I. ¿CÓMO FUNCIONA EL CEREBRO?: PRINCIPIOS GENERALES
| El cerebro de la iguana, el de Kant y el del hombre común funcionan con los mismos principios básicos |
EL CEREBRO, como todo en el organismo animal, está formado por
células, pero las del cerebro son excepcionales por su impresionante diversidad,
por la complejidd de sus formas, por la intrincadísima red que comunica a unas
células con otras. Algunas son modestamente estrelladas, otras recuerdan, por
su forma, a los animales marinos, calamares y medusas, otras tienen bifurcaciones
complejas, y otras más, en fin, exhiben increíbles penachos con ramificaciones
que se extienden en áreas muchas veces mayores que el cuerpo de la célula (figura
I.l). Las células del cerebro se llaman neuronas.1
![[Nota 1]](../img/mcommnt.gif) La estructura y la comunicación de las neuronas, en los albores de este siglo,
fueron descritas magistralmente por el sabio español Santiago Ramón y Cajal
—un gigante de la ciencia— quien encontró en el minucioso escudriñar
de las laminillas bajo el microscopio una característica fundamental de la comunicación
entre las células nerviosas: casi nunca se tocan, están separadas por pequeñísimos
espacios, cuyo significado y enorme importancia vendría a conocerse mucho tiempo
después. A pesar de las diferencias en la forma de las neuronas, su estructura
en los sitios en los que se comunican unas con otras es muy similar. La parte
de la neurona que "habla" con otra neurona tiene siempre una estructura típica,
y la región de la neurona que recibe ese contacto también tiene una forma característica.
A esta zona de interacción de las neuronas se le llama sinapsis (del
griego sunayiV = unión, enlace), y su funcionamiento
es esencial para explicar prácticamente todas las acciones del cerebro, desde
las más sencillas como ordenar a los músculos que se contraigan y se relajen
en forma coordinada para llevar a cabo un simple movimiento, hasta las más complicadas
tareas intelectuales, pasando también por las funciones que originan, controlan
y modulan las emociones.
La estructura y la comunicación de las neuronas, en los albores de este siglo,
fueron descritas magistralmente por el sabio español Santiago Ramón y Cajal
—un gigante de la ciencia— quien encontró en el minucioso escudriñar
de las laminillas bajo el microscopio una característica fundamental de la comunicación
entre las células nerviosas: casi nunca se tocan, están separadas por pequeñísimos
espacios, cuyo significado y enorme importancia vendría a conocerse mucho tiempo
después. A pesar de las diferencias en la forma de las neuronas, su estructura
en los sitios en los que se comunican unas con otras es muy similar. La parte
de la neurona que "habla" con otra neurona tiene siempre una estructura típica,
y la región de la neurona que recibe ese contacto también tiene una forma característica.
A esta zona de interacción de las neuronas se le llama sinapsis (del
griego sunayiV = unión, enlace), y su funcionamiento
es esencial para explicar prácticamente todas las acciones del cerebro, desde
las más sencillas como ordenar a los músculos que se contraigan y se relajen
en forma coordinada para llevar a cabo un simple movimiento, hasta las más complicadas
tareas intelectuales, pasando también por las funciones que originan, controlan
y modulan las emociones.
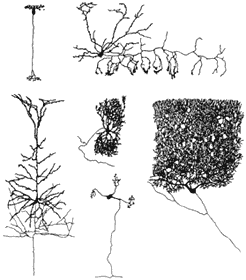
Figura I.1 Variedad de formas de las neuronas.
A través de esta comunicación las neuronas forman redes complicadísimas, que por supuesto estamos muy lejos de conocer por completo. Sabemos que algunos de estos circuitos están relacionados con el movimiento, otros con el sueño, y otros más con las emociones y la conducta. La identificación de estos circuitos puede lograrse con distintos métodos, pero uno relativamente simple consiste en estimular una neurona o un grupo de neuronas y luego tomar un registro en las neuronas que sospechamos se comunican con las primeras. Tanto la estimulación como el registro se llevan a cabo mediante los electrodos, los cuales son pequeñísimos tubos de vidrio que contienen soluciones que permiten el paso de la corriente eléctrica. A través del electrodo se hace pasar una corriente eléctrica muy pequeña, y si la neurona estimulada está en conexión con la que se está registrando, se advertirá una señal eléctrica. De esta forma pueden rastrearse los contactos funcionales entre las células nerviosas.
Los primeros circuitos funcionales identificados fueron los más sencillos, como aquellos que, partiendo de la corteza cerebral, terminan en distintos músculos del cuerpo. El procedimiento para su localización también fue muy rudimentario. Las observaciones pioneras en este campo se hicieron durante la guerra entre Prusia y Dinamarca, alrededor de 1864, cuando el médico alemán Theodor Fritsch se dio cuenta que al tocar algunas áreas descubiertas del cerebro de algunos heridos se producían movimientos musculares siempre en el mismo lugar. Terminada la guerra, al volver a la práctica médica en Berlín, él y un colega suyo, Eduardo Hitzig, comenzaron a diseñar experimentos para demostrar esta posibilidad. Como no contaban con instalaciones ni laboratorios equipados de ninguna naturaleza, hicieron sus experimentos en la casa del doctor Hitzig, utilizando perros a los cuales anestesiaban y estudiaban sobre la mesa de costura de la señora Hitzig, quien ciertamente debió ser una mujer muy tolerante.
Estos experimentos demostraron la localización de las funciones motoras en la corteza del cerebro y la existencia de conexiones neuronales desde ésa hasta los músculos. Otros investigadores prosiguieron esta tarea con más detalle y, suponemos, con mejores condiciones para realizar su trabajo. Fue así como se pudo identificar, primero en perros, luego en primates y finalmente en el hombre, cuáles son las áreas de la corteza cerebral que se conectan con los distintos músculos del cuerpo, de la cara y de las extremidades. Lo mismo se hizo para la percepción sensorial. Se observó en estos experimentos que el movimiento y la sensibilidad de algunas regiones del cuerpo requieren un mayor número de neuronas en la corteza, por ejemplo, las manos y la lengua. Se elaboró así el famoso mapa del "homúnculo" (hombrecito), reproducido en la figura I.2.
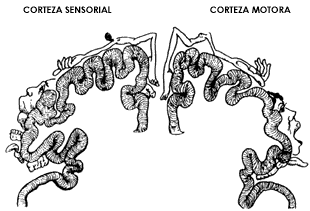
Figura I.2. Mapa del homúnculo (hombrecito). Área en la que se ubican las neuronas en la corteza sensorial y en la corteza motora que tienen bajo su control las distintas regiones del cuerpo, la cabeza y las extremidades.
Con estas bases, otros investigadores emprendieron la tarea de hacer un "mapa" de otras funciones localizadas principalmente en la corteza, y de esta forma se pudo determinar que existen áreas visuales (corteza visual), auditivas (corteza temporal) o para la percepción táctil (figura I.3). Con técnicas más elaboradas se localizaron también áreas de la corteza relacionadas con funciones más complejas, como la actividad intelectual, y también con la conducta. Estas últimas, sin embargo, están localizadas sólo parcialmente en la corteza cerebral y se encuentran más bien en otras estructuras del cerebro situadas debajo de la corteza. En particular, el conjunto de estructuras que se conocen como sistema límbico (punteado en la figura I.4A) tienen gran importancia en el origen y el control de las emociones. Dentro de este gran circuito, una pequeña región, el hipotálamo (figura I.4A), está asociada a muchas conductas emocionales y a funciones como el hambre y la sed. En efecto, en experimentos hechos con ratas, se ha podido observar que destruyendo algunos núcleos del hipotálamo (figura I.4B) —los núcleos son grupos de neuronas— el animal deja de comer y puede incluso morir de hambre literalmente en medio de la más apetitosa comida. Con estos estudios y otros similares se concluyó que a través de este núcleo es que se siente la necesidad de comer. Al ser destruidas las células de este núcleo, el animal tiene la continua sensación de estar lleno, y por tanto es incapaz de comer. A esta región del hipotálamo se le conoce como el centro de la saciedad. (Estos experimentos nos indican que las ratas no conocen el pecado de la gula, tan frecuente en la especie humana, ya que a diferencia de muchos de nosotros, el animal al sentirse saciado deja de comer.) En una región cercana a este núcleo de la saciedad se encuentra su opuesto, es decir un grupo de neuronas que, al ser destruidas, hacen que el animal pierda la capacidad de sentirse saciado y siga comiendo, sin cesar, hasta que no puede prácticamente moverse por la cantidad de alimento que ha ingerido. Por supuesto, estos núcleos del hipotálamo responden a señales, como el nivel de glucosa en la sangre que lo induce a alimentarse y que se encuentran bajo otras influencias nerviosas, principalmente de la corteza, incluidas las del origen del pensamiento y la imaginación. Así, sobre todo en el humano, el impulso de comer se puede modificar ante la vista o aun ante la simple evocación de alimentos apetitosos.
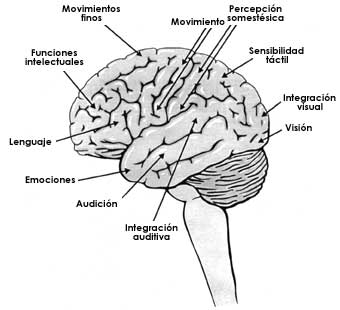
Figura I.3. Áreas en la corteza cerebral donde se localizan las neuronas relacionadas con distintas funciones.
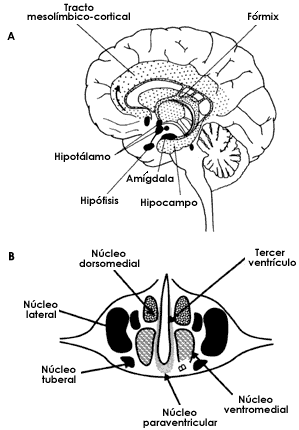
Figura I.4. A) localización del hipotálamo en el cerebro. El hipotálamo forma parte de un conjunto de estructuras cerebrales conocidas como sistema límbico (punteado) que participan de manera importante en la modulación de las emociones. B) en el hipotálamo se encuentra una serie de núcleos (grupos de neuronas) que tienen a su cargo funciones relacionadas con la generación, supresión y regulación.
También en el hipotálamo y en otras áreas del sistema límbico se localizan núcleos celulares que al ser estimulados provocan respuestas de cólera y agresividad en los animales, sin el concurso de los agentes externos que normalmente los causan, por ejemplo, la presencia de un ratón en el caso del gato. Estos núcleos del hipotálamo están modulados por influencias de la corteza y de otros centros que son los que determinan la amplitud y el vigor de la respuesta hipotalámica. En esta misma estructura nerviosa se localizan núcleos cuya función es más compleja que la del simple alimentarse, atacar o reproducirse. Esta posibilidad se derivó de las observaciones llevadas a cabo por James Olds y sus estudiantes en la Universidad McGill, en Canadá, en los años cincuenta. Estos investigadores se hallaban interesados en el estudio del sueño y la vigilia, y el diseño experimental para su investigación incluía la estimulación por medio de un pequeño electrodo en otra región del mismo hipotálamo y que el animal debía autoadministrarse pisando una palanca si quería recibir alimento como recompensa (figura I.5). Por error, en una ocasión el electrodo de estimulación fue implantado un poco más abajo de la zona deseada y, para sorpresa de los investigadores, al cabo del primer autoestímulo en esta región con el recurso de pisar la palanquita, la rata ya no tenía mayor interés en la recompensa o en explorar los espacios, sino que volvía una y otra vez a oprimir la palanca, y con ello a aplicarse el estímulo en el lugar del hipotálamo en el que se encontraba el electrodo. Evidentemente, los fisiólogos se percataron de inmediato de la importancia de su descubrimiento, y olvidando su proyecto anterior acerca del sueño se dedicaron a afinar y desarrollar una investigación acerca de este fenómeno asociado a lo que denominaron el núcleo del placer.

Figura I.5. Dispositivo experimental en el cual la rata puede autoadministrarse estímulos eléctricos o sustancias químicas apretando la palanca.
No parece ilógico extrapolar al ser humano estas observaciones hechas en el gato o la rata. Los científicos saben que las diferencias entre la especie humana y los otros animales no son tan grandes, en lo que se refiere a su comportamiento biológico, y que la enorme diferencia que evidentemente existe entre el gato y un ciudadano común, por no hablar de las mentes privilegiadas como Kant o Einstein, radica no en una diferencia en los principios generales con los que opera el sistema nervioso, que son exactamente los mismos, sino en la extrema complejidad de las conexiones interneuronales y tal vez en otros elementos que aún desconocemos. No hay que olvidar que el problema mente-cerebro, es decir, el de la localización celular de las funciones mentales superiores, no se ha resuelto, y es uno de los grandes retos de la neurobiología moderna.
Sin embargo, es posible imaginar, a la luz de estos sencillos experimentos, que la diferencia entre un individuo colérico y otro apacible puede ser que en el primero estos centros de la agresividad en el hipotálamo estén menos controlados por acciones inhibidoras de otras neuronas, o más activados por una preeminencia de neuronas excitadoras. El mismo razonamiento podría aplicarse a los centros hipotalámicos del hambre y la saciedad e imaginar que esa afición por la comida, que tenemos muchos de nosotros y que por supuesto y desafortunadamente se refleja en las redondeces de la figura, tenga una explicación, en parte, en el tipo de control que la corteza u otras estructuras ejercen sobre los núcleos del hipotálamo. No es tan descabellado suponer que la afirmación popular acerca del buen carácter de los gorditos tenga una base neurofisiológica a nivel del control de los núcleos del hipotálamo, relacionados con la regulación del apetito y con las distintas fases de la conducta agresiva.
La extrapolación podría parecer bastante simplista, pero no deja de tener su contraparte experimental cuando sabemos que la administración de ciertas drogas, como las anfetaminas, que precisamente actúan aumentando la eficiencia de algunas conexiones neuronales del tipo de las que se encuentran en el hipotálamo, da como resultado una pérdida casi total del apetito, además de modificar espectacularmente muchos rasgos del carácter del individuo, como veremos en otros capítulos. El esquema de que lo que consideramos una actitud emocional tiene su asiento en el sistema nervioso, va cobrando así cierta lógica.
LA CONVERSACIÓN ENTRE LAS NEURONAS
Si consideramos que la riqueza y la complejidad del pensamiento y del comportamiento humanos son, en buena medida, un reflejo de la comunicación que existe entre sus neuronas, se justifica que brevemente dediquemos nuestra atención a este tema.
Las neuronas tienen dos tipos de prolongaciones. Unas generalmente ramificadas, que confieren a estas células su aspecto estrellado o arborizado característico, y otras más largas y más sencillas, los axones, que son aquellas a través de las cuales las neuronas se comunican entre sí (figura I.6). La parte final del axón, que establece la comunicación con la neurona adyacente, se llama terminal sináptica o presinapsis, y se identifica en un gran número de sinapsis por la presencia muy característica de estructuras esféricas: las vesículas sinápticas (figura I.7) cuya función es clave para la comunicación interneuronal, como se verá más adelante. En la parte de la neurona que recibe esta comunicación, la neurona postsináptica, no se observan estructuras tan características, pero sí se sabe que están presentes unas proteínas muy importantes, los receptores, encargados de recibir el mensaje que la neurona presináptica quiere comunicar. Esta descripción corresponde a las sinapsis llamadas químicas, porque, como se verá después, se comunican a través de un mensajero químico. Existen, aunque en menor número, otro tipo de sinapsis en las cuales la comunicación entre las dos neuronas es directa y no necesita de un puente químico. Éstas son las sinapsis eléctricas que llevan a cabo una comunicación rápida y sencilla entre las neuronas. Las sinapsis químicas, en cambio, aunque más lentas, tienen mayores posibilidades, como se explicará después.
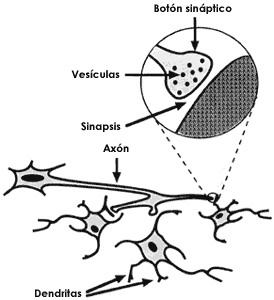
Figura I.6 Una neurona típica está formada por el soma y dos tipos de prolongaciones: las dendritas, cortas y ramificadas y el axón, más largo. En el extremo del axón se establece la comunicación con otras neuronas a través de las terminaciones o botones sinápticos que contienen las vesículas sinápticas donde se almacenan los neurotransmisores.
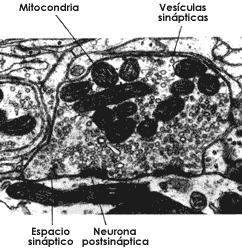
Figura I.7. Estructura de la sinapsis en la que se observan el espacio sináptico, las vesículas sinápticas de la neurona presináptica y los engrosamientos típicos de la neurona postsináptica.
Las células del cerebro reciben decenas de estos mensajes de otras neuronas, la mayor parte de los cuales se transmiten a través de sinapsis de esta naturaleza. ¿Cómo se sabe que la neurona recibió un mensaje de otra neurona? Las neuronas manejan un lenguaje eléctrico, es decir, a base de cambios en las cargas eléctricas que llevan algunos elementos químicos, muy importantes para la función del cerebro que son los iones. Los más destacados son el sodio y el potasio, que tienen carga eléctrica positiva, y el cloro con carga eléctrica negativa. Estos iones son fundamentales para el sistema de comunicación de las neuronas. En el interior de las células nerviosas predomina el potasio y algunas proteínas también con carga eléctrica, mientras que afuera existe una alta concentración de sodio y cloro. Estas diferencias en la concentración de las moléculas cargadas dan como resultado una diferencia en la distribución de las cargas eléctricas y éste es el lenguaje que entienden las neuronas. Cuando la neurona está "callada", su interior es más negativo eléctricamente que el exterior, pero esta situación cambia abruptamente cuando la neurona se comunica con otras neuronas. En realidad, una neurona se comunica con muchísimas otras neuronas al mismo tiempo (figura I.8). Le puede llegar una cantidad enorme de mensajes que la neurona integra conjuntamente y, de acuerdo con la resultante de esta integración, tendrá una carga más negativa o más positiva que en el estado de reposo. Estos mensajes en realidad consisten en un cambio en la distribución de las cargas eléctricas adentro de la neurona porque su membrana se hizo más o menos permeable a los iones y el cambio de la permeabilidad de la membrana se debe a la acción de sustancias químicas, los neurotransmisores, que son los comunicadores de la relación entre las neuronas.
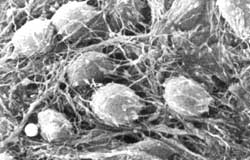
Figura I.8. Imagen de microscopía electrónica en la que se observan los cuerpos de las neuronas, los axones y los botones sinápticos.
¿Qué sucede entonces? Si la neurona tiene una carga más positiva se genera una onda de información eléctrica, el potencial de acción (figura I.9) la cual se propaga muy rápidamente en el interior de la célula, en todas direcciones y también a través del axón que, recordemos, tiene en su extremo la terminal por la que se comunicará con la siguiente neurona. Si el potencial de acción al final del axón llega a una sinapsis eléctrica, la corriente pasa directamente a la siguiente neurona, pero si se trata de una sinapsis química lo que sucede es que el cambio en la carga eléctrica abre unos poros por los cuales entra a la célula el ion calcio, muy importante para el funcionamiento del sistema nervioso. Cuando aumenta la concentración de calcio en la terminación presináptica, la neurona lanza al exterior el neurotransmisor, que constituirá un puente químico entre las dos neuronas. Lo que sucede después merece un párrafo aparte.
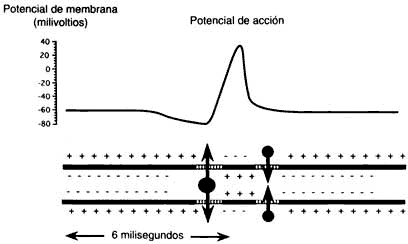
Figura I.9. El cambio en la permeabiliad de la membrana al sodio genera una onda eléctrica, el potencial de acción que se transmite por el axón hasta llegar a la terminación sináptica en la que induce la liberación del neurotransmisor.
LOS TRANSMISORES QUÍMICOS, INTERLOCUTORES DE LA COMUNICACIÓN NEURONAL
A principios de este siglo se iniciaron las investigaciones que poco a poco han ido esclareciendo el complicadísimo proceso de la comunicación entre las células nerviosas. El descubrimiento de que esta comunicación se lleva a cabo mediante sustancias químicas está asociado a una anécdota muy simpática, como hay muchas en la historia de los descubrimientos. El investigador alemán Otto Loewi, en los años treinta, estaba estudiando la forma como las células nerviosas transmiten su mensaje a las fibras musculares del corazón de la rana, y tenía la idea de que esta comunicación estaba mediada por una sustancia química que, liberada de los nervios, o sea, de la sinapsis del final del axón, transmitiría una señal a las fibras musculares del corazón, del mismo modo que una neurona se comunica con otra. El mismo cuenta que una noche, cuando estaba medio dormido, se le ocurrió una forma muy simple para probar su hipótesis. Vio con claridad meridiana las posibilidades de demostrar sus ideas gracias a un diseño experimental muy sencillo (que luego describiremos) y rápidamente tomando un papel y un lápiz esbozó los lineamientos del experimento. Feliz con su ocurrencia se durmió profundamente. Al día siguiente, en la adusta atmósfera del laboratorio, releyó las líneas garrapateadas la noche anterior, y analizando críticamente el experimento, desechó la idea de llevarlo a cabo, considerándolo demasiado elemental. Algunas semanas después, de nuevo en la semiinconsciencia del sueño, volvió a ver con claridad las potencialidades del experimento que había concebido. Esta vez no esperó a la mañana siguiente. En ese mismo momento se vistió, fue al laboratorio, tomó sus ranas y realizó el experimento que dio inicio a toda la moderna bioquímica del cerebro.
¿EN QUÉ CONSISTIÓ ESTE EXPERIMENTO?
Todo aquel que haya pasado por la escuela secundaria sabe que el corazón de los animales, y el de las ranas en particular; durante un tiempo sigue latiendo después de haberse extraído, si se coloca en una solución que contenga los elementos básicos del plasma sanguíneo. Una técnica un poco más complicada es la extracción del corazón junto con los nervios que modulan el latido cardiaco. Un nervio no es otra cosa que un haz de axones de un conjunto de neuronas. Como describimos en el párrafo anterior; a través de los axones se envía el mensaje a la siguiente neurona o a una fibra muscular. Si se estimula el nervio, por ejemplo, con un choque eléctrico, el latido del corazón disminuye su fuerza y su frecuencia y esto puede registrarse mediante un sencillo equipo de laboratorio. En el experimento a que nos referimos, que se ilustra en la figura I.10, se extrajo un corazón de la rana, con su nervio respectivo, y se lo colocó en un mismo recipiente con otro corazón de rana pero sin que existiera entre ellos ningún contacto. La hipótesis del doctor Loewi era que la comunicación entre la neurona a través de su axón y la fibra muscular se llevaba a cabo a consecuencia de la liberación de una sustancia química. Por tanto, si esto fuera así, la estimulación del nervio del corazón número uno produciría no sólo una disminución en la fuerza contráctil de este corazón, sino que al difundirse la sustancia química hipotética a través del líquido en el que estaban bañados los dos corazones produciría una atenuación en la contracción similar en el corazón número dos, al cual no se había estimulado. Podemos imaginar la emoción del doctor Loewi cuando la plumilla del aparato que registraba las señales del corazón número dos comenzó a disminuir su ritmo hasta casi cesar las contracciones del corazón, y demostrando así su hipótesis. El análisis y la identificación de la sustancia química que constituía el puente de comunicación entre el nervio y el músculo no fueron muy complicados, y así se descubrió el primer neurotransmisor químico, al que se llamó acetilcolina.
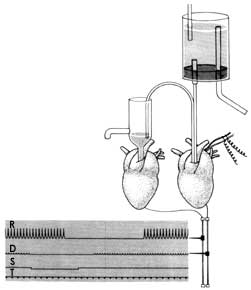
Figura I.10. Dispositivo experimental utilizado por Otto Loewi para demostrar la existencia de un neurotransmisor, en este caso la acetilcolina que reduce la contracción cardiaca.
¿CÓMO SON LOS TRANSMISORES QUÍMICOS?
Son generalmente sustancias sencillas, cuyas fórmulas químicas se representan en las figuras I.11A y I.11B. Considerando el número enorme de contactos que se establecen entre las neuronas, es sorprendente el número tan pequeño de moléculas que la naturaleza ha diseñado para transmitir los cientos de miles de mensajes entre las neuronas. Algunos de estos neurotransmisores participan sobre todo en las funciones motoras, como la acetilcolina, que es el transmisor de las órdenes que las neuronas dan a los músculos voluntarios, pero en muchos casos un solo neurotransmisor puede intervenir en la comunicación de neuronas que controlan funciones muy distintas. Por ejemplo, la propia acetilcolina participa también en los procesos de la memoria; la dopamina, como veremos más adelante, parece ser crucial en la génesis de trastornos mentales muy severos como la esquizofrenia, pero también está involucrada en el movimiento y, su deficiencia es la causa de las alteraciones motoras que se observan en los enfermos de Parkinson.
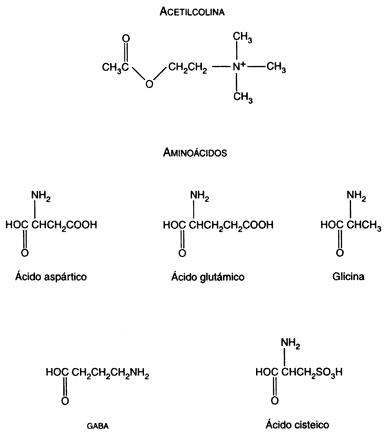
Figura I.11.A. Estructura química de los principales neurotransmisores.
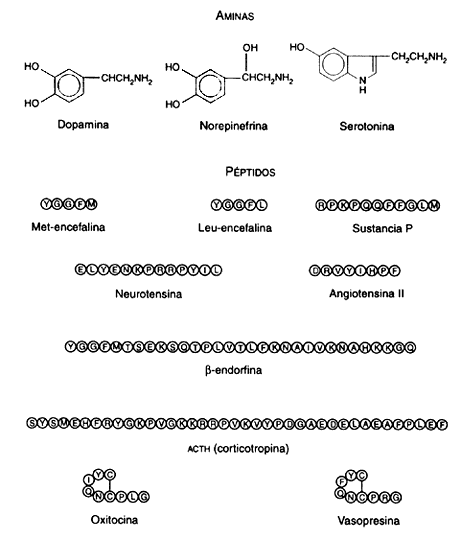
Figura I.11.B.
Los neurotransmisores pueden clasificarse, desde el punto de vista de su estructura, en tres grandes grupos: los aminoácidos, las aminas y los péptidos. Todos ellos parecen intervenir en el origen y control de las emociones, aunque de algunos de ellos sabemos más que de otros.
LA CONVERSACIÓN ENTRE LAS NEURONAS: TRANSMISIÓN SINÁPTICA
En párrafos anteriores describimos cómo una neurona, después de integrar todos los mensajes que recibe, puede transmitir su propio mensaje a la célula con la que se comunica. Este mensaje es llevado por el neurotransmisor químico, también llamado mensajero químico, el cual finalmente conducirá a un cambio en la permeabilidad de la membrana de la neurona a la cual fue enviado, con lo que el mensaje se habrá transmitido. Si se trata de un transmisor inhibidor; el cambio en la permeabilidad de la membrana hará el interior de la célula más negativo eléctricamente, dificultando de esta forma la excitación de la neurona. Si, por el contrario, se trata de un transmisor excitador, el cambio de la permeabilidad de la membrana será de naturaleza tal que haga menos negativo el interior de la célula, facilitando así la generación del impulso nervioso. Todas estas etapas son de profundo interés para el tema de este libro, ya que una alteración en cualquiera de estos pasos de comunicación puede dar como resultado profundos cambios en el comportamiento. Vale la pena, pues, dedicar cierto espacio a describirlos en forma sencilla.
¿CÓMO SE LIBERA EL NEUROTRANSMISOR?
Al llegar el impulso nervioso a la sinapsis, el cambio en la carga eléctrica abre canales por los cuales pasa el calcio. Este ion existe en concentraciones pequeñísimas en el interior de la célula cuando ésta se encuentra en reposo, pero al abrirse estos canales entra al extremo del axón, que es un elemento clave para la liberación del neurotransmisor. A pesar de que este hecho se conoce desde los trabajos clásicos que un compatriota nuestro, Ricardo Miledi, hiciera en colaboración con Bernard Katz, a fines de los años sesenta en Inglaterra, los detalles del proceso todavía no están aclarados por completo. Sea cual fuere el mecanismo, el resultado es que los neurotransmisores son expulsados de la neurona presináptica para llevar el mensaje a la postsináptica. Los neurotransmisores se almacenan en las estructuras características de la presinapsis, las vesículas sinápticas, y permanecen ahí secuestrados hasta que el calcio los hace salir en camino hacia la neurona a la que han de transmitir el mensaje (figura 1.12).
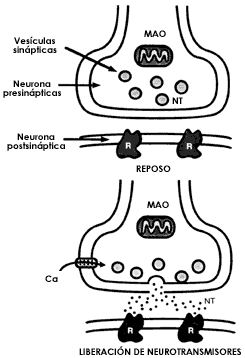
Figura I.12. Esquema de la estructura de las sinapsis mostrando la terminación de la neurona presináptica con las vesículas donde se almacenan los neurotransmisores y la neurona postsináptica en la que se encuentran los receptores. En el esquema superior se muestra la sinapsis en reposo. En el inferior se ilustra el proceso de liberación del transmisor de las vesículas, subsecuente a la apertura de los canales de calcio.
LAS MOLÉCULAS QUE RECIBEN EL MENSAJE: LOS RECEPTORES
Una vez que los neurotransmisores han salido de la presinapsis, cruzan el espacio sináptico, y ya en la membrana de la neurona postsináptica interactúan con una proteína, el receptor; que se halla inserto en la membrana y que los reconoce, casi como una cerradura reconoce una sola llave. Este contacto del receptor con el transmisor origina el mensaje que reconocen las neuronas, es decir; un cambio en la permeabilidad celular a un determinado ion y el cambio consecuente en la distribución de las cargas eléctricas. En algunos casos, el receptor es el propio canal a través del cual entran los iones, por ejemplo, el sodio. Normalmente el canal está cerrado, pero se abre al entrar en contacto con el transmisor (figura I.13). Aquí podría hablarse de una conversación directa. En otros casos, la interacción transmisor-receptor desencadena una serie compleja de reacciones químicas que culminan con la apertura de muchos canales iónicos, llevando al resultado final que es el cambio en la permeabilidad de las neuronas, es decir; el mensaje que la neurona quería transmitir (figura I.13).
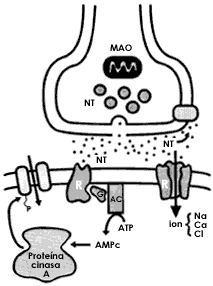
Figura I.13. Los receptores postsinápticos son de dos tipos. En uno de ellos el propio receptor es el canal por donde se mueven los iones que cambiarán el estado eléctrico de las neuronas. El receptor-canal se activa al interactuar con el neurotransmisor. En el otro tipo el receptor, al unirse al neurotransmisor, desencadena una serie de reacciones metabólicas mediadas por sistemas de segundos mensajeros como las proteínas G y el AMPc que conducen finalmente a la activación de un canal iónico.
Los receptores postsinápticos desempeñan un papel clave en la fisiología de la conducta, como veremos después. En un principio se pensaba que cada neurotransmisor se comunicaba con un solo tipo de molécula receptora y así se hablaba del receptor del GABA, de la dopamina, de la serotonina, etc. Poco a poco se ha ido descubriendo que los receptores de un mismo neurotransmisor no son siempre iguales, sino que existen familias de receptores que, si bien interactúan con el mismo neurotransmisor; tienen diferencias tales en su estructura que obligan a pensar que son moléculas distintas. Como se han identificado estas diferencias es por la manera en que estos receptores reaccionan con distintas sustancias creadas en los laboratorios de investigación. Así se ha encontrado, por ejemplo, que existen al menos cinco subtipos del receptor de la dopamina, cuatro subtipos del receptor de la serotonina, y muchos más que están por descubrirse. Esta variedad de subtipos de receptores es muy importante ya que tal vez sea el mecanismo que permita que, manejando un solo neurotransmisor; puedan ejercerse acciones diferentes en las distintas células. Además, esta circunstancia hace posible que en los laboratorios puedan sintetizarse distintos fármacos para los diferentes subtipos de receptores, permitiendo así una manipulación más eficiente y selectiva de las funciones a cargo de un determinado neurotransmisor.
Una vez que el mensaje ha sido transmitido, el neurotransmisor; ya terminada su función, debe dejar de interactuar con el receptor y desaparecer del espacio sináptico para que pueda iniciarse una nueva comunicación, si es necesario. Existen dos tipos de acciones que permiten que esto suceda: en algunos casos, el neurotransmisor es destruido, en los más, es transportado de nuevo a las neuronas. Estos mecanismos de inactivación de los mensajeros químicos pueden estar modificados en muchas de las alteraciones de la conducta, como se verá después, y son también el sitio de acción de muchas drogas.
AMPLIFICANDO LA CONVERSACIÓN: LOS SEGUNDOS Y TERCEROS MENSAJEROS
Como se describió en el párrafo anterior; a veces los receptores son, en sí mismos, canales a través de los cuales pasan los iones, lo que equivale a decir que cada receptor abre una sola puerta a los iones que representarán el mensaje de la neurona presináptica. Es ésta una comunicación rapidísima que dura sólo milésimas de segundo. Otras veces, lo que hacen los receptores una vez activados por su interacción con el transmisor (es decir; cuando la llave abrió la cerradura) es desencadenar una serie de reacciones químicas, ilustradas en la figura I.14, mediadas por lo que se llaman segundos mensajeros (los primeros son los neurotransmisores) que llevan al resultado final que, en vez de abrir una sola puerta para la entrada de los iones, abren muchísimas al mismo tiempo. Aunque más lenta, dura algunos segundos, esta comunicación a través de los segundos mensajeros es finalmente, mucho más eficiente.
Los segundos mensajeros son moléculas pequeñas como los nucleótidos cíclicos
(el AMP cíclico y el GMP cíclico), el calcio, algunos
fosfoionosítidos, así como el ácido araquidónico y sus derivados (figura I.14).
La comunicación entre los receptores y los segundos mensajeros la hacen moléculas
más grandes, las llamadas proteínas G, las cuales a su vez regulan la actividad
de las proteínas efectoras, como las adenilciclasas, las fosfolipasas A2
y C y las fosfodiesterasas (figura I.14) El óxido nítrico es una molécula cuya
función como segundo mensajero se ha descubierto recientemente. Este compuesto
muy pequeño se forma en algunas neuronas, a partir del aminoácido arginina,
se difunde libremente en el tejido nervioso y tiene la capacidad de incrementar
la formación del GMP cíclico, un importante segundo mensajero (figura
I.14).
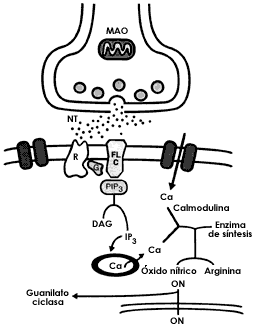
Figura I.14. Esquema de los sistemas de segundos mensajeros que tienen a su cargo la amplificación de la conversación entre las neuronas.
A través de los segundos mensajeros, como se mencionó antes, a partir de una sola interacción transmisor-receptor conducen a la apertura de muchos canales para la entrada de iones, es decir; de muchos mensajes eléctricos, para la neurona, con lo que la conversación entre las neuronas se ve enormemente amplificada Es necesario hacer notar que es un proceso muy complejo, en el que los distintos eslabones deben funcionar de manera acoplada y muy precisa para llegar sin tropiezos al resultado final.
Algunos de los segundos mensajeros, como el AMP cíclico, el calcio
y el diacilglicerol que, como se mencionó, se forman como consecuencia de la
interacción del neurotransmisor con el receptor; pueden llegar a actuar hasta
a un nivel genético, acelerando la transcripción de los genes de las neuronas.
La interacción entre los segundos mensajeros y los genes ocurre a través de
los llamados factores de transcripción que incluyen entre los más comunes
el CREB (por su nombre en inglés cyclic AMP response
element-binding protein), que funciona en asociación con el AMP
cíclico y los llamados fos y jun. Estos elementos y sus
acciones sobre los genes constituyen una forma de respuesta de las neuronas
a más largo plazo, que pueden modificar, por ejemplo, las enzimas que forman
los neurotransmisores para incrementar su concentración o bien, como en el caso
de fos y jun, que llevan a la expresión inmediata de genes después
de la estimulación nerviosa, podrían participar en fenómenos de más corta duración.
El estudio de este grupo de terceros mensajeros es relativamente reciente y
está en plena expansión.
INTERFERENCIAS EN LA COMUNICACIÓN NEURONAL
La comunicación entre las neuronas consta, como se ha visto, de numerosas etapas, y en cada una de ellas puede darse una interrupción por diversas causas. Muchos venenos animales como los de las serpientes, arañas y escorpiones, producen daño y a veces la muerte, porque precisamente interrumpen la comunicación entre las neuronas.
El proceso de salida del neurotransmisor se puede alterar, por ejemplo, con sustancias que cierran la entrada de los canales de calcio, interceptando así la señal para liberar el neurotransmisor. Algunas sustancias de este tipo son útiles en el tratamiento de los desarreglos de la presión arterial, modificando la salida del neurotransmisor que comunica los nervios con los músculos de los vasos sanguíneos. Otra forma de modificar el proceso de liberación del neurotransmisor es impedir su entrada a las vesículas sinápticas. Cuando esto ocurre, el transmisor que se va acumulando en la terminal presináptica inunda la sinapsis y altera, en consecuencia, el proceso de comunicación normal entre las neuronas. Algunas toxinas, como el veneno de la viuda negra, incrementan en forma extraordinaria e indiscriminada la salida de los neurotransmisores de las vesículas, con lo que alteran los mecanismos normales de comunicación, en particular la de las neuronas con los músculos, y las personas afectadas mueren generalmente de parálisis respiratoria. La muerte por botulismo ocasionada por la acción de una de las más potentes toxinas que producen las bacterias que se desarrollan en alimentos contaminados, se debe a que la toxina impide la liberación de los neurotransmisores. Como se verá en su oportunidad, muchas de las drogas que alteran la conducta humana ejercen sus efectos precisamente en esta parte del proceso de comunicación interneuronal.
Los inhibidores de los transportadores que, como vimos, tienen la importante función de terminar la acción de los neurotransmisores, se están usando actualmente para evitar el daño producido por los accidentes vasculares cerebrales y en algunos casos de epilepsia. Algunas de las drogas psicoactivas, como la cocaína y las anfetaminas, actúan en parte a través de una modificación en la eficiencia del transportador de algunos neurotransmisores, como se verá en el capítulo correspondiente.
Los receptores, es decir las proteínas con las que interactúan los neurotransmisores, también pueden ser afectados por sustancias, algunas naturales, otras sintetizadas en el laboratorio, que son capaces de unirse a ellos como si fueran los propios neurotransmisores, pero que no actúan como tales y, dependiendo de cuál sea su efecto sobre el receptor y cuál el neurotransmisor afectado, tendrán consecuencias importantes en la comunicación interneuronal.
La mordedura de una cobra causa un cuadro bien identificado. El individuo tiene abundante salivación, dificultad para respirar; y puede morir de un paro respiratorio. Ahora sabemos qué sucede a nivel molecular con el veneno de la cobra. Este contiene una serie de compuestos que por un lado destruyen las membranas de las células haciendo que el veneno penetre más rápido, pero el principio activo más potente, que finalmente es el causante de la muerte, es una sustancia que se conoce como bungarotoxina y cuya acción está perfectamente identificada. La bungarotoxina se combina con el receptor de la acetilcolina que, como mencionamos antes, es el neurotransmisor que comunica las neuronas con los músculos. La bungarotoxina se une a este receptor muy rápidamente y es capaz de desplazar de la unión con el receptor a la propia acetilcolina. Pero a diferencia de la unión con su transmisor, que es transitoria, la bungarotoxina se une en forma permanente, de modo que impide la comunicación de los nervios con el músculo, precisamente a nivel de la sinapsis. Éste es un ejemplo muy claro de lo que se llama un antagonista, es decir; un compuesto suficientemente parecido a un neurotransmisor como para ocupar su lugar en el receptor; pero que, por una parte, no lleva a cabo la función del transmisor que, como ya sabemos, es la de cambiar la permeabilidad de la membrana y, por otra, no permite que el verdadero neurotransmisor ocupe su lugar en el receptor. En estas condiciones, la comunicación entre las neuronas queda interrumpida y las consecuencias pueden ser fatales. La bungarotoxina es, por lo tanto, un antagonista del receptor a la acetilcolina.
Otro ejemplo de esta acción de los antagonistas es el curare. Conocido desde tiempo inmemorial por los indígenas del Amazonas, este veneno se extrae de las raíces de una planta. Al ser introducido al organismo mediante la punta de una flecha, se transporta a través de la circulación hasta las sinapsis neuromusculares y ahí, provoca el mismo efecto que el veneno de la cobra, es decir; antagoniza a los receptores de la acetilcolina, con lo cual se impide la interacción normal de este transmisor con su receptor y se interrumpe la comunicación nervio-músculo. El resultado es la muerte por paro respiratorio. Es por ello que las flechas envenenadas con curare son mortales a pesar de que no lesionen ninguna víscera vital.
Los antagonistas de los receptores actúan de esa forma en muchas ocasiones porque en su estructura química tienen una parte de su molécula suficientemente parecida a la del neurotransmisor natural, lo que les permite acomodarse en el sitio activo del receptor; o sea, la parte de la proteína con la que se asocia el transmisor. Pero la similitud llega hasta allí. El receptor no responde a la interacción con esta molécula "impostora" y, por supuesto, no tienen lugar las reacciones que normalmente ocurren con el transmisor natural. Es como si en una cerradura se hubiera introducido, tal vez con un poco de esfuerzo, una llave falsa para abrir la puerta que, en el caso de la sinapsis que emplea la acetilcolina como neurotransmisor; es un canal de sodio. Peor aún, en muchos casos, como en el del veneno de la cobra, la falsa llave se queda atorada en la cerradura y la deja inutilizada, a veces durante tanto tiempo que la muerte sobreviene antes de que la unión se rompa.
Los agonistas son moléculas que, en muchos casos por la similitud que tienen con la estructura del neurotransmisor; también pueden ocupar el sitio activo del receptor —la cerradura— pero, a diferencia de los antagonistas, los agonistas funcionan aún mejor que los verdaderos neurotransmisores, lo cual puede ser bueno, pero también muy peligroso. Esto es cierto particularmente en el caso de que los neurotransmisores sean las catecolaminas, mismas que, como veremos, han tenido un papel clave en la asociación de la neuroquímica con las emociones.
![[Inicio]](../img/begin.gif)
![[Anterior]](../img/prevsec.gif)

![[Siguiente]](../img/nextsec.gif)