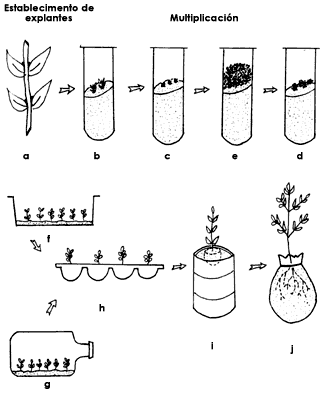
Figura 26. Del explante (a) se obtienen yemas (b) que se desarrollan formando
tallos adventicios (c, d, e). Estos tallos pueden aislarse para lograr su diferenciación
y después trasplantarse a recipientes con suelo (f-j).
El cultivo de tejidos y células vegetales tiene otras aplicaciones más o menos relacionadas con la propagación clonal. Entre éstas se encuentra la obtención de líneas de plantas genéticamente muy uniformes, con fines experimentales; el almacenamiento o transporte de germoplasma vegetal (de lo cual trataremos ampliamente más adelante); el control cada vez más importante de infecciones sistémicas crónicas en las plantas mediante la obtención de propágulos libres de virus y otros agentes infecciosos. En la ingeniería genética moderna el cultivo de tejidos tiene importancia en los procesos de transgénesis, hibridación interespecífica y en la generación y selección de variantes genéticas de las plantas.
Cada uno de estos temas es de gran complejidad y se necesitaría una obra mucho más extensa para hablar de ellos, lo cual no es el propósito de este libro. Aquí se verán los aspectos relativos al cultivo de células y tejidos vegetales exclusivamente en lo que respecta a las técnicas más sencillas de micropropagación de árboles actualmente en uso en laboratorios de tecnología sencilla en muchos lugares del mundo. Por mucho tiempo las técnicas de micropropagación han sido consideradas demasiado complicadas para convertirse en una opción viable para la propagación de árboles en regiones tropicales subdesarrolladas. Sin embargo, tal punto de vista es poco realista ya que existen métodos de micropropagación que sólo requieren recursos técnicos e instalaciones mínimas.
La bibliográfia científica moderna contiene innumerables trabajos que describen técnicas de cultivo de tejidos y órganos vegetales en medios complejos y finamente formulados, colocados en condiciones ambientales controladas con mucha precisión. Esto no sucede con las técnicas sencillas de micropropagación que rara vez se describen en los libros especializados disponibles. Yesto es un grave error, pues una vez que se ha probado que una especie de planta puede ser micropropagada exitosamente, los procedimientos de micropropagación se simplifican y se vuelven accesibles para las instalaciones sencillas que se encargan de la propagación de plantas.
Los procedimientos más usados por los laboratorios de micropropagación son cuatro métodos diferentes para multiplicar plantas in vitro: 1) la activación de la ramificación axilar, 2) los segmentos nodales, 3) los tallos adventicios y 4) las embriogénesis somáticas. De los cuatro, este último es el que se utiliza menos. Las técnicas que dan los mejores resultados varían dependiendo de las especies y del propósito por el cual se propagan.
1) Activación de la ramificación axilar (figura 27A). Quizá sea el método más frecuentemente usado; en él se emplean meristemos tanto terminales como axilares del tallo para establecer los cultivos. Manipulando el cultivo mediante cambios en su ambiente químico (hormonas) y en el fragmento que se cultiva, se suprime la dominancia apical y se induce la ramificación axilar. Subsecuentes subdivisiones del cultivo en fragmentos permiten multiplicar los explantes hasta alcanzar el número deseado de tallos. A pesar de que la multiplicación inicial es lenta, se incrementa rápidamente permitiendo varios subcultivos. Los tallos seccionados del cultivo pueden enraizar in vitro o directamente en el suelo.
2) Segmentos nodales (figura 27B). El método de los segmentos nodales tiene la ventaja de que genera poca variación genética en las muestras, ya que en ningún momento se forma tejido calloso. Se inicia escindiendo segmentos de tallo con su correspondiente yema axilar. En el cultivo las yemas crecen y se alargan produciendo nuevos segmentos nodales susceptibles de ser a su vez escindidos. En un momento determinado los segmentos nodales producidos pueden dar origen a tallos y raíces ya sea in vitro o en el suelo, mediante una alteración de la composición hormonal del medio. Muchas plantas no pueden ser propagadas exitosamente de esta manera.
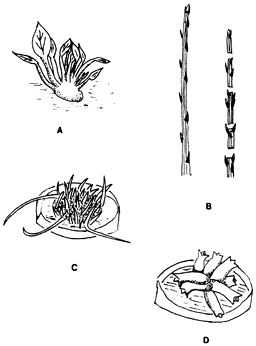
Figura 27. Tipos de propagación por cultivo de tejidos: A) activación de
la ramificación axilar, B) segmentos nodales, C) tallos adventicios y embriogénesis
somática.
3) Tallos adventicios (figura 27C). Pueden ser inducidos a partir de callos originados por explantes de ramas u hojas y continuarse cultivando hasta obtener un gran número de callos, simplemente cambiando el ambiente hormonal en el que se desarrolla el callo original o subsecuentes fragmentos de éste. Los tallos obtenidos pueden enraizar in vitro o en el suelo. De todos los métodos de propagación éste es el que genera más variabilidad genética debido a que se utiliza un estadio calloso intermediario, por lo que no se recomienda para la micropropagación de plantas en las que la integridad genética es muy importante. Al formarse el callo ocurre una desdiferenciación completa del tejido vegetal, que frecuentemente conlleva a la alteración genética, lo cual induce variabilidad en la información genética entre las células del callo.
4) Embriogénesis somática (figura 27D). Embriones somáticos o asexuales pueden resultar de un proceso de diferenciación directa o indirecta a partir de células, órganos o estados callosos intermedios de una planta. Muchas veces los embriones somáticos proliferan y producen embriones secundarios a partir de meristemos somáticos, como las yemas axilares, partes de las flores o de semillas. Estos embriones adquieren una morfología y comportamiento similar a los embriones naturales de origen sexual de las plantas y, como ellos, pueden germinar en el suelo. La embriogénesis somática puede convertirse en la forma más eficiente de propagación, ya que teóricamente se obtendrían millones de plantas a partir de una cantidad pequeña de tejido en cultivo.
La producción de embriones somáticos se ha estudiado a fondo sólo en un número muy pequeño de especies. Los embriones obtenidos de esta manera pueden utilizarse para producir semillas artificiales por medio del encapsulado con testas artificiales. Esta técnica es muy prometedora, ya que potencialmente se incrementaría de manera significativa la disponibilidad de propágulos de plantas valiosas de cultivo, se acortaría el ciclo de crecimiento y se desarrollarían nuevos métodos de conservación y almacenamiento de germoplasma.
La técnica para producir embriones somáticos se inicia con la producción de un tejido calloso que, mediante el uso de combinaciones especiales de sustancias químicas, da origen a un protoplasto en el que han desaparecido las paredes de la célula, asemejándose a un saco embrionario floral. Al colocar los protoplastos en el medio adecuado de cultivo comienzan a desarrollarse los embriones somáticos a partir del protoplasto.
Esta técnica ha sido aplicada con éxito a ciertos árboles tropicales, como el caucho, el mango y otros. Los embriones somáticos pueden incluso encapsularse en testas artificiales para originar semillas sintéticas que se cultiven en el suelo o utilizarse para la conservación de germoplasma vivo por mucho tiempo, utilizando técnicas de deshidratación y vitrificación, y conservación en nitrógeno líquido. Esto es posible incluso en plantas que originalmente producen semillas recalcitrantes.
Las técnicas de cultivo de tejidos requieren instalaciones y precauciones con cierto nivel de complejidad. La higiene y desinfección son muy importantes para evitar la contaminación microbiana de los cultivos. La formulación de los medios de cultivo es variada, sin embargo, éstos contienen fundamentalmente sales minerales de los macroelementos y oligoelementos como: calcio, potasio, magnesio, nitrógeno, fósforo, azufre, manganeso, hierro, cloro, sodio, zinc, boro, molibdeno y cobre, los cuales son indispensables para el crecimiento de las plantas. Además, algunos medios incorporan compuestos orgánicos sencillos como fuentes de carbono (generalmente carbohidratos), reguladores hormonales del crecimiento (auxinas, citoquininas y giberelinas), otros reguladores del crecimiento (aminoácidos y vitaminas) y a veces antioxidantes (ácido ascórbico).
En la figura 28 se indican todas las alternativas biotecnológicas que surgen a partir del cultivo de tejidos.
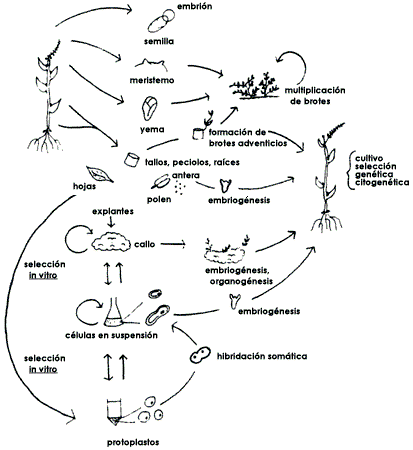
Figura 28. Opciones biotecnológicas para la propagación de plantas que se
originan a partir del cultivo de tejidos, en las que se incluyen también técnicas
de modificación genética.
La micropropagación y la propagación vegetativa permiten emplear técnicas de selección y mejoramiento de las características favorables de las plantas por medio de la selección clonal. Las características que pueden mejorarse cubren un amplio rango de posibilidades; por ejemplo, la resistencia de las plantas a la temperatura, a la sequía, a crecer en suelos pobres o con características desfavorables, como acidez o alcalinidad excesiva, salinidad alta o saturación de humedad; también puede mejorarse el rendimiento del forraje y frutos, su sabor y calidad nutricional, la velocidad de crecimiento, la calidad de la madera producida y la concentración de compuestos secundarios valiosos como sustancias químicas, látex, gomas, etc.
A continuación describimos brevemente las dos técnicas básicas de selección clonal: 1) Se buscan en la naturaleza las plantas que presenten la característica deseada en forma óptima (por ejemplo, los frutos más deliciosos y grandes), y se toma de ese individuo los meristemos o segmentos que se vayan a utilizar para la propagación vegetativa, para así obtener muchos individuos con la característica deseada. 2) Se recolectan semillas, segmentos o meristemos de muchos individuos de una o varias poblaciones de la especie que se desea propagar. Con este material se producen muchas plantas pequeñas en un vivero y se someten a las condiciones desfavorables para las que se desea que tengan mayor resistencia; también se puede comparar su velocidad de crecimiento, su producción de forraje o cualquier otra cualidad que se desee resaltar. Se escogen los individuos que muestren las características óptimas según el caso y se utilizan para propagarlos vegetativamente y obtener así individuos mejorados.
Esta técnica incluye la exploración de las diversas poblaciones de una especie en el medio natural, ya que en el área natural de distribución geográfica de una especie existe gran variación en muchos de los atributos deseables de la especie.
Siguiendo la segunda técnica descrita ha sido posible obtener cultivos de árboles tropicales con características muy favorables. Por ejemplo, especies de acacia resistentes a la salinidad o al suelo ácido; mezquites ornamentales resistentes al frío; especies de guaje productoras de abundante forraje de alta calidad y con rápido crecimiento en suelos pobres y, por último, árboles maderables cuyo crecimiento está determinado por una fuerte dominancia apical. Esto permitirá repoblar las selvas con individuos que producen un fuste o tronco recto y alto, muy apropiado para seguir técnicas óptimas de aserrado y uso en ebanistería, como es el caso del nogal africano y la caoba americana (figura 29).
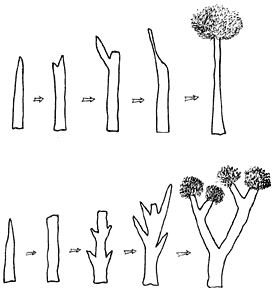
Figura 29. Selección clonal. En este caso se eligen los segmentos enraizados
que muestran la mayor dominancia apical, es decir, los que tienden a crecer
en un solo eje, y se desechan los que tienden a ramificar. De esta manera se
obtienen árboles con un fuste recto que permite un mejor uso industrial.
![[Inicio]](../img/begin.gif)
![[Anterior]](../img/prevsec.gif)

![[Siguiente]](../img/nextsec.gif)